
Analysis of Combining Ability to Obtain Tropical Carrot Hybrids for Production Traits

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Material, Experimental Site, and Flow
2.2. Assessment of Agricultural Traits in the Field
2.3. Diallel Analysis
2.4. GGE Biplot
2.5. Biochemical Analysis of Pigment Extraction from Leaves and Root
3. Results and Discussion
3.1. Diallel Analysis
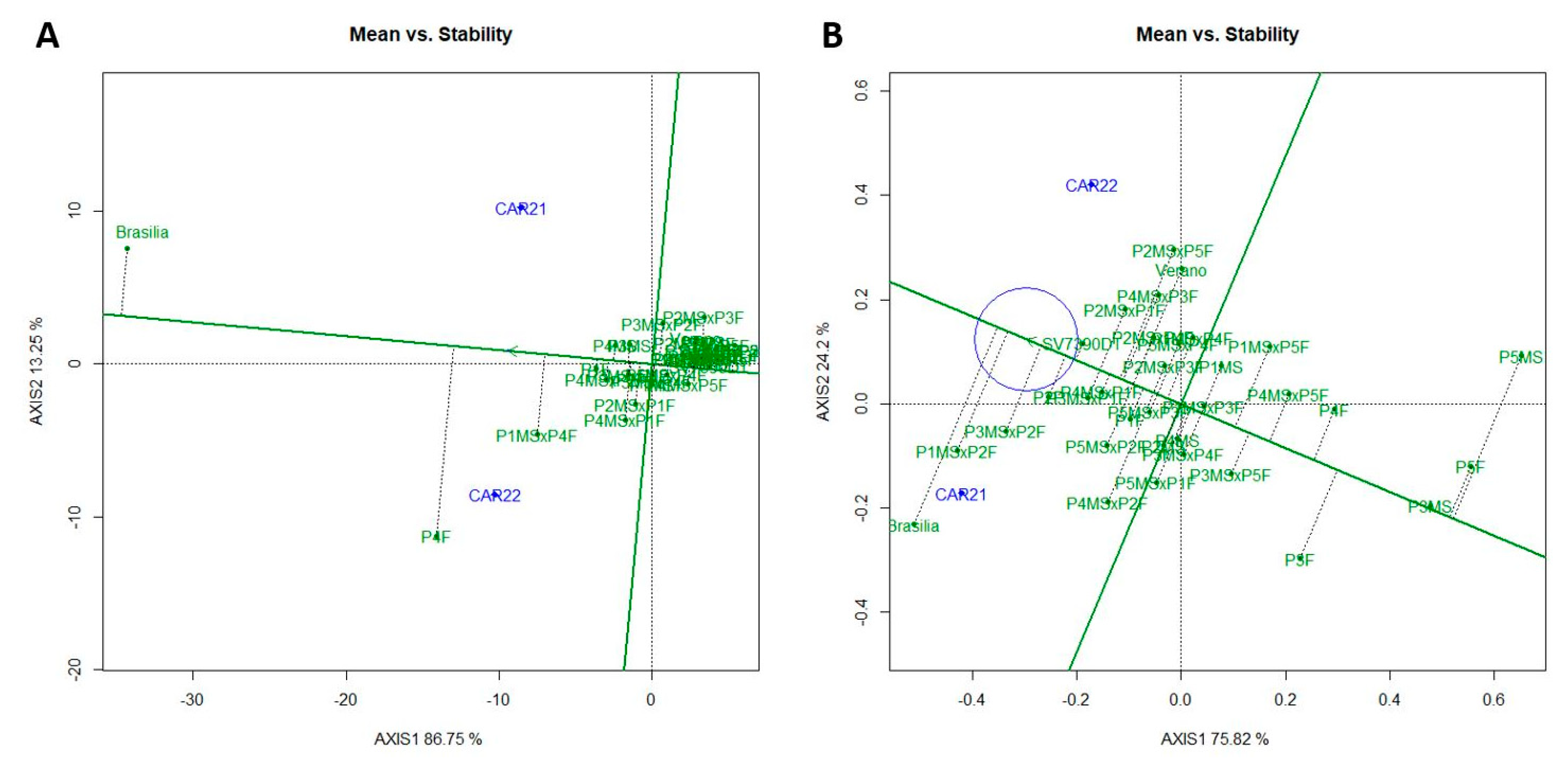
3.2. GGE Biplot
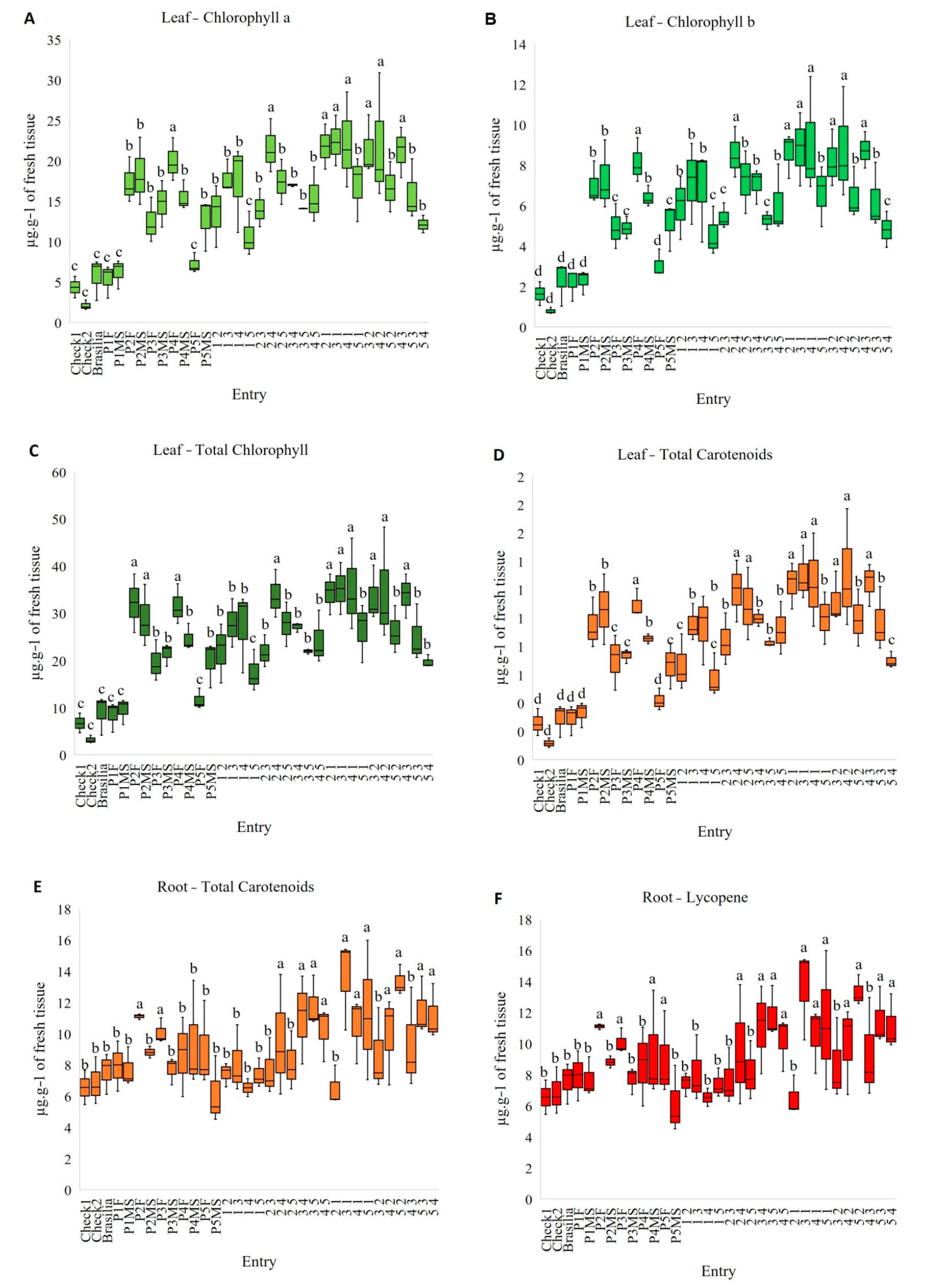
3.3. Biochemical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Selvakumar, R.; Kalia, P.; Raje, R.S. Genetic analysis of nutritional traits in tropical carrot (Daucus carota L.). Genetika 2019, 51, 641–660. [Google Scholar] [CrossRef]
- Yoo, K.S.; Bang, H.; Pike, L.; Patil, B.S.; Lee, E.J. Comparing carotene, anthocyanins, and terpenoid concentrations in selected carrot lines of different colors. Hortic. Environ. Biotechnol. 2020, 61, 385–393. [Google Scholar] [CrossRef]
- Chevalier, W.; Moussa, S.A.; Ottoni, M.M.N.; Dubois-Laurent, C.; Huet, S.; Aubert, C.; Desnoues, E.; Navez, B.; Cottet, V.; Chalot, G.; et al. Multisite evaluation of phenotypic plasticity for specialized metabolites, some involved in carrot quality and disease resistance. PLoS ONE 2021, 16, e0249613. [Google Scholar] [CrossRef] [PubMed]
- FAO. Food and Agriculture Organization of the United Nations Statistics Database. Available online: http://www.fao.org/faostat/en/#data (accessed on 2 May 2023).
- IBGE. Instituto Brasileiro de Geografia e Estatística. In Produção de Cenoura; 2017. Available online: https://www.ibge.gov.br/explica/producao-agropecuaria/cenoura/br (accessed on 14 February 2024).
- Turner, S.D.; Maurizio, P.L.; Valdar, W.; Yandell, B.S.; Simon, P.W. Dissecting the genetic architecture of shoot growth in carrot (Daucus carota L.) using a diallel mating design. G3 Genes Genomes Genet. 2018, 8, 411–426. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, A.D.F.D.; Silva, G.O.D.; Pereira, R.B. Capacidade de combinação de genitores de cenoura para caracteres de produtividade de raízes e tolerância à queima-das-folhas. Rev. Ceres 2016, 63, 183–190. [Google Scholar] [CrossRef]
- Cruz, C.D.; Regazzi, A.J.; Carneiro, P.C. Métodos Biométricos Aplicados ao Melhoramento Genético, 4th ed.; UFV: Viçosa, Brazil, 2012; p. 414. [Google Scholar]
- El-Azeem, A.; Aly, R.S.H.; El Sayed, W.M.; Hassan, N.A. Combining ability and gene action using 10 x 10 diallel crosses of ten maize inbred lines (Zea mays L.). J. Plant Prod. 2021, 12, 1205–1211. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, H.R.; Kumar, M. Heterosis for post harvest and nutritional quality traits in tomato (Solanum lycopersicum L.). J. Appl. Nat. Sci. 2016, 8, 1987–1991. [Google Scholar] [CrossRef]
- Datta, D.R.; Rafii, M.Y.; Misran, A.; Jusoh, M.; Yusuff, O.; Haque, M.A.; Jatto, M.I. Half diallel analysis for biochemical and morphological traits in cultivated eggplants (Solanum melongena L.). Agronomy 2021, 11, 1769. [Google Scholar] [CrossRef]
- Patil, D.G.; Subramaniam, V.R. Combining Ability Analysis for Yield and Processing Qualities in White Onion (Allium cepa L.). Curr. Agric. Res. 2020, 8, 18–24. [Google Scholar] [CrossRef]
- Pereira, R.B.; De Carvalho, A.D.F.; Pinheiro, J.B.; Da Silva, G.O.; Vieira, J.V. Resistência de populações de cenoura à queima-das-folhas com diferentes níveis de germoplasma tropical. Hortic. Bras. 2012, 30, 489–493. [Google Scholar] [CrossRef]
- Griffing, B.A. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Cruz, C.D. Genes: A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Yan, W. GGE biplot–A Windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 23 May 2023).
- Francis, F.J. Analysis of anthocyanins. In Anthocyanins as Food Colors, 1st ed.; Markakis, P., Ed.; Academic Press: New York, NY, USA, 1982; Volume 1, pp. 182–208. [Google Scholar]
- Cassetari, L.S.; Gomes, M.S.; Santos, D.C.; Santiago, W.D.; Andrade, J.; Guimarães, A.C.; Souza, J.A.; Cardoso, M.G.; Maluf, W.R.; Gomes, L.A. β-Carotene and chlorophyll levels in cultivars and breeding lines of lettuce. Acta Hortic. 2015, 1083, 469–473. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. Harvestplus Handbook for Carotenoid Analysis; International Food Policy Research Institute (IFPRI): Washington, DC, USA; International Center for Tropical Agriculture (CIAT): Palmira, CO, USA, 2004; 58p. [Google Scholar]
- De Carvalho, A.D.F.; Da Silva, G.O.; Pereira, G.E. Direct selection for phenotypic traits in carrot genotypes. Hortic. Bras. 2019, 37, 354–358. [Google Scholar] [CrossRef]
- Cruz, C.D.; Vencovsky, R. Comparação de alguns métodos de análise dialélica. Rev. Bras. Genet. 1989, 12, 425–436. [Google Scholar]
- Carvalho, A.D.F.; Silva, G.O.; Pereira, R.B.; Pinheiro, J.B.; Vieira, J.V. Capacidade combinatória em cenoura para componentes de produção e tolerância à queima-das-folhas. Hortic. Bras. 2014, 32, 190–193. [Google Scholar] [CrossRef]
- Jagosz, B. Combining ability of carrot (Daucus carota L.) lines and heritability of yield and its quality components. Folia Hortic. 2013, 24, 115–122. [Google Scholar] [CrossRef]
- Kaur, K.; Dhillon, T.S.; Singh, R. GGE biplot analysis for seed production potential of European carrot (Daucus carota L.) genotypes in plains and submountenous zones of Punjab. Genetika 2022, 54, 233–254. [Google Scholar] [CrossRef]
- Frutos, E.; Galindo, M.P.; Leiva, V. An interactive biplot implementation in R for modeling genotype-by-environment interaction. Stochastic Environmental Research and Risk. Assessment 2014, 28, 1629–1641. [Google Scholar]
- Singamsetti, A.; Shahi, J.P.; Zaidi, P.H.; Seetharam, K.; Vinayan, M.T.; Kumar, M.; Singla, S.; Shikha, K.; Madankar, K. Genotype × environment interaction and selection of maize (Zea mays L.) hybrids across moisture regimes. Field Crops Res. 2021, 270, 108224. [Google Scholar] [CrossRef]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Silva, G.O.D.; Carvalho, A.D.F.D.; Vieira, J.V.; Benin, G. Verificação da adaptabilidade e estabilidade de populações de cenoura pelos métodos AMMI, GGE biplot e REML/BLUP. Bragantia 2011, 70, 494–501. [Google Scholar] [CrossRef]
- Yadav, A.; Jakhar, R.K.; Kumari, N.; Yadav, G.N.; Kant, R.; Sharma, S.L.; Kumar, A. Response of organic manures and fertilizers on yield and quality of carrot under sandy soil condition. Pharma Innov. J. 2022, 11, 1822–1827. [Google Scholar]
- Terletskaya, N.V.; Korbozova, N.K.; Kudrina, N.O.; Kobylina, T.N.; Kurmanbayeva, M.S.; Meduntseva, N.D.; Tolstikova, T.G. The influence of abiotic stress factors on the morphophysiological and phytochemical aspects of the acclimation of the plant Rhodiola semenowii Boriss. Plants 2021, 10, 1196. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SV | DF | Mean Square | ||||||
|---|---|---|---|---|---|---|---|---|
| BP | DIS | RL | RD | LW | MYH | TYH | ||
| Block/Environment | 4 | 45.37 | 1.11 | 8.84 | 0.15 | 5.12 | 921.77 | 824.70 |
| Block | 2 | 60.16 | 1.51 | 17.58 | 0.26 | 0.37 | 1423.26 | 1293.43 |
| Block × Environment | 2 | 30.59 | 0.71 | 0.09 | 0.03 | 9.86 | 420.28 | 355.97 |
| Genotypes | 29 | 47.67 ** | 1.45 ** | 3.55 ** | 0.16 ** | 4.33 ** | 242.98 ** | 364.96 ** |
| Environment | 1 | 344.20 ns | 0.09 ns | 516.81 ** | 0.23 ns | 6.08 ns | 8572.84 * | 3603.09 ns |
| Genotypes × Environment | 29 | 26.11 ** | 0.72 * | 2.49 ns | 0.06 * | 0.37 ns | 80.44 ns | 88.58 ns |
| Residual | 116 | 3.19 | 0.42 | 1.59 | 0.04 | 0.37 | 85.80 | 69.92 |
| Overall mean | 3.06 | 3.38 | 18.78 | 3.31 | 3.44 | 30.95 | 47.24 | |
| CV (%) | 58.30 | 19.22 | 6.72 | 5.85 | 17.58 | 29.93 | 17.70 | |
| H2 (%) | 93.31 | 70.87 | 55.17 | 76.52 | 91.57 | 64.69 | 80.84 | |
| CVg (%) | 88.90 | 12.24 | 3.04 | 4.31 | 23.65 | 16.54 | 14.84 | |
| CVg/CVe | 1.52 | 0.64 | 0.45 | 0.74 | 1.35 | 0.55 | 0.84 | |
| SV | DF | Mean Square | ||||||
|---|---|---|---|---|---|---|---|---|
| BP | DIS | RL | RD | LW | MYH | TYH | ||
| Environment 1 | ||||||||
| Genotype (G) | 19 | 5.40 ** | 0.65 * | 2.78 ns | 0.08 * | 1.81 ** | 158.17 ns | 89.44 ns |
| GCA | 4 | 11.15 ** | 1.00 * | 4.10 ns | 0.13 * | 6.14 ** | 43.92 ns | 55.02 ns |
| SCA | 5 | 5.49 * | 0.81 ns | 0.91 ns | 0.01 ns | 1.11 * | 265.84 ns | 184.16 ns |
| Reciprocal | 10 | 3.07 ns | 0.43 ns | 3.18 ns | 0.09 * | 0.42 ns | 150.03 ns | 55.86 ns |
| Residual | 38 | 1.76 | 0.33 | 2.64 | 0.03 | 0.43 | 175.67 ns | 128.42 ns |
| Environment 2 | ||||||||
| Genotype (G) | 19 | 36.15 ** | 0.96 ns | 1.86 * | 0.04 ns | 1.76 ** | 48.08 ns | 58.09 ** |
| GCA | 4 | 107.52 ** | 1.52 ns | 3.82 ** | 0.03 ns | 6.62 ** | 60.57 ns | 100.23 ** |
| SCA | 5 | 13.34 * | 0.39 ns | 1.75 ns | 0.03 ns | 0.57 ns | 16.09 ns | 13.79 ns |
| Reciprocal | 10 | 19.02 ** | 1.01 ns | 1.13 ns | 0.05 ns | 0.42 ns | 59.07 * | 63.38 |
| Residual | 38 | 4.05 | 0.62 | 0.93 | 0.03 | 0.39 | 27.34 | 23.40 |
| Entry | BP | DIS | RL | RD | LW | MYH | TYH |
|---|---|---|---|---|---|---|---|
| Environment 1 | |||||||
| 1 | 0.11 | 0.17 | 0.29 | 0.03 | 0.17 | 2.05 | 2.02 |
| 2 | 0.45 | −0.33 | −0.76 | 0.12 | −0.84 | −0.78 | −0.22 |
| 3 | 0.38 | 0.28 | 0.34 | 0.02 | 0.14 | 0.34 | 0.99 |
| 4 | 0.46 | −0.06 | 0.31 | −0.06 | −0.22 | −2.13 | −0.13 |
| 5 | −1.38 | −0.06 | −0.18 | −0.10 | 0.75 | 0.53 | −2.66 |
| Environment 2 | |||||||
| 1 | 2.23 | 0.23 | −0.46 | 0.02 | 0.13 | −1.03 | −1.17 |
| 2 | −1.18 | −0.43 | −0.23 | 0.06 | −0.89 | 2.75 | 3.36 |
| 3 | −1.49 | 0.29 | −0.23 | −0.03 | −0.01 | 0.08 | 0.74 |
| 4 | 2.98 | −0.10 | 0.69 | 0.00 | −0.04 | −2.17 | −3.02 |
| 5 | −2.54 | 0.01 | 0.24 | −0.05 | 0.81 | 0.38 | 0.09 |
| Entry | BP | DIS | RL | RD | LW | MYH | TYH |
|---|---|---|---|---|---|---|---|
| Environment 1 | |||||||
| F1s | |||||||
| 1 × 2 | −1.11 | 0.34 | 0.14 | 0.04 | 0.31 | 1.48 | 0.83 |
| 1 × 3 | 0.15 | −0.11 | 0.21 | −0.03 | −0.04 | 0.95 | 3.62 |
| 1 × 4 | 0.32 | −0.28 | 0.13 | 0.01 | −0.31 | 1.89 | −1.72 |
| 1 × 5 | 0.64 | 0.06 | −0.47 | −0.02 | 0.05 | −4.32 | −2.74 |
| 2 × 3 | 1.17 | −0.45 | 0.03 | −0.01 | −0.25 | −3.83 | −6.02 |
| 2 × 4 | 0.40 | 0.22 | −0.02 | 0.00 | 0.44 | 6.01 | 6.64 |
| 2 × 5 | −0.46 | −0.11 | −0.15 | −0.03 | −0.50 | −3.65 | −1.45 |
| 3 × 4 | −0.93 | 0.28 | −0.48 | −0.01 | −0.15 | −6.50 | −3.36 |
| 3 × 5 | −0.39 | 0.28 | 0.25 | 0.05 | 0.44 | 9.38 | 5.75 |
| 4 × 5 | 0.21 | −0.22 | 0.37 | 0.00 | 0.02 | −1.40 | −1.57 |
| Reciprocal | |||||||
| 2 × 1 | −0.33 | 0.0 | 1.27 | 0.20 | 0.40 | 10.09 | 4.99 |
| 3 × 1 | −0.92 | 0.17 | −0.57 | −0.10 | −0.21 | −8.88 | −2.76 |
| 4 × 1 | 1.47 | 0.0 | −0.33 | −0.10 | 0.28 | −1.89 | −3.41 |
| 5 × 1 | 0.20 | 0.34 | 1.30 | −0.15 | 0.19 | 7.26 | 0.60 |
| 3 × 2 | −0.71 | 0.33 | 0.67 | −0.17 | −0.01 | −0.49 | −4.24 |
| 4 × 2 | −0.95 | 0.67 | 0.07 | −0.10 | −0.04 | 0.54 | −0.25 |
| 5 × 2 | 0.0 | 0.0 | −0.10 | −0.13 | 0.07 | 1.13 | −0.47 |
| 4 × 3 | 0.33 | 0.0 | 0.30 | 0.04 | 0.22 | −2.20 | 0.39 |
| 5 × 3 | 0.0 | 0.0 | 0.67 | −0.05 | −0.52 | −1.64 | −4.17 |
| 5 × 4 | −0.68 | 0.17 | −0.77 | −0.08 | −0.25 | −2.02 | −3.59 |
| Environment 2 | |||||||
| F1s | |||||||
| 1 × 2 | −0.32 | 0.08 | −0.22 | 0.02 | 0.04 | 0.79 | −0.48 |
| 1 × 3 | 0.01 | −0.31 | 0.55 | 0.00 | −0.29 | −0.14 | −0.54 |
| 1 × 4 | 2.05 | −0.09 | 0.13 | 0.03 | −0.04 | −1.90 | −1.21 |
| 1 × 5 | −1.74 | 0.31 | −0.46 | −0.05 | 0.29 | 1.25 | 2.23 |
| 2 × 3 | 0.31 | 0.19 | −0.01 | 0.00 | 0.38 | −1.48 | 0.08 |
| 2 × 4 | −0.80 | −0.08 | −0.39 | −0.10 | −0.04 | 0.73 | 0.73 |
| 2 × 5 | 0.82 | −0.20 | 0.62 | 0.07 | −0.37 | −0.03 | −0.32 |
| 3 × 4 | −1.24 | 0.20 | −0.06 | 0.04 | −0.05 | 2.01 | 1.43 |
| 3 × 5 | 0.93 | −0.08 | −0.48 | −0.04 | −0.04 | −0.38 | −0.96 |
| 4 × 5 | −0.01 | −0.03 | 0.33 | 0.03 | 0.13 | −0.84 | −0.95 |
| Reciprocal | |||||||
| 2 × 1 | −2.86 | −0.17 | −0.17 | −0.07 | −0.11 | 1.13 | −0.28 |
| 3 × 1 | −1.95 | −0.17 | −0.34 | −0.05 | −0.04 | −0.77 | −0.45 |
| 4 × 1 | 2.51 | 0.67 | 0.50 | 0.02 | 0.24 | −2.10 | −2.30 |
| 5 × 1 | −0.24 | 0.50 | 0.20 | 0.08 | 0.17 | −1.75 | −1.30 |
| 3 × 2 | −1.20 | −0.67 | 0.53 | 0.00 | 0.65 | 6.50 | 6.74 |
| 4 × 2 | −3.03 | 0.00 | 0.94 | 0.13 | 0.02 | 1.69 | 1.63 |
| 5 × 2 | 0.46 | 0.33 | −0.17 | 0.15 | 0.15 | 2.26 | 1.87 |
| 4 × 3 | 1.57 | −0.33 | 0.47 | −0.15 | −0.37 | −3.49 | −4.47 |
| 5 × 3 | −0.24 | 0.17 | −0.13 | −0.09 | −0.02 | −4.64 | −3.17 |
| 5 × 4 | −0.33 | −0.50 | −0.20 | −0.09 | −0.16 | −2.30 | −4.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, F.G.T.; Maciel, G.M.; Siquieroli, A.C.S.; de Oliveira, C.S.; Parrella, N.N.L.D.; da Silva, A.F.; Luz, J.M.Q.; Nogueira, A.P.O. Analysis of Combining Ability to Obtain Tropical Carrot Hybrids for Production Traits. Horticulturae 2024, 10, 442. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050442
Coelho FGT, Maciel GM, Siquieroli ACS, de Oliveira CS, Parrella NNLD, da Silva AF, Luz JMQ, Nogueira APO. Analysis of Combining Ability to Obtain Tropical Carrot Hybrids for Production Traits. Horticulturae. 2024; 10(5):442. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050442
Chicago/Turabian StyleCoelho, Fernanda Gabriela Teixeira, Gabriel Mascarenhas Maciel, Ana Carolina Silva Siquieroli, Camila Soares de Oliveira, Nádia Nardely Lacerda Durães Parrella, Amilton Ferreira da Silva, José Magno Queiroz Luz, and Ana Paula Oliveira Nogueira. 2024. "Analysis of Combining Ability to Obtain Tropical Carrot Hybrids for Production Traits" Horticulturae 10, no. 5: 442. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050442