
Development of SSRs Based on the Whole Genome and Screening of Bolting-Resistant SSR Marker in Brassica oleracea L.
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Whole-Genome Sequence of B. oleracea
2.2. SSR Identification
2.3. Genomic SSR Primer Design
2.4. Detection and Transferability of SSR Markers
2.5. Construction of Bolting-Resistant/Bolting-Easy Segregated Population and Agronomic Traits Investigation
2.6. Identification of SSR Markers Linked with Bolting Loci
2.7. Haplotype Analysis with 34 Cabbage Accessions and Sequencing of BolSSR040196
3. Results
3.1. SSR Information Analysis of the Whole Genome of B. oleracea
3.2. SSR Primer Design Based on the Whole Genome of B. oleracea
3.3. Transferability Evaluation of the Genomic SSRs
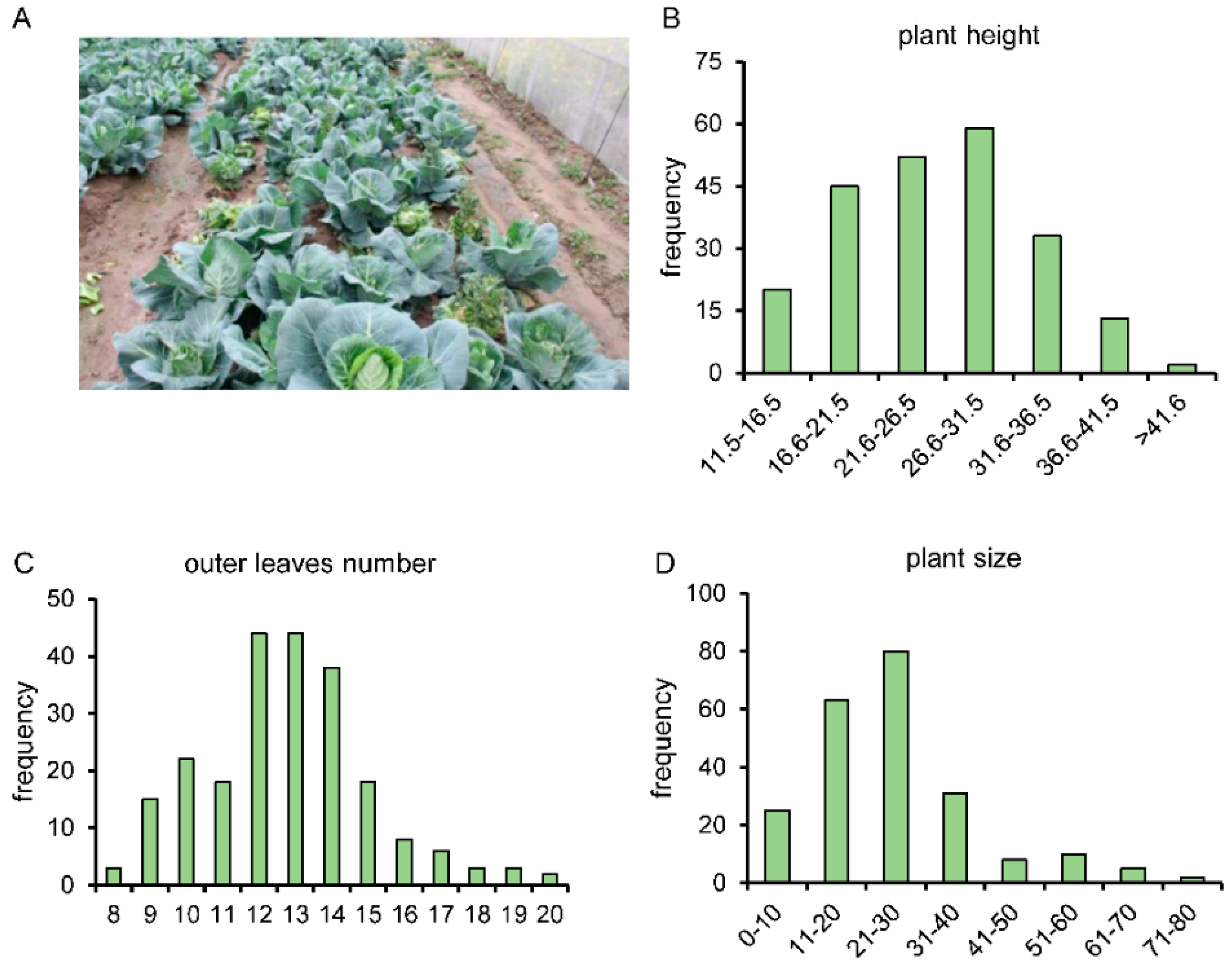
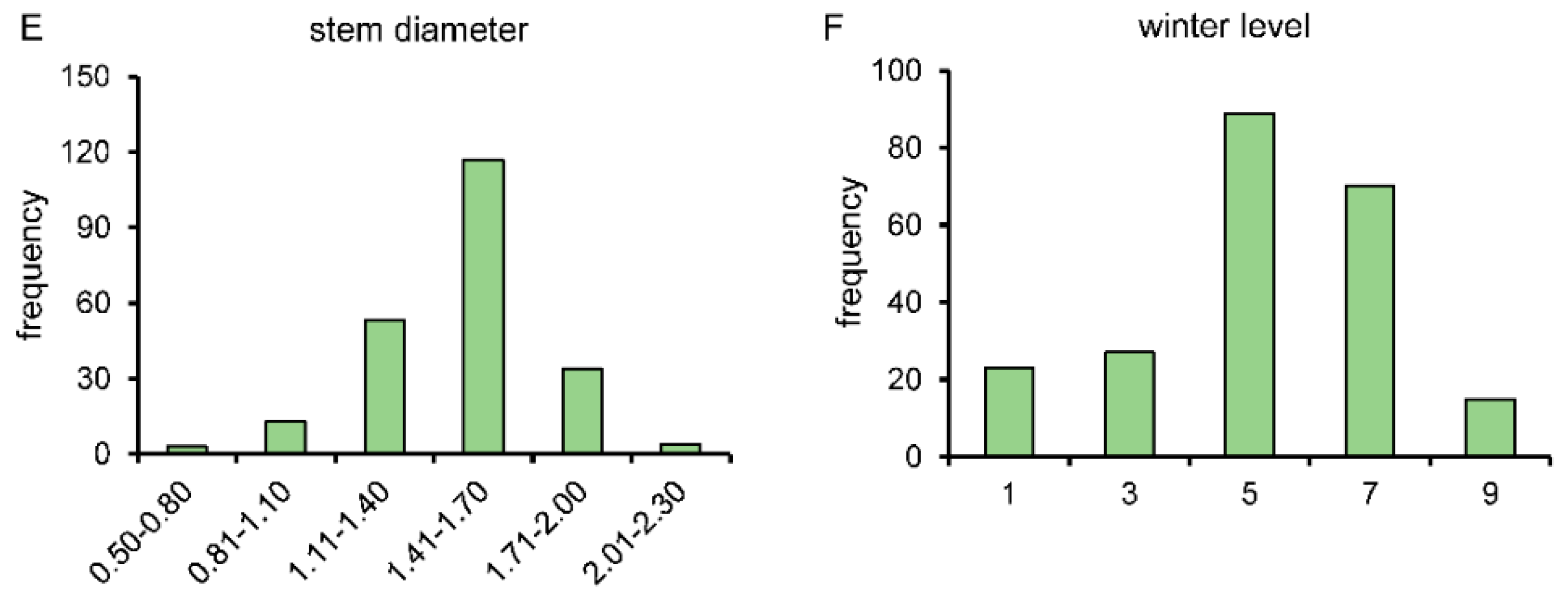
3.4. Construction of BSA Segregated Population and Investigation of Related Traits
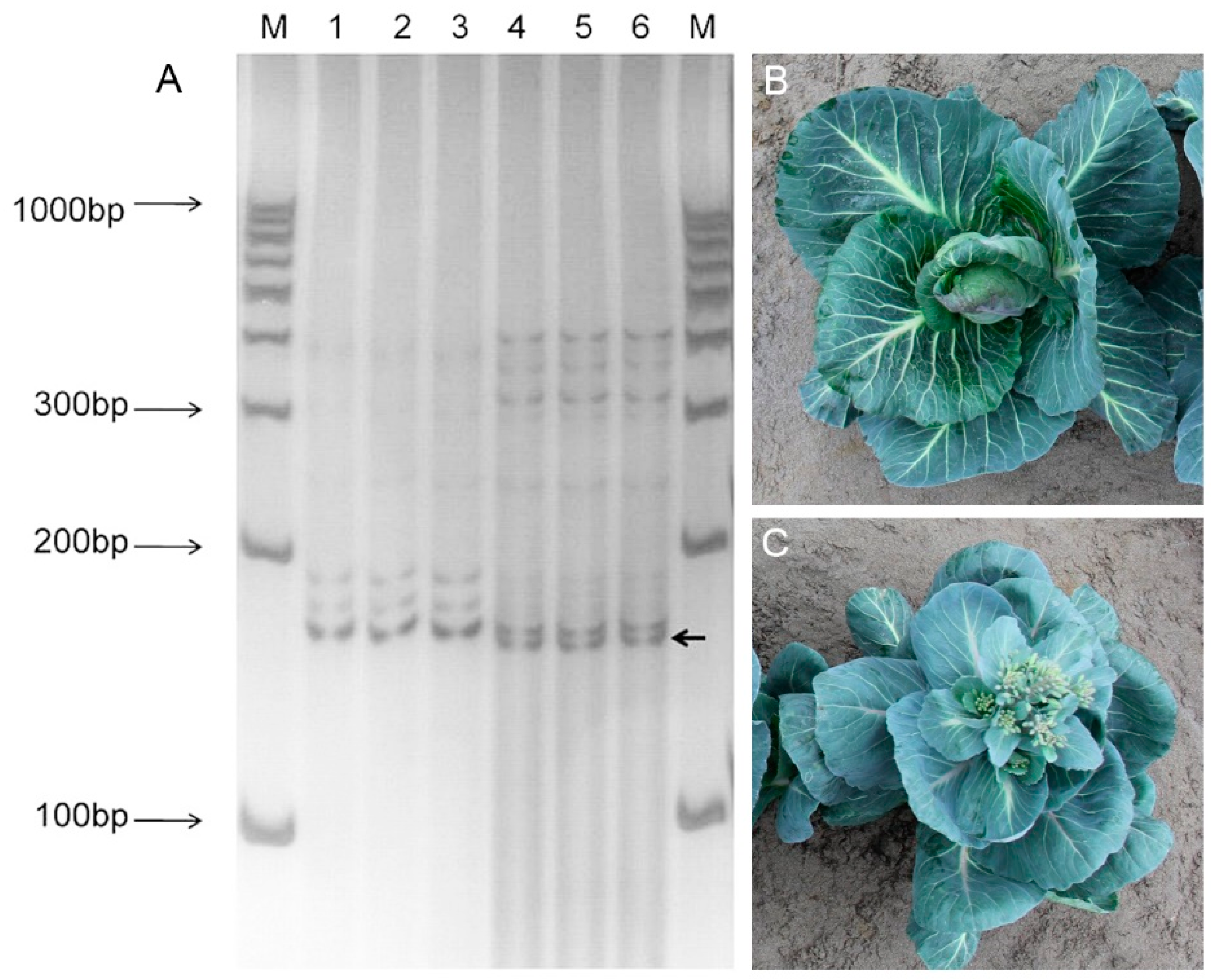
3.5. Screening of Bolting Linkage Markers
3.6. Identification of SSR Markers and Linkage Analysis of BolSSR040196
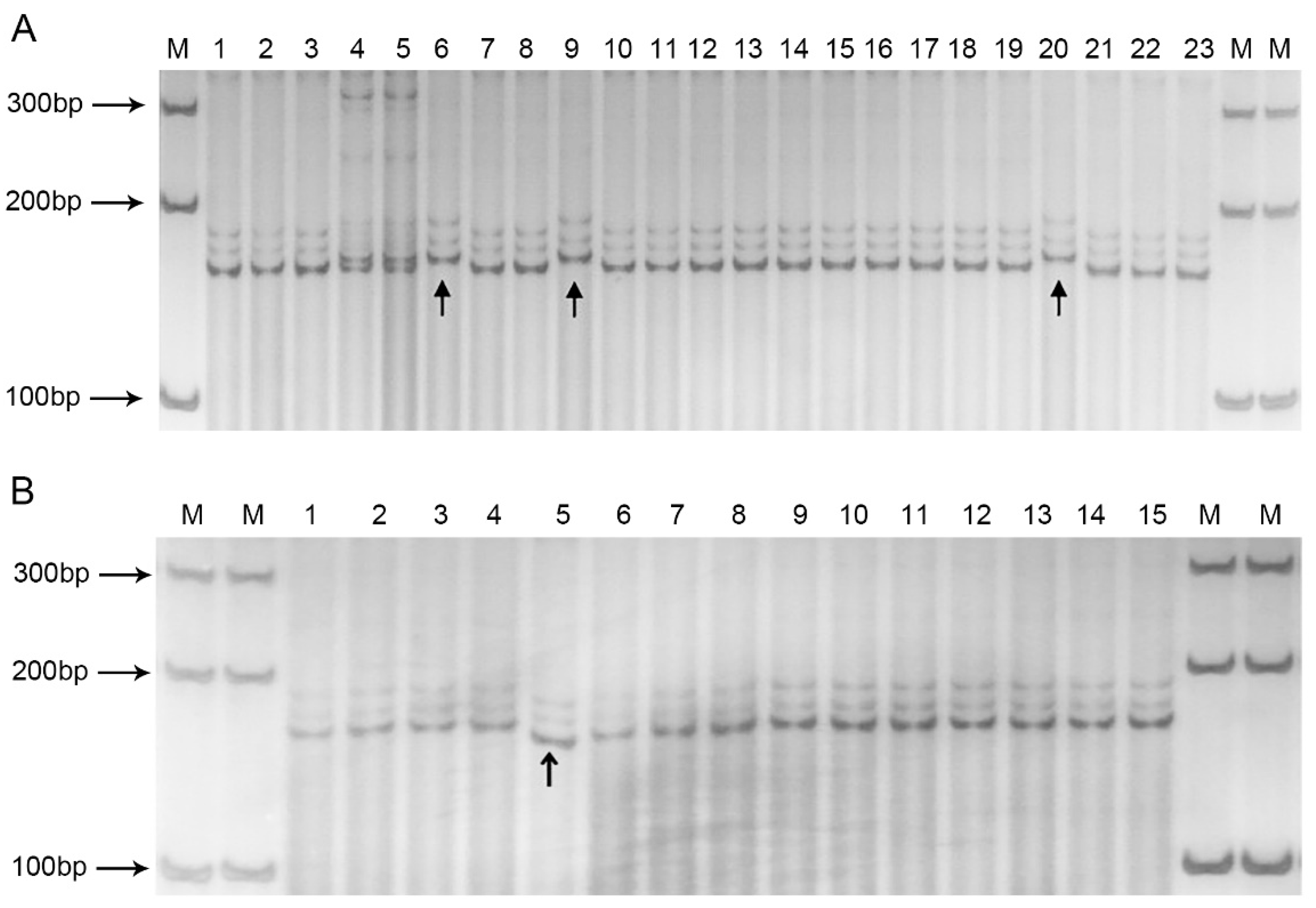
3.7. Haplotype Analysis with 34 Cabbage Accessions
3.8. Sequencing of BolSSR040196
4. Discussion
4.1. Distribution Feature of the SSR on the Whole Genome of B. oleracea
4.2. Application of the Newly Developed Genome-Wide B. oleracea SSR Markers
4.3. The Genetic Linkage Distance between BolSSR040196 and Bolting-Resistant Locus Is 10.69 cM
4.4. Application of the BolSSR040196 Marker in Genetic Improvement of Cabbage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ananga, A.O.; Cebert, E.; Soliman, K.; Kantety, R.; Pacumbaba, R.P.; Konan, K. RAPD markers associated with resistance to blackleg disease in Brassica species. Afr. J. Biotechnol. 2006, 5, 2041–2048. [Google Scholar] [CrossRef]
- Christensen, S.; von Bothmer, R.; Poulsen, G.; Maggioni, L.; Phillip, M.; Andersen, B.A.; Jorgensen, R.B. AFLP analysis of genetic diversity in leafy kale (Brassica oleracea L. convar. acephala (DC.) Alef.) landraces, cultivars and wild populations in Europe. Genet. Resour. Crop Evol. 2010, 58, 657–666. [Google Scholar] [CrossRef]
- Rahman, M.; Li, G.; Schroeder, D.; McVetty, P.B.E. Inheritance of seed coat color genes in Brassica napus (L.) and tagging the genes using SRAP, SCAR and SNP molecular markers. Mol. Breed. 2010, 26, 439–453. [Google Scholar] [CrossRef]
- Zeng, X.H.; Wen, J.; Wan, Z.J.; Yi, B.; Shen, J.X.; Ma, C.Z.; Tu, J.X.; Fu, T.D. Effects of Bleomycin on microspore embryogenesis in Brassica napus and detection of somaclonal variation using AFLP molecular markers. Plant Cell Tiss. Org. 2009, 101, 23–29. [Google Scholar] [CrossRef]
- Panigrahi, J.; Kole, P.; Kole, C. RFLP mapping of loci controlling self-incompatibility in Brassica campestris and their comparative mapping with B. napus and B. oleracea. Biol. Plant. 2011, 55, 54–60. [Google Scholar] [CrossRef]
- Rezaeizad, A.; Wittkop, B.; Snowdon, R.; Hasan, M.; Mohammadi, V.; Zali, A.; Friedt, W. Identification of QTLs for phenolic compounds in oilseed rape (Brassica napus L.) by association mapping using SSR markers. Euphytica 2010, 177, 335–342. [Google Scholar] [CrossRef]
- Shirasawa, K.; Oyama, M.; Hirakawa, H.; Sato, S.; Tabata, S.; Fujioka, T.; Kimizuka-Takagi, C.; Sasamoto, S.; Watanabe, A.; Kato, M.; et al. An EST-SSR linkage map of Raphanus sativus and comparative genomics of the Brassicaceae. DNA Res. 2011, 18, 221–232. [Google Scholar] [CrossRef]
- Xu, Y.; Xing, M.; Song, L.; Yan, J.; Lu, W.; Zeng, A. Genome-wide analysis of simple sequence repeats in cabbage (Brassica oleracea L.). Front. Plant Sci. 2021, 12, 726084. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.J.; Moule, C.; Trick, M.; Edwards, K.J. Efficient large-scale development of microsatellites for marker and mapping applications in Brassica crop species. Theor. Appl. Genet. 2004, 108, 1103–1112. [Google Scholar] [CrossRef]
- Burgess, B.; Mountford, H.; Hopkins, C.J.; Love, C.; Ling, A.E.; Spangenberg, G.C.; Edwards, D.; Batley, J. Identification and characterization of simple sequence repeat (SSR) markers derived in silico from Brassica oleracea genome shotgun sequences. Mol. Ecol. Notes 2006, 6, 1191–1194. [Google Scholar] [CrossRef]
- Xu, J.S.; Qian, X.J.; Wang, X.F.; Li, R.Y.; Cheng, X.M.; Yang, Y.A.; Fu, J.; Zhang, S.C.; King, G.J.; Wu, J.S.; et al. Construction of an integrated genetic linkage map for the a genome of Brassica napus using SSR markers derived from sequenced BACs in B. rapa. BMC Genom. 2010, 11, 594. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.M.; Xu, J.S.; Xia, S.; Gu, J.X.; Yang, Y.; Fu, J.; Qian, X.J.; Zhang, S.C.; Wu, J.S.; Liu, K.D. Development and genetic mapping of microsatellite markers from genome survey sequences in Brassica napus. Theor. Appl. Genet. 2009, 118, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Li, H.T.; Chen, X.; Yang, Y.; Xu, J.S.; Gu, J.X.; Fu, J.; Qian, X.J.; Zhang, S.C.; Wu, J.S.; Liu, K.D. Development and genetic mapping of microsatellite markers from whole genome shotgun sequences in Brassica oleracea. Mol. Breed. 2011, 28, 585–596. [Google Scholar] [CrossRef]
- McCouch, S.R.; Teytelman, L.; Xu, Y.B.; Lobos, K.B.; Clare, K.; Walton, M.; Fu, B.Y.; Maghirang, R.; Li, Z.K.; Xing, Y.Z.; et al. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 2002, 9, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.J.; Shi, J.R.; Singh, S.; Fickus, E.W.; Costa, J.M.; Lewis, J.; Gill, B.S.; Ward, R.; Cregan, P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theor. Appl. Genet. 2005, 110, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.C.; Grant, D.; Olson, T.; Warren, W.C.; Wing, R.; Yu, Y.; Kim, H.; Cregan, P.; Joseph, B.; Futrell-Griggs, M.; et al. Microsatellite discovery from BAC end sequences and genetic mapping to anchor the soybean physical and genetic maps. Genome 2008, 51, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.C.; Wang, Y.; Huang, L.S.; Wang, Y.J.; Yu, Y.J.; Yang, L. Large-scale development of SSR markers in the genome of cacao. J. Shandong Agric. Univ. Nat. Sci. 2013, 44, 340–344. [Google Scholar]
- Cai, B.; Li, C.H.; Yao, Q.H.; Zhou, J.; Tao, J.M.; Zhang, Z. Analysis of SSRs in grape genome and development of SSRs database. J. Nanjing Agric. Univ. 2009, 32, 28–32. [Google Scholar]
- Qu, J.T.; Liu, J. A genome-wide analysis of simple sequence repeats in maize and the development of polymorphism markers from next-generation sequence data. BMC Res. Notes 2013, 6, 403. [Google Scholar] [CrossRef]
- Jia, H.M.; Shen, Y.T.; Jiao, Y.; Wang, G.Y.; Dong, X.; Jia, H.J.; Du, F.; Liang, S.M.; Zhou, C.C.; Mao, W.H.; et al. Development of 107 SSR markers from whole genome shotgun sequences of Chinese bayberry (Myrica rubra) and their application in seedling identification. J. Zhejiang Univ. Sci. B 2014, 15, 997–1005. [Google Scholar] [CrossRef]
- La Rota, M.; Kantety, R.V.; Yu, J.K.; Sorrells, M.E. Nonrandom distribution and frequencies of genomic and EST-derived microsatellite markers in rice, wheat, and barley. BMC Genom. 2005, 6, 23. [Google Scholar] [CrossRef]
- Cardle, L.; Ramsay, L.; Milbourne, D.; Macaulay, M.; Marshall, D.; Waugh, R. Computational and experimental characterization of physically clustered simple sequence repeats in plants. Genetics 2000, 156, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.H.; Alabady, M.S.; Ulloa, M.; Sickler, B.; Wilkins, T.A.; Yu, J.; Stelly, D.M.; Kohel, R.J.; El-Shihy, O.M.; Cantrell, R.G. Genetic mapping of new cotton fiber loci using EST-derived microsatellites in an interspecific recombinant inbred line cotton population. Mol. Genet. Genom. 2005, 274, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.Q.; Chen, X.P.; Hong, Y.B.; Liu, H.Y.; Zhou, G.Y.; Li, S.X.; Guo, B.Z. Utility of EST-derived SSR in cultivated peanut (Arachis hypogaea L.) and Arachis wild species. BMC Plant Biol. 2009, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Gopalakrishna, T. Development of unigene-derived SSR markers in cowpea (Vigna unguiculata) and their transferability to other Vigna species. Genome 2010, 53, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Huang, S.; Zhan, J.; Yu, J.; Wang, X.; Hua, W.; Liu, S.; Liu, G.; Wang, H. Genome-wide microsatellite characterization and marker development in the sequenced Brassica crop species. DNA Res. 2014, 21, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Boudry, P.; Wieber, R.; Saumitou-Laprade, P.; Pillen, K.; Dijk, H.V.; Jung, C.J.T.; Genetics, A. Identification of RFLP markers closely linked to the bolting gene B and their significance for the study of the annual habit in beets (Beta vulgaris L.). Theor. Appl. Genet. 1994, 88, 852–858. [Google Scholar] [CrossRef]
- El-Mezawy, A.; Dreyer, F.; Jacobs, G.; Jung, C. High-resolution mapping of the bolting gene B of sugar beet. Theor. Appl. Genet. 2002, 105, 100–105. [Google Scholar] [CrossRef]
- Teutonico, R.A.; Osborn, T.C. Mapping loci controlling vernalization requirement in Brassica rapa. Theor. Appl. Genet. 1995, 91, 1279–1283. [Google Scholar] [CrossRef]
- Rosental, L.; Still, D.W.; You, Y.; Hayes, R.J.; Simko, I. Mapping and identification of genetic loci affecting earliness of bolting and flowering in lettuce. Theor. Appl. Genet. 2021, 134, 3319–3337. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Huang, S.N.; Gao, Y.; Zhang, M.D.; Qu, G.Y.; Wang, N.; Liu, Z.Y.; Feng, H. Role of BrSDG8 on bolting in Chinese cabbage (Brassica rapa). Theor. Appl. Genet. 2020, 133, 2937–2948. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Wang, J.L.; Li, B.Y.; Wei, Q.Z.; Hu, T.H.; Hu, H.J.; Bao, C.L. Genetic and QTL Mapping Analysis of Bolting Time in Cabbage(Brassica oleracea). Acta Hortic. Sin. 2020, 47, 974–982. [Google Scholar] [CrossRef]
- Wei, X.C.; Rahim, M.A.; Zhao, Y.Y.; Yang, S.J.; Su, H.N.; Wang, Z.Y.; Shahriar, S.A.; Li, J.D.; Yang, Z.Y.; Yuan, Y.X.; et al. Inheritance and Genetic Mapping of Late-Bolting to Early-Bolting Gene, BrEb-1, in Chinese Cabbage (Brassica rapa L.). Agronomy 2022, 12, 1048. [Google Scholar] [CrossRef]
- Zhu, X.W.; Bo, T.Y.; Chen, J.X.; Tai, X.; Ren, Y.Y. Research Progress on Mapping of Genes Associated with Main Agronomic Traits of Cabbage. Acta Hortic. Sin. 2017, 44, 1729–1737. [Google Scholar] [CrossRef]
- Du, L.; Zhang, C.; Liu, Q.; Zhang, X.; Yue, B.; Hancock, J. Krait: An ultrafast tool for genome-wide survey of microsatellites and primer design. Bioinformatics 2018, 34, 681–683. [Google Scholar] [CrossRef]
- da Maia, L.C.; Palmieri, D.A.; de Souza, V.Q.; Kopp, M.M.; de Carvalho, F.I.; Costa de Oliveira, A. SSR Locator: Tool for Simple Sequence Repeat Discovery Integrated with Primer Design and PCR Simulation. Int. J. Plant Genom. 2008, 2008, 412696. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W.; Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- An, Z.S.; Gao, C.H.; Li, J.N.; Fu, D.H.; Tang, Z.L.; Ortegon, O. Large-scale development of functional markers in Brassica species. Genome 2011, 54, 763–770. [Google Scholar] [CrossRef]
- Sim, S.C.; Yu, J.K.; Jo, Y.K.; Sorrells, M.E.; Jung, G. Transferability of cereal EST-SSR markers to ryegrass. Genome 2009, 52, 431–437. [Google Scholar] [CrossRef]
- Castillo, A.; Budak, H.; Varshney, R.K.; Dorado, G.; Graner, A.; Hernandez, P. Transferability and polymorphism of barley EST-SSR markers used for phylogenetic analysis in Hordeum chilense. BMC Plant Biol. 2008, 8, 97. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.M.; Hou, X.L.; Dong, Y.X. Development of SSR Primers of Non-heading Chinese Cabbage and Transferability among Closely Related Species. Sci. Technol. Rev. 2005, 23, 20–25. [Google Scholar]
- Chen, C.; Zhuang, M.; Li, K.N.; Liu, Y.M.; Yang, L.M.; Zhang, Y.Y.; Cheng, F.; Sun, P.T.; Fang, Z.Y. Development and Utility of EST-SSR Marker in Cabbage. Acta Hortic. Sin. 2010, 37, 221–228. [Google Scholar] [CrossRef]
- Chen, C.; Zhuang, M.; Fang, Z.Y.; Wang, Q.B.; Zhang, Y.Y.; Liu, Y.M.; Yang, L.M.; Cheng, F. A Co-Dominant Marker BoE332 Applied to Marker-Assisted Selection of Homozygous Male-Sterile Plants in Cabbage (Brassica oleracea var. capitata L.). J. Integr. Agr. 2013, 12, 596–602. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Y.; Li, Z.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y. QTL Analysis of Head Splitting Resistance in Cabbage (Brassica oleracea L. var. capitata) Using SSR and InDel Makers Based on Whole-Genome Re-Sequencing. PLoS ONE 2015, 10, e0138073. [Google Scholar] [CrossRef]
- Cao, W.R.; Wang, C. RAPD Marker of Later Bolting Gene on Cabbage. Biotechnol. Bull. 2007, 5, 167–169. [Google Scholar] [CrossRef]
- Mao, W.W.; Gao, H.S.; Bo, T.Y.; Ma, J.J.; Xu, D.J.; Chen, X.H.; Jia, Z.M.; Wang, Y.L. Analysis of Bolting Trait of Brassica campestris L. ssp. Chinensis var. utilis Tsen et Lee By ISSR and SRAP Markers. Jiangsu J. Agric. Sci. 2009, 25, 829–833. [Google Scholar]
- Tränkner, C.; Pfeiffer, N.; Kirchhoff, M.; Kopisch-Obuch, F.J.; van Dijk, H.; Schilhabel, M.; Hasler, M.; Emrani, N. Deciphering the complex nature of bolting time regulation in Beta vulgaris. Theor. Appl. Genet. 2017, 130, 1649–1667. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Liu, Z.Y.; Feng, C.D.; Zhang, H.L.; Xu, Z.S.; Wang, X.W.; Wu, J.; She, H.B.; Qian, W. Quantitative trait locus mapping and identification of candidate genes controlling bolting in spinach (Spinacia oleracea L.). Front. Plant Sci. 2022, 13, 850810. [Google Scholar] [CrossRef] [PubMed]
- Wako, T.; Tsukazaki, H.; Yaguchi, S.; Yamashita, K.; Ito, S.; Shigyo, M. Mapping of quantitative trait loci for bolting time in bunching onion (Allium fistulosum L.). Euphytica 2016, 209, 537–546. [Google Scholar] [CrossRef]
- Wang, Q.B.; Wang, Y.P.; Zhang, L. Inheritance and Molecular Marker for Flowering Time in Radish (Raphanus sativus L.). Plant Mol. Biol. Rep. 2018, 36, 878–887. [Google Scholar] [CrossRef]
- Dai, Y.; Sun, X.; Wang, C.G.; Li, F.; Zhang, S.F.; Zhang, H.; Li, G.L.; Yuan, L.Y.; Chen, G.H.; Sun, R.F.; et al. Gene co-expression network analysis reveals key pathways and hub genes in Chinese cabbage (Brassica rapa L.) during vernalization. BMC Genom. 2021, 22, 236. [Google Scholar] [CrossRef]
- Hedden, P.; Sponsel, V. A century of gibberellin research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.H.; Ren, X.D.; Mason, A.S.; Li, J.N.; Wang, W.; Xiao, M.L.; Fu, D.H. Revisiting an important component of plant genomes: Microsatellites. Funct. Plant Biol. 2013, 40, 645–661. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSR | B. rapa | B. nigra | B. oleracea | B. juncea | B. napus | B. carinata | R. sativus |
|---|---|---|---|---|---|---|---|
| BolSSR000026 | + | + | + | − | − | + | − |
| BolSSR001052 | + | + | + | + | − | + | + |
| BolSSR002169 | + | + | + | + | − | + | − |
| BolSSR002506 | + | + | + | + | − | + | + |
| BolSSR003205 | + | + | + | + | − | + | + |
| BolSSR004460 | + | + | + | + | − | + | − |
| BolSSR007539 | + | + | + | + | + | + | + |
| BolSSR007971 | + | + | + | + | + | + | + |
| BolSSR008152 | + | + | + | + | − | + | + |
| BolSSR009441 | + | + | + | + | + | + | + |
| BolSSR009823 | − | + | + | + | + | + | + |
| BolSSR014499 | + | + | + | + | + | + | − |
| BolSSR057533 | + | − | + | − | + | + | − |
| BolSSR016035 | − | − | − | + | − | + | − |
| BolSSR017491 | − | + | + | + | + | + | + |
| BolSSR018633 | + | + | + | + | − | + | + |
| BolSSR021757 | + | + | + | + | + | + | + |
| BolSSR024526 | + | − | − | − | − | − | − |
| BolSSR027265 | + | + | + | + | + | + | + |
| BolSSR028997 | + | − | + | − | + | + | − |
| BolSSR029692 | + | − | + | − | + | + | − |
| BolSSR035058 | − | + | + | + | − | + | + |
| BolSSR031249 | + | + | + | + | + | + | + |
| BolSSR038129 | + | + | + | + | + | + | + |
| BolSSR039332 | + | + | + | + | − | + | + |
| BolSSR039798 | + | + | + | + | + | + | − |
| BolSSR040726 | + | + | + | + | + | + | + |
| BolSSR044427 | + | + | + | + | − | + | + |
| BolSSR045579 | + | + | + | + | + | + | − |
| BolSSR030903 | + | − | + | + | + | + | − |
| BolSSR059526 | + | − | + | + | + | − | − |
| BolSSR060281 | − | − | + | − | + | + | − |
| BolSSR061903 | + | + | + | + | + | − | + |
| BolSSR056868 | + | + | + | + | + | + | + |
| BolSSR063024 | + | + | + | + | − | + | + |
| BolSSR055078 | + | + | + | + | + | − | − |
| BolSSR057239 | + | + | + | + | + | − | + |
| BolSSR053957 | + | + | + | + | − | + | + |
| BolSSR051866 | + | + | + | + | − | + | − |
| BolSSR052736 | + | + | + | + | + | + | − |
| BolSSR065129 | + | + | + | + | + | − | + |
| BolSSR064292 | − | − | + | + | − | + | − |
| BolSSR066510 | + | + | + | + | + | + | − |
| BolSSR066830 | − | − | + | − | + | + | − |
| BolSSR070186 | + | + | + | + | + | + | + |
| BolSSR071665 | + | − | + | + | − | + | + |
| BolSSR072679 | + | + | + | + | − | + | + |
| BolSSR073675 | + | + | + | + | + | + | + |
| BolSSR076246 | + | + | + | + | + | + | − |
| BolSSR078112 | + | + | + | + | + | − | + |
| BolSSR064485 | + | + | + | + | + | + | + |
| BolSSR080017 | + | + | + | + | + | + | + |
| BolSSR081968 | + | + | + | + | − | + | + |
| BolSSR082001 | + | + | + | + | + | + | + |
| Accession No. | Actual Phenotype | Result of Validation | Accession No. | Actual Phenotype | Result of Validation |
|---|---|---|---|---|---|
| C27 | easy | resistant | C70 | easy | easy |
| C28 | easy | easy | C71 | easy | easy |
| C32 | resistant | resistant | C79 | easy | easy |
| C33 | resistant | resistant | C80 | easy | easy |
| C35 | easy | easy | C81 | easy | easy |
| C40 | resistant | resistant | C88 | easy | easy |
| C44 | resistant | resistant | C93 | resistant | resistant |
| C50 | easy | resistant | C95 | easy | easy |
| C51 | resistant | resistant | C96 | easy | resistant |
| C53 | resistant | easy | C103 | easy | easy |
| C54 | resistant | resistant | C110 | easy | resistant |
| C55 | resistant | resistant | C128 | easy | easy |
| C56 | resistant | resistant | C129 | easy | easy |
| C57 | resistant | resistant | C131 | easy | easy |
| C59 | resistant | resistant | C133 | easy | easy |
| C62 | easy | easy | C134 | easy | easy |
| C65 | easy | easy | C135 | resistant | resistant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, T.; Miao, L.; Zou, M.; Hussain, I.; Yu, H.; Li, J.; Sun, N.; Kong, L.; Wang, S.; Li, J.; et al. Development of SSRs Based on the Whole Genome and Screening of Bolting-Resistant SSR Marker in Brassica oleracea L. Horticulturae 2024, 10, 443. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050443
Zhao T, Miao L, Zou M, Hussain I, Yu H, Li J, Sun N, Kong L, Wang S, Li J, et al. Development of SSRs Based on the Whole Genome and Screening of Bolting-Resistant SSR Marker in Brassica oleracea L. Horticulturae. 2024; 10(5):443. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050443
Chicago/Turabian StyleZhao, Tong, Liming Miao, Minghua Zou, Iqbal Hussain, Hongrui Yu, Jia Li, Nairan Sun, Lijun Kong, Shenyun Wang, Jianbin Li, and et al. 2024. "Development of SSRs Based on the Whole Genome and Screening of Bolting-Resistant SSR Marker in Brassica oleracea L." Horticulturae 10, no. 5: 443. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050443