
Elevated Altitude and Limestone Soil Promoted Fungal Diversity in Rhizosphere Soil of Sophora japonica
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Experimental Equipment
2.3. Determination of Physicochemical Properties of S. japonica Rhizosphere Soil
2.4. Determination of Microbial Diversity of S. japonica Rhizosphere Soil
3. Data Analysis
3.1. Bioinformatics Analysis
3.2. Statistical Analyses
4. Results
4.1. Physicochemical Properties of S. japonica Rhizosphere Soil
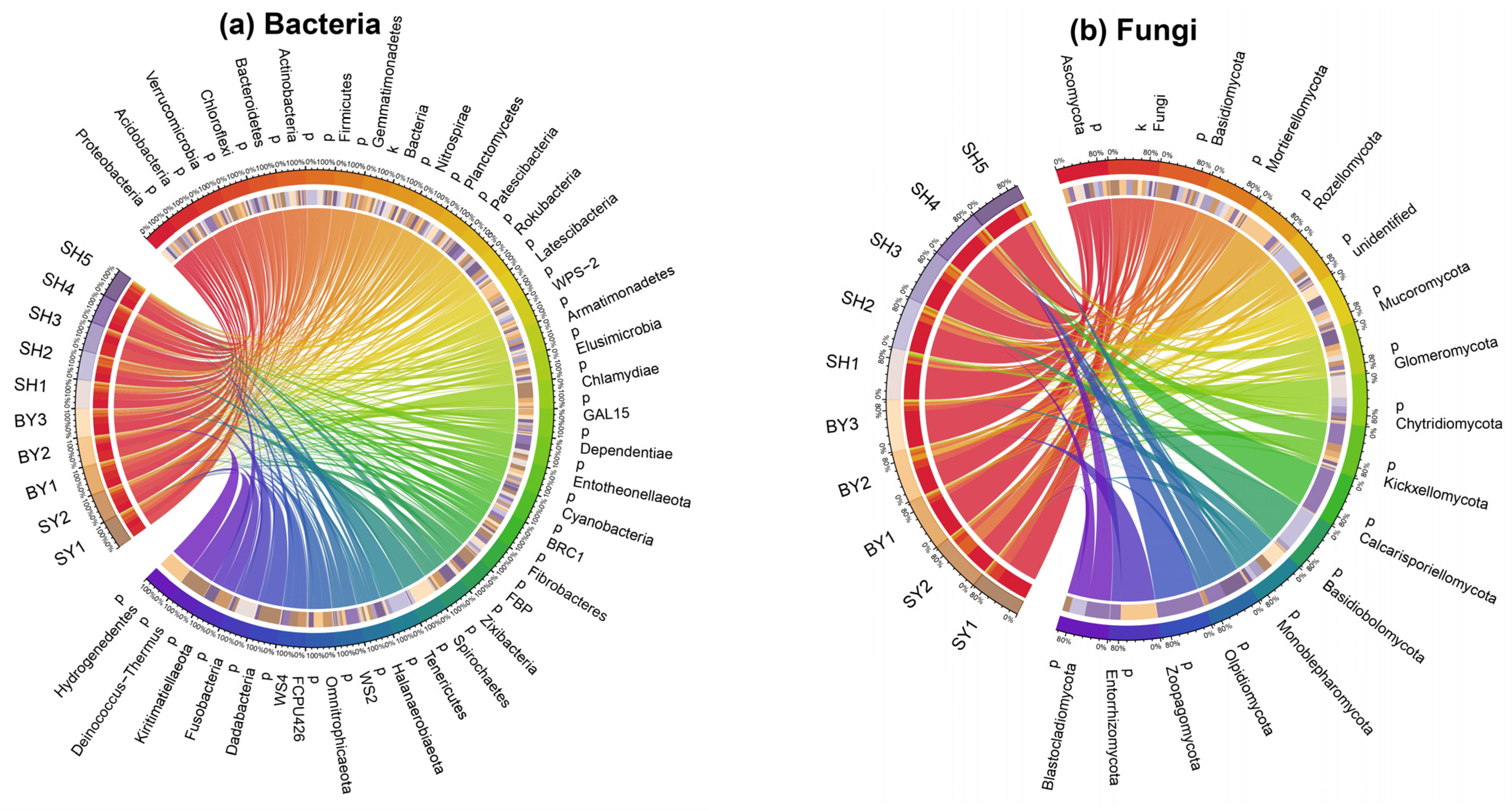
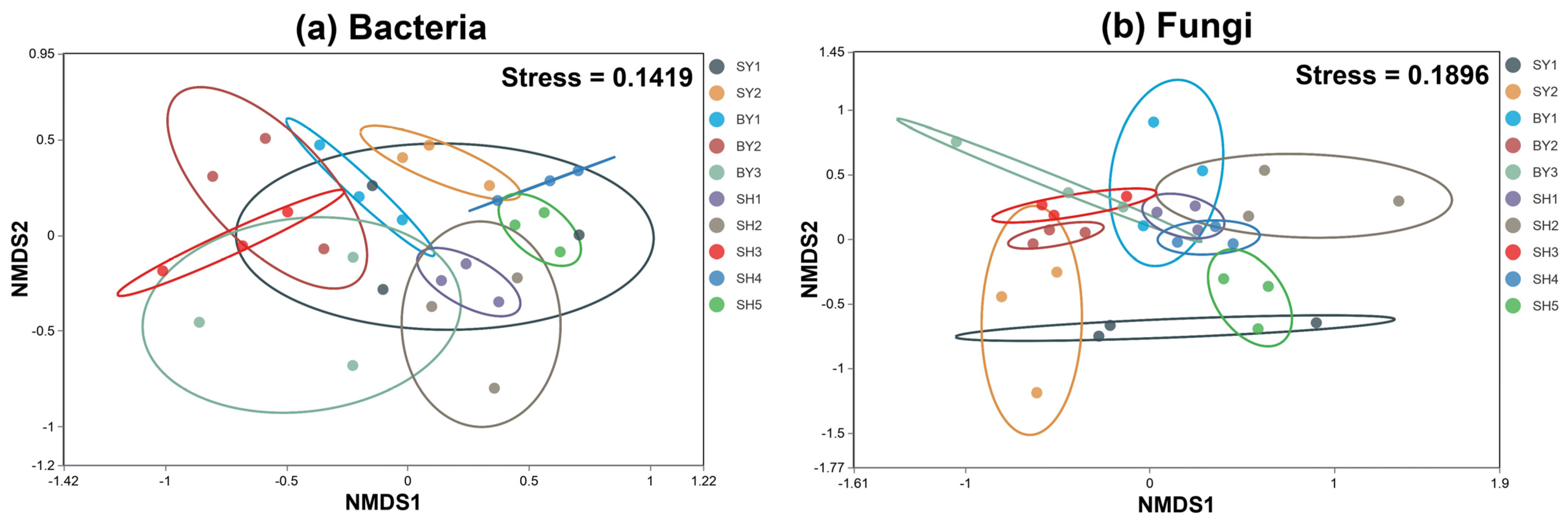
4.2. Rhizosphere Microbial Community Composition of S. japonica
4.3. Microorganism α Diversity of Rocky Desertification Degree and Altitude Gradient Pattern
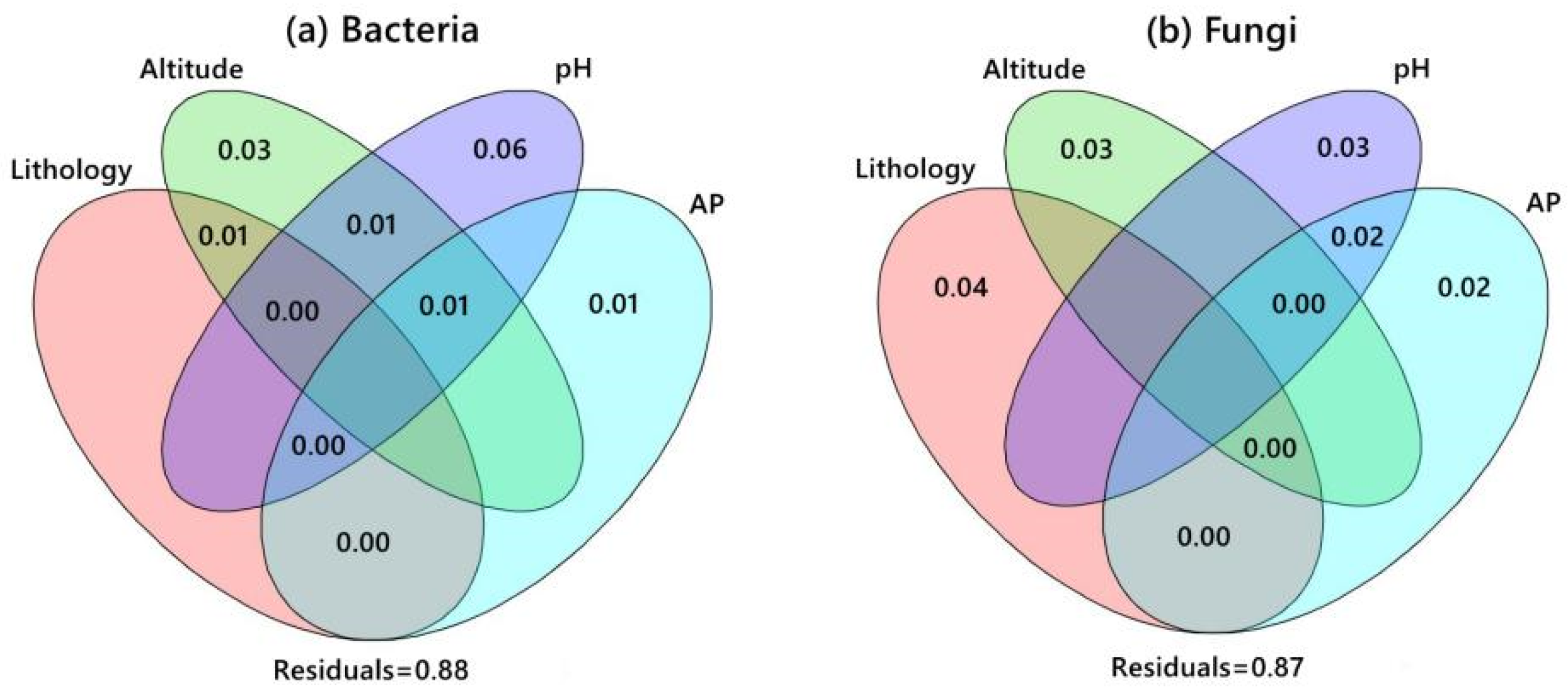
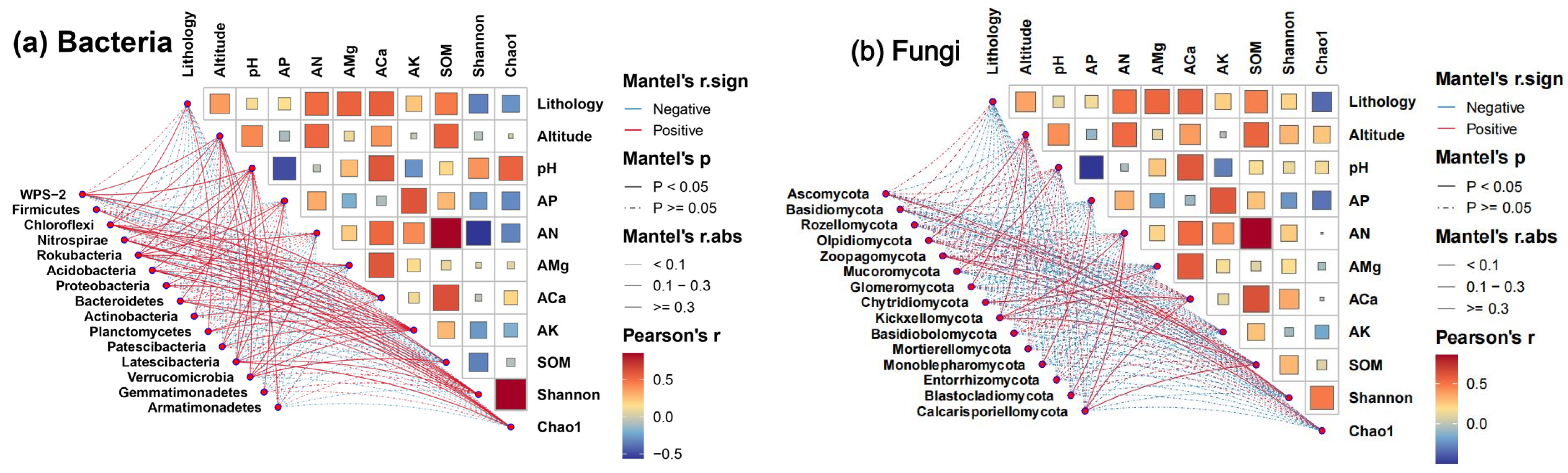
4.4. Relationship between External Factors and Microorganisms
4.5. Correlation between Dominant Microorganisms and Ecological Factors
5. Discussion
5.1. Rhizosphere Microorganisms Composition of S. japonica
5.2. Pattern of Microbial Diversity with Altitude
5.3. Relationship between Microbial Community and Lithology
5.4. Relationship between Microorganisms and Soil Factors
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gil-Martínez, M.; López-García, Á.; Domínguez, M.T.; Kjøller, R.; Navarro-Fernández, C.M.; Rosendahl, S.; Marañón, T. Soil fungal diversity and functionality are driven by plant species used in phytoremediation. Soil Biol. Biochem. 2021, 153, 108102. [Google Scholar] [CrossRef]
- Peay, K.G.; von Sperber, C.; Cardarelli, E.; Toju, H.; Francis, C.A.; Chadwick, O.A.; Vitousek, P.M. Convergence and contrast in the community structure of Bacteria, Fungi and Archaea along a tropical elevation-climate gradient. FEMS Microbiol. Ecol. 2017, 93, fix045. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Gunina, A.; Luo, Y.; Wang, J.; He, J.; Kuzyakov, Y.; Hemp, A.; Classen, A.T.; Ge, Y. Contrasting patterns and drivers of soil bacterial and fungal diversity across a mountain gradient. Environ. Microbiol. 2020, 22, 3287–3301. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Green, J.L. Microbes on mountainsides contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Fierer, N.; Turner, B.L.; Whitaker, J.; Ostle, N.J.; McNamara, N.P.; Bardgett, R.D.; Leff, J.W.; Salinas, N.; Silman, M.; et al. Microbes Follow Humboldt: Temperature drives plant and soil microbial diversity patterns from the Amazon to the Andes. Ecology 2018, 99, 2455–2466. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Shi, Y.; Fan, K.; He, J.S.; Adams, J.M.; Ge, Y.; Chu, H. Soil pH dominates elevational diversity pattern for bacteria in high elevation alkaline soils on the Tibetan Plateau. FEMS Microbiol. Ecol. 2019, 95, fiz003. [Google Scholar] [CrossRef]
- Tang, J.; Tang, X.; Qin, Y.; He, Q.; Yi, Y.; Ji, Z. Karst rocky desertification progress: Soil calcium as a possible driving force. Sci. Total Environ. 2019, 649, 1250–1259. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Fang, J.; Ji, C.; Shen, H.; Zheng, C.; Zhu, B. Soil microbial carbon and nutrient constraints are driven more by climate and soil physicochemical properties than by nutrient addition in forest ecosystems. Soil Biol. Biochem. 2019, 141, 107657. [Google Scholar] [CrossRef]
- Adamczyk, M.; Hagedorn, F.; Wipf, S.; Donhauser, J.; Vittoz, P.; Rixen, C.; Frossard, A.; Theurillat, J.; Frey, B. The soil microbiome of Gloria mountain summits in the Swiss Alps. Front. Microbiol. 2019, 10, 447428. [Google Scholar] [CrossRef]
- Zhang, J.; Li, T.; Jia, J.; Zhang, J.; Zhang, F. Bacterial taxa and fungal diversity are the key factors determining soil multifunctionality in different cropping systems. Land Degrad. Dev. 2021, 32, 5012–5022. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Hu, Y.; Fair, H.; Liu, Q.; Wang, Z.; Duan, B.; Lu, X. Diversity and co-occurrence networks of bacterial and fungal communities on two typical debris-covered glaciers, southeastern Tibetan Plateau. Microbiol. Res. 2023, 273, 127409. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Reitzel, K.; Nielsen, U.G.; Dupuy-Galet, B.X.; Leivers, S.; Horn, S.J.; Shapaval, V. Calcium affects polyphosphate and lipid accumulation in Mucoromycota fungi. J. Fungi. 2021, 7, 300. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Liu, J.; Zou, R.; Tang, J.; Xiong, Z.; Jiang, Y. Determination of the effect of different producing areas and harvest time on the content of rutin in Sophora japonica by high performance liquid chromatography. Lishizhen Med. Mater. Med. Res. 2017, 28, 709–711. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=SZGY201703074&DbName=CJFQ2017 (accessed on 15 August 2023).
- Xiao, D.; Tan, Y.; Liu, X.; Yang, R.; Zhang, W.; He, X.; Wang, K. Effects of different legume species and densities on arbuscular mycorrhizal fungal communities in a karst grassland ecosystem. Sci. Total Environ. 2019, 678, 551–558. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar] [CrossRef]
- Mebius, L. A rapid method for the determination of organic carbon in soil. Anal. Chim. Acta. 1960, 22, 120–124. [Google Scholar] [CrossRef]
- Sheng, H.J.; Tian, L.Y.; Jiang, X.; Wang, X.L. Discussion on determination of available nitrogen in the greenhouse soil with alkali hydrolysis diffusion method. Res. Explor. Lab. 2022, 41, 5–7+35. Available online: https://link.cnki.net/doi/10.19927/j.cnki.syyt.2022.02.002 (accessed on 20 August 2023).
- Moonrungsee, N.; Pencharee, S.; Jakmunee, J. Colorimetric analyzer based on mobile phone camera for determination of available phosphorus in soil. Talanta 2015, 136, 204–209. [Google Scholar] [CrossRef]
- Cox, A.E.; Joern, B.C.; Brouder, S.M.; Gao, D. Plant-available potassium assessment with a modified sodium tetraphenylboron method. Soil Sci. Soc. Am. J. 1999, 63, 902–911. [Google Scholar] [CrossRef]
- Rajtor, M.; Piotrowska-Seget, Z. Prospects for arbuscular mycorrhizal fungi (AMF) to assist in phytoremediation of soil hydrocarbon contaminants. Chemosphere 2016, 162, 105–116. [Google Scholar] [CrossRef]
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L.; et al. The real-time polymerase chain reaction. Mol. Asp. Med. 2006, 27, 95–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.N.; Feng, K.; Li, S.Z.; Wang, Z.J.; Zhang, Z.J.; Deng, Y. In-silico evaluation and improvement on 16S/18S/ITS primers for amplicon high-throughput sequencing. Microbiol. China 2020, 47, 2897–2912. [Google Scholar] [CrossRef]
- Tom, C.J.; Hill, K.A.; Walsh, J.A.; Harris, B.F. Moffett, Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McMahon, S.; Schimel, J. Bacterial and fungal community structure in Arctic tundra tussock and shrub soils. FEMS Microbiol. Ecol. 2007, 59, 428–435. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Lundberg, D.S.; Lazarovits, G.; Reis, V.M.; Raizada, M.N. Bacterial populations in juvenile maize rhizospheres originate from both seed and soil. Plant Soil. 2016, 405, 337–355. [Google Scholar] [CrossRef]
- Dimitriu, P.A.; Grayston, S.J. Relationship between soil properties and patterns of bacterial β-diversity across reclaimed and natural boreal forest soils. Microb. Ecol. 2010, 59, 563–573. [Google Scholar] [CrossRef]
- Kielak, A.M.; Cipriano, M.A.P.; Kuramae, E.E. Acidobacteria strains from subdivision 1 act as plant growth-promoting bacteria. Arch. Microbiol. 2016, 198, 987–993. [Google Scholar] [CrossRef]
- Bastida, F.; Hernández, T.; Albaladejo, J.; García, C. Phylogenetic and functional changes in the microbial community of long-term restored soils under semiarid climate. Soil Biol. Biochem. 2013, 65, 12–21. [Google Scholar] [CrossRef]
- Djemiel, C.; Grec, S.; Hawkins, S. Characterization of bacterial and fungal community dynamics by high-throughput sequencing (HTS) metabarcoding during flax dew-retting. Front. Microbiol. 2017, 8, 294096. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, J.; Wang, C.; Li, W.; Chen, G.; Bai, Y. Altitude distribution characteristics of farmland soil bacteria in loess hilly region of Ningxia. Environ. Sci. 2023, 1–11. [Google Scholar] [CrossRef]
- Zhou, H.; Li, S.; Sun, J.; Qu, J.; Zhang, Z.; Ma, L.; Qin, R.; Wei, J.; Chang, T.; Su, H.; et al. The variation characteristics of plant community and soil physical and chemical properties of alpine meadow along altitude gradient in Sanjiangyuan region. Acta Agrestia Sin. 2023, 31, 1735–1743. Available online: https://kns.cnki.net/kcms/detail/11.3362.S.20230511.1052.004.html (accessed on 2 October 2023).
- Wu, Z.; Lin, W.; Chen, Z.; Fang, C.; Zhang, Z.; Wu, L.; Zhou, M.; Shen, L. Soil microbial community characteristics of different vegetation types in Wuyishan National Nature Reserve. Chin. J. Appl. Ecol. 2013, 24, 2301–2309. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Wu, G.; Xiang, J.; Li, M.; Yu, L. Migration characteristics of main elements in different geological backgrounds and their correlation with plant transpiration in Maocun Village. Guilin. Soils 2007, 5, 746–752. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=TURA200705014&DbName=CJFQ2007 (accessed on 27 October 2023).
- Liang, Y.; Su, Y.; He, X.; Chen, X. Effects of lithology on community structure and abundance of nitrogen-fixing bacteria and arbuscular mycorrhizal fungi in karst shrub soil. Environ. Sci. 2017, 38, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar]
- Jia, S.; Yu, L. Analysis of soil physical and chemical properties of limestone and dolomite in karst rocky desertification area-A case study of Xingyi City, Guizhou Province. Guizhou Sci. 2010, 28, 29–33+55. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=GZKX201003008&DbName=CJFQ2010 (accessed on 2 January 2024).
- Weemstra, M.; Peay, K.G.; Davies, S.J.; Mohamad, M.; Itoh, A.; Tan, S.; Russo, S.E. Lithological constraints on resource economies shape the mycorrhizal composition of a Bornean rain forest. New Phytol. 2020, 228, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, S.; Zhou, D.; Zhang, D.; Li, F.; Zhou, Z.; Xiong, K. Correlation analysis of lithology and land rocky desertification in karst area of Guizhou. Acta Geogr. Sin. 2003, 2, 314–320. Available online: https://kns.cnki.net/kcms2/article/abstract?v=LD-wYsOa3DgxakyuZUjGbLFEaXNOom2CD_BDByTft-eCZsHGhmkhAkVsUgRAIe_yAWhvAE-KasGOv-MX2HketpVSmJjXYYPQtzefKJSaCsfJnd5-1dufmQochb8Nm3rr&uniplatform=NZKPT&language=CHS (accessed on 10 January 2024).
- Wang, G.; Wang, L.; Ma, F. Effects of earthworms and arbuscular mycorrhizal fungi on improvement of fertility and microbial communities of soils heavily polluted by cadmium. Chemosphere 2022, 286, 131567. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Wang, X.; Wang, J.; Liu, G.; Zhang, C. Nitrogen fertilization increases fungal diversity and abundance of saprotrophs while reducing nitrogen fixation potential in a semiarid grassland. Plant Soil. 2021, 465, 515–532. [Google Scholar] [CrossRef]
- Lauro, F.M.; McDougald, D.; Thomas, T.; Williams, T.J.; Egan, S.; Rice, S.; DeMaere, M.Z.; Ting, L.; Ertan, H.; Johnson, J.; et al. The genomic basis of trophic strategy in marine bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 15527–15533. [Google Scholar] [CrossRef] [PubMed]
- Bastida, F.; Torres, I.F.; Moreno, J.L.; Baldrian, P.; Ondoño, S.; Ruiz-Navarro, A.; Hernández, T.; Richnow, H.H.; Starke, R.; García, C.; et al. The active microbial diversity drives ecosystem multifunctionality and is physiologically related to carbon availability in Mediterranean semi-arid soils. Mol. Ecol. 2016, 25, 4660–4673. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China. Soil Tillage Res. 2019, 195, 104355. [Google Scholar] [CrossRef]
- Hou, Y.; Zeng, W.; Hou, M.; Wang, Z.; Luo, Y.; Lei, G.; Zhou, B.; Huang, J. Responses of the soil microbial community to salinity stress in Maize Fields. Biology 2021, 10, 1114. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, J.; Banerjee, S.; Zhou, N.; Zhao, Z.; Zhang, K.; Hu, M.; Tian, C. Biogeographical distribution of bacterial communities in saline agricultural soil. Geoderma 2020, 361, 114095. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, S.; Li, K.; Qiao, J.; Guo, Y.; Liu, Z.; Guo, X. Responses of soil bacterial and fungal communities to the long-term monoculture of grapevine. Appl. Microbiol. Biotechnol. 2021, 105, 7035–7050. [Google Scholar] [CrossRef]
- Tian, M.; Liang, J.; Liu, S.; Yu, R.; Zhang, X. Effects of watermelon cropping management on soil bacteria and fungi biodiversity. Agric 2023, 13, 1010. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Noya, Y.E.; Montoya-Ciriaco, N.; Muñoz-Arenas, L.C.; Hereira-Pacheco, S.; Estrada-Torres, A.; Dendooven, L. Conversion of a high-altitude temperate forest for agriculture reduced alpha and beta diversity of the soil fungal communities as revealed by a metabarcoding analysis. Front. Microbiol. 2021, 12, 667566. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, Z.; Zhou, L.; Huang, K. Effects of altitude and continuous cropping on arbuscular mycorrhizal fungi community in Siraitia grosvenorii rhizosphere. Agric 2023, 13, 1548. [Google Scholar] [CrossRef]
- Yang, W.; Ji, Z.; Wu, A.; He, D.; Rensing, C.; Chen, Y.; Chen, C.; Wu, H.; Muneer, M.; Wu, L. Inconsistent responses of soil bacterial and fungal community’s diversity and network to magnesium fertilization in tea (Camellia sinensis) plantation soils. Appl. Soil Ecol. 2023, 191, 105055. [Google Scholar] [CrossRef]
- Senbayram, M.; Gransee, A.; Wahle, V.; Thiel, H. Role of magnesium fertilisers in agriculture: Plant-soil continuum. Crop Pasture Sci. 2015, 66, 1219. [Google Scholar] [CrossRef]
- Yang, X.; Ni, K.; Shi, Y.; Yi, X.; Zhang, Q.; Fang, L.; Ma, L.; Ruan, J. Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 2018, 252, 74–82. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equipment | Manufacturer | City | Country |
|---|---|---|---|
| pH meter (FE28) | Mettler Toledo | Shanghai | China |
| oil bath (HH-S) | Jiangsu Kexi Instrument | Changzhou | China |
| Kjeldahl nitrogen analyzer (K1160) | Shandong Hanon Scientific Instruments | Dezhou | China |
| Ultraviolet–visible spectrophotometer (UV-1800PC) | Shanghai Mapada Instruments | Shanghai | China |
| flame photometer (FP640) | Shanghai Jingke Electronics | Shanghai | China |
| ICAP (7200) | Thermo Fisher Scientific | Waltham | America |
| Reagents | Concentration |
|---|---|
| K2Cr2O7 | 0.8 mol L−1 |
| H2SO4 | 98% |
| FeSO4 | 0.2 mol L−1 |
| MgCl2 | analytical pure |
| HCl/H2SO4 | 4.0 mL HCl + 0.7 mL H2SO4 |
| NH4OAc | 1 mol L−1 |
| CH3COONH4 | 1 mol L−1 |
| Sample Code | Altitude (m) | Longitude and Latitude | pH | AP (mg kg−1) | AN (g kg−1) | AMg (mg kg−1) | ACa (mg kg−1) | AK (mg kg−1) | SOM (g kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| SY1 | 171.45 | 25°4′8″ 110°18′3″ | 5.99 ± 0.52 a | 35 ± 12 a | 0.09 ± 0.00 a | 66 ± 17 a | 1062 ± 286 a | 175 ± 160 a | 14 ± 0.31 a |
| SY2 | 672.25 | 25°50′19″ 111°12′27″ | 5.89 ± 0.37 a | 55 ± 39 a | 0.16 ± 0.05 a | 55 ± 12 a | 835 ± 126 a | 87 ± 26 a | 21 ± 6.21 a |
| BY1 | 171.45 | 25°53′25″ 111°3′33″ | 5.44 ± 0.12 a | 73 ± 58 b | 0.13 ± 0.01 b | 47 ± 7 a | 698 ± 72 a | 52 ± 8 b | 19 ± 0.38 b |
| BY2 | 230.40 | 25°55′17″ 111°0′52″ | 4.72 ± 0.32 b | 166 ± 121 b | 0.12 ± 0.02 b | 31 ± 17 a | 440 ± 143 a | 145 ± 85 b | 18 ± 4.34 b |
| BY3 | 25°49′26″ 111°2′18″ | 4.62 ± 0.34 b | 825 ± 72 a | 0.21 ± 0.03 a | 49 ± 23 a | 1155 ± 661 a | 298 ± 33 a | 29 ± 3.16 a | |
| SH1 | 319.10 | 25°52′28″ 111°6′1″ | 5.65 ± 0.73 b | 67 ± 42 b | 0.18 ± 0.06 b | 306 ± 14 a | 2432 ± 242 b | 171 ± 125 a | 19 ± 2.01 c |
| SH2 | 438.50 | 25°51′36″ 111°8′8″ | 6.48 ± 0.38 a | 181 ± 90 b | 0.16 ± 0.04 b | 136 ± 65 b | 2279 ± 347 b | 218 ± 119 a | 23 ± 7.17 bc |
| SH3 | 537.40 | 25°53′54″ 111°10′41″ | 4.66 ± 0.34 c | 381 ± 19 a | 0.24 ± 0.10 b | 106 ± 16 b | 544 ± 128 d | 241 ± 79 a | 25 ± 4.71 bc |
| SH4 | 672.25 | 25°49′28″ 111°15′37″ | 6.10 ± 0.25 ab | 182 ± 95 b | 0.35 ± 0.02 a | 115 ± 13 b | 3312 ± 145 a | 188 ± 49 a | 53 ± 0.53 a |
| SH5 | 714.00 | 25°50′19″ 111°12′28″ | 6.51 ± 0.25 a | 195 ± 95 b | 0.19 ± 0.05 b | 93 ± 5 b | 1782 ± 347 c | 142 ± 83 a | 28 ± 5.21 b |
| Sample ID | Phylum | Class | Order | Family | Genus | Species | OTUs |
|---|---|---|---|---|---|---|---|
| SY1 | 33 (11) | 86 (35) | 150 (79) | 206 (153) | 352 (191) | 254 (116) | 6987 (1458) |
| SY2 | 31 (11) | 83 (36) | 154 (80) | 210 (137) | 324 (187) | 227 (93) | 6685 (1588) |
| BY1 | 27 (11) | 80 (35) | 148 (76) | 193 (146) | 311 (190) | 217 (100) | 6529 (1391) |
| BY2 | 30 (11) | 83 (31) | 150 (71) | 203 (130) | 330 (175) | 246 (79) | 6469 (1355) |
| BY3 | 31 (9) | 82 (29) | 156 (69) | 214 (128) | 375 (171) | 274 (101) | 6463 (1208) |
| SH1 | 28 (8) | 85 (31) | 160 (72) | 212 (130) | 380 (167) | 277 (92) | 6516 (1391) |
| SH2 | 28 (11) | 85 (30) | 161 (65) | 223 (121) | 379 (161) | 269 (91) | 6499 (1159) |
| SH3 | 29 (10) | 79 (32) | 142 (74) | 192 (127) | 315 (154) | 221 (86) | 5486 (1219) |
| SH4 | 29 (13) | 84 (35) | 143 (81) | 205 (140) | 319 (185) | 220 (97) | 6303 (1534) |
| SH5 | 34 (10) | 88 (34) | 171 (74) | 220 (134) | 352 (172) | 247 (97) | 6709 (1395) |
| Total | 37 (15) | 98 (46) | 201 (112) | 274 (239) | 531 (359) | 441 (259) | 19,634 (4252) |
| Sample ID | Richness Index | Chao1 Index | Shannon Index | Simpson Index |
|---|---|---|---|---|
| SY1 | 4215 ± 399 a | 4216 ± 399 a | 2.82 ± 0.07 a | 0.01 ± 0.00 a |
| SY2 | 4045 ± 63 a | 4046 ± 63 a | 2.83 ± 0.08 a | 0.01 ± 0.00 a |
| BY1 | 3864 ± 480 a | 3865 ± 480 a | 2.72 ± 0.14 a | 0.01 ± 0.00 a |
| BY2 | 3588 ± 726 a | 3589 ± 726 a | 2.61 ± 0.17 a | 0.01 ± 0.00 a |
| BY3 | 3551 ± 498 a | 3552 ± 498 a | 2.65 ± 0.16 a | 0.01 ± 0.00 a |
| SH1 | 3874 ± 303 a | 3875 ± 303 a | 2.77 ± 0.03 ab | 0.01 ± 0.00 a |
| SH2 | 3760 ± 321 ab | 3761 ± 321 ab | 2.68 ± 0.20 ab | 0.01 ± 0.01 a |
| SH3 | 3137 ± 575 b | 3138 ± 575 b | 2.43 ± 0.34 b | 0.03 ± 0.04 a |
| SH4 | 3813 ± 122 ab | 3814 ± 122 ab | 2.51 ± 0.12 ab | 0.02 ± 0.01 a |
| SH5 | 4047 ± 378 a | 4048 ± 378 a | 2.82 ± 0.09 a | 0.01 ± 0.00 a |
| Sample ID | Richness Index | Chao1 Index | Shannon Index | Simpson Index |
|---|---|---|---|---|
| SY1 | 816 ± 91 a | 817 ± 90 a | 1.62 ± 0.10 a | 0.07 ± 0.01 a |
| SY2 | 926 ± 106 a | 927 ± 106 a | 1.61 ± 0.49 a | 0.13 ± 0.17 a |
| BY1 | 776 ± 78 a | 777 ± 78 a | 1.49 ± 0.20 a | 0.12 ± 0.06 a |
| BY2 | 735 ± 36 a | 736 ± 36 a | 1.66 ± 0.21 a | 0.07 ± 0.04 a |
| BY3 | 676 ± 116 a | 677 ± 115 a | 1.43 ± 0.42 a | 0.15 ± 0.16 a |
| SH1 | 779 ± 123 a | 780 ± 122 a | 1.78 ± 0.12 abc | 0.05 ± 0.02 ab |
| SH2 | 647 ± 37 a | 649 ± 37 a | 1.51 ± 0.22 c | 0.11 ± 0.06 a |
| SH3 | 695 ± 88 a | 696 ± 88 a | 1.54 ± 0.18 bc | 0.09 ± 0.04 ab |
| SH4 | 827 ± 152 a | 829 ± 151 a | 1.88 ± 0.02 a | 0.04 ± 0.01 b |
| SH5 | 797 ± 90 a | 798 ± 90 a | 1.82 ± 0.13 ab | 0.05 ± 0.02 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Zhang, Z.; Liu, P.; Zhou, L.; Tan, S.; Tang, C.; Li, Y. Elevated Altitude and Limestone Soil Promoted Fungal Diversity in Rhizosphere Soil of Sophora japonica. Horticulturae 2024, 10, 441. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050441
Yu L, Zhang Z, Liu P, Zhou L, Tan S, Tang C, Li Y. Elevated Altitude and Limestone Soil Promoted Fungal Diversity in Rhizosphere Soil of Sophora japonica. Horticulturae. 2024; 10(5):441. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050441
Chicago/Turabian StyleYu, Limin, Zhongfeng Zhang, Peiyuan Liu, Longwu Zhou, Shuhui Tan, Chungui Tang, and Ying Li. 2024. "Elevated Altitude and Limestone Soil Promoted Fungal Diversity in Rhizosphere Soil of Sophora japonica" Horticulturae 10, no. 5: 441. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050441