

Toxicological Characteristics of Bacterial Nanocellulose in an In Vivo Experiment—Part 1: The Systemic Effects

 ,
,  ,
,  , ,
, ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples of BNC and Methods of Its Characterization
2.2. Experimental Design
2.3. Analytical Methods
2.4. Statistical Processing of Experimental Data
3. Results
3.1. Microbiome Characterization of Symbiotic Culture Medusomyces Gisevii
3.2. Characterization of BNC Sample
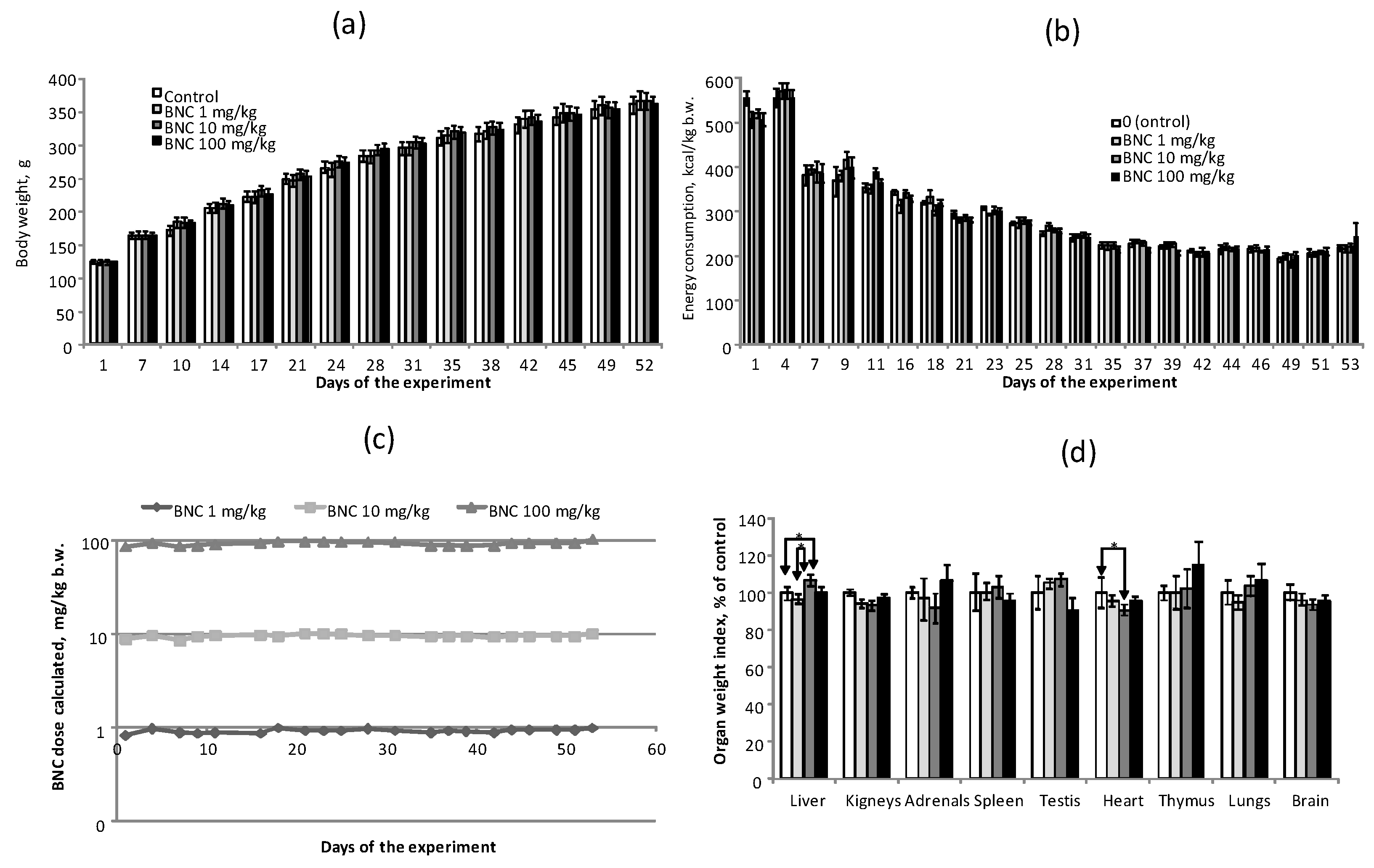
3.3. Integral Indicators in a Subacute 8-Week Experiment
3.4. Influence of BNC on the Behavioral Reactions in Rats
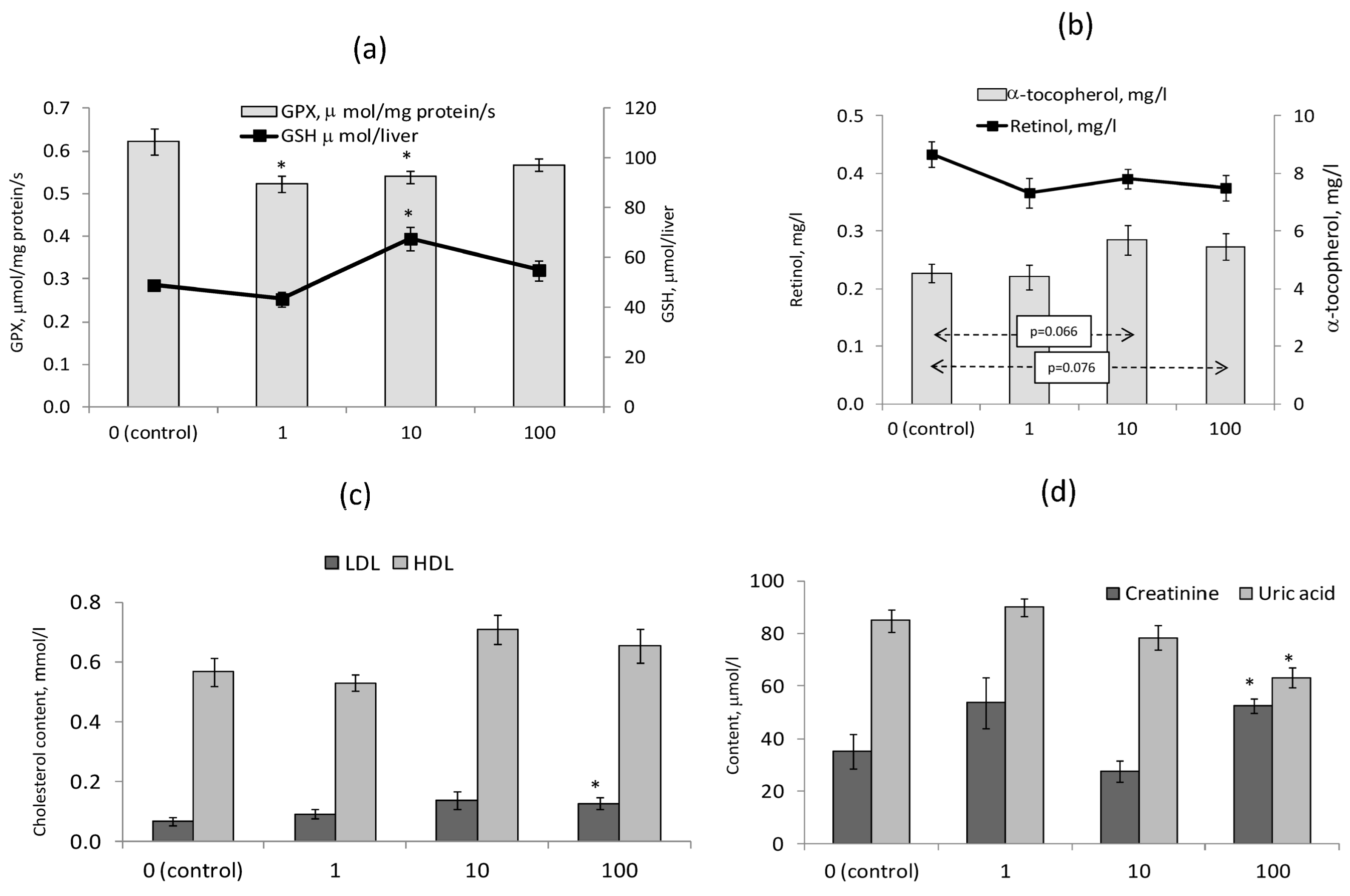
3.5. Blood Biochemical Indicators and the Indices of Fat-Soluble Vitamins Safety
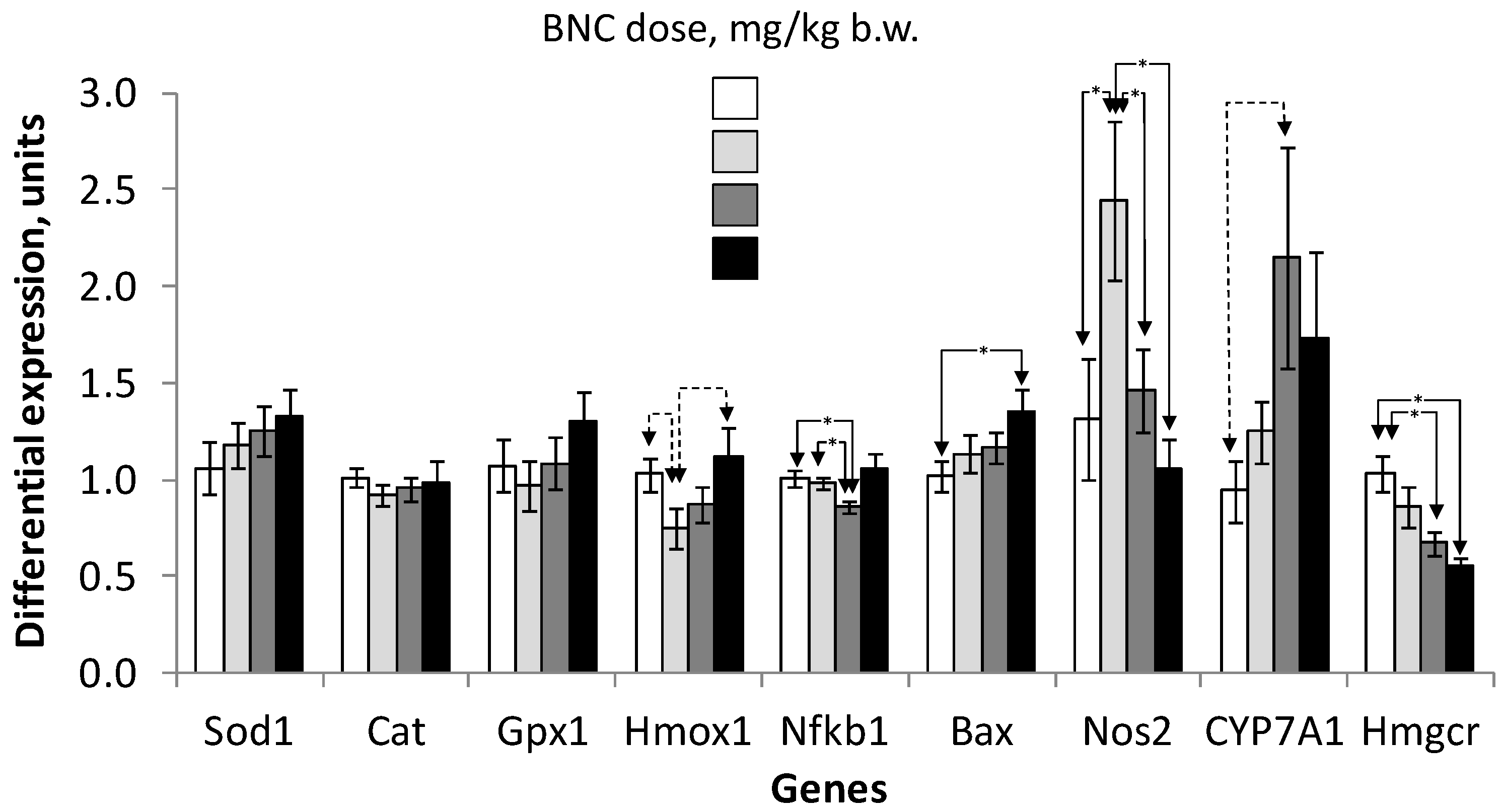
3.6. Morphological Indicators and Gene Expression in the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azeredo, H.M.C.; Barud, H.; Farinas, C.S.; Vasconcellos, V.M.; Claro, A.M. Bacterial cellulose as a raw material for food and food packaging applications. Front. Sustain. Food Syst. 2019, 3, 7. [Google Scholar] [CrossRef]
- Thomas, P.; Duolikun, T.; Rumjit, N.P.; Moosavi, S.; Lai, C.W.; Bin Johan, M.R.; Fen, L.B. Comprehensive review on nanocellulose: Recent developments, challenges and future prospects. J. Mech. Behav. Biomed. Mater. 2020, 110, 103884. [Google Scholar] [CrossRef] [PubMed]
- Portela da Gama, F.M.; Dourado, F. Bacterial nanocellulose: What future? Bioimpacts 2018, 8, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Dourado, F.; Leal, M.; Martins, D.; Fontao, A.; Rodrigues, A.C.; Gama, M. Chapter 7: Celluloses as food ingredients/additives: Is there room for BNC. In Bacterial Nanocellulose, from Biotechnology to Bio-Economy, 1st ed.; Gama, M., Dourado, F., Bielecki, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 123–132. ISBN 9780444634665. [Google Scholar]
- Shi, Z.; Zhang, Y.; Phillips, G.O.; Yang, G. Utilization of bacterial cellulose in food. Food Hydrocoll. 2014, 35, 539–545. [Google Scholar] [CrossRef]
- Chau, C.F.; Yang, P.; Yu, C.M.; Yen, G.C. Investigation on the lipid- and cholesterol-lowering abilities of biocellulose. J. Agric. Food Chem. 2008, 56, 2291–2295. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP); Bampidis, V.; Azimonti, G.; de Lourdes Bastos, M.; Christensen, H.; Dusemund, B.; Durjava, M.K.; Kouba, M.; Lopez-Alonso, M.; Puente, L.; et al. Safety and efficacy of microcrystalline cellulose for all animal species. EFSA J. 2020, 18, e06209. [Google Scholar] [CrossRef]
- Brand, W.; van Kesteren, P.C.E.; Swart, E.; Oomen, A.G. Overview of potential adverse health effects of oral exposure to nanocellulose. Nanotoxicology 2022, 16, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.F.; Frankos, V.H.; Westland, J.; Zoetis, T. Toxicologic evaluation of cellulon fiber; genotoxicity, pyrogenicity, acute and subchronic toxicity. J. Am. Coll. Toxicol. 1991, 10, 541–554. [Google Scholar] [CrossRef]
- Pinto, F.C.M.; De-Oliveira, A.C.A.X.; De-Carvalho, R.R.; Gomes-Carneiro, M.R.; Coelho, D.R.; Lima, S.V.C.; Paumgartten, F.J.R.; Aguiar, J.L.A. Acute toxicity, cytotoxicity, genotoxicity and antigenotoxic effects of a cellulosic exopolysaccharide obtained from sugarcane molasses. Carbohydr. Polym. 2016, 137, 556–560. [Google Scholar] [CrossRef]
- Pértile, R.A.; Moreira, S.; Gil da Costa, R.M.; Correia, A.; Guãrdao, L.; Gartner, F.; Vilanova, M.; Gama, M. Bacterial cellulose: Long-term biocompatibility studies. J. Biomater. Sci. Polym. Ed. 2012, 23, 1339–1354. [Google Scholar] [CrossRef]
- Hagiwara, A.; Imai, N.; Sano, M.; Kawabe, M.; Tamano, S.; Kitamura, S.; Omoto, T.; Asai, I.; Yasuhara, K.; Hayashi, S. A 28-day oral toxicity study of fermentation-derived cellulose, produced by Acetobacter aceti subspecies xylinum, in F344 rats. J. Toxicol. Sci. 2010, 35, 317–325. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Dusemund, B.; Filipic, M.; Frutos, M.J.; Galtier, P.; et al. Re-evaluation of celluloses E 460(i), E 460(ii), E 461, E 462, E 463, E 464, E 465, E 466, E 468 and E 469 as food additives. EFSA J. 2018, 16, e05047. [Google Scholar] [CrossRef] [PubMed]
- Cubas, A.L.V.; Provin, A.P.; Dutra, A.R.A.; Mouro, C.; Gouveia, I.C. Advances in the production of biomaterials through kombucha using food waste: Concepts, challenges, and potential. Polymers 2023, 15, 1701. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef]
- Goh, W.N.; Rosma, A.; Kaur, B.; Fazilah, A.; Karim, A.A.; Rajeev, B. Fermentation of black tea broth (Kombucha): I. Effects of sucrose concentration and fermentation time on the yield of microbial cellulose. Int. Food Res. J. 2012, 19, 109–117. [Google Scholar]
- Campano, C.; Balea, A.; Blanco, A.; Negro, C. Enhancement of the fermentation process and properties of bacterial cellulose: A review. Cellulose 2016, 23, 57–91. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Mende, D.R.; Letunic, I.; Maistrenko, O.M.; Schmidt, T.S.B.; Milanese, A.; Paoli, L.; Hernández-Plaza, A.; Orakov, A.N.; Forslund, S.K.; Sunagawa, S.; et al. proGenomes2: An improved database for accurate and consistent habitat, taxonomic and functional annotations of prokaryotic genomes. Nucleic Acids Res. 2019, 48, D621–D625. [Google Scholar] [CrossRef]
- Shavyrkina, N.A.; Budaeva, V.V.; Skiba, E.A.; Mironova, G.F.; Bychin, N.V.; Gismatulina, Y.A.; Kashcheyeva, E.I.; Sitnikova, A.E.; Shilov, A.I.; Kuznetsov, P.S.; et al. Scale-up of biosynthesis process of bacterial nanocellulose. Polymers 2021, 13, 1920. [Google Scholar] [CrossRef] [PubMed]
- Sedova, I.B.; Zakharova, L.P.; Kiseleva, M.G.; Chalyy, Z.A.; Timonin, A.N.; Aristarkhova, T.V.; Tutelyan, V.A. Deoxynivalenol as a risk factor causing food grain contamination: Monitoring over crops in 1989–2018 in Russian Federation. Health Risk Anal. 2021, 3, 85–98. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals; Institute for Laboratory Animal Research (ILAR); Division on Earth and Life Studies (DELS); National Research Council of the National Academies. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Apryatin, S.A.; Shipelin, V.A.; Trusov, N.V.; Mzhelskaya, K.V.; Evstratova, V.S.; Kirbaeva, N.V.; Soto, J.S.; Fesenko, Z.S.; Gainetdinov, R.R.; Gmoshinski, I.V. Comparative analysis of the influence of a high-fat/high-carbohydrate diet on the level of anxiety and neuromotor and cognitive functions in Wistar and DAT-KO rats. Physiol. Rep. 2019, 7, e13987. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Vrzhesinskaya, O.A.; Beketova, N.A.; Kosheleva, O.V.; Kudan, P.V.; Evstratova, A.D.; Kodentsova, V.M.; Gmoshinksy, I.V. Indicators of vitamin supply in experimental nutritional hyperlipidemia in rodents. Vopr. Pitan. 2017, 1, 6–16. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Trusov, N.V.; Semin, M.O.; Shipelin, V.A.; Apryatin, S.A.; Gmoshinski, I.V. Gene expression in the liver of rats fed with the diet a complex of resveratrol and L-carnitine in normal and obese conditions. Vopr. Pitan. 2021, 5, 26–37. (In Russian) [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Polymenakou, P.N.; Nomikou, P.; Hannington, M.; Petersen, S.; Kilias, S.P.; Anastasiou, T.I.; Papadimitriou, V.; Zaka, E.; Kristoffersen, J.B.; Lampridou, D.; et al. Taxonomic diversity of microbial communities in sub-seafloor hydrothermal sediments of the active Santorini-Kolumbo volcanic field. Front. Microbiol. 2023, 14, 1188544. [Google Scholar] [CrossRef]
- Hasanin, M.S.; Abdelraof, M.; Hashem, A.H.; El Saied, H. Sustainable bacterial cellulose production by Achromobacter using mango peel waste. Microb. Cell Factories 2023, 22, 24. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yukphan, P.; Vu, H.T.L.; Muramatsu, Y.; Ochaikul, D.; Nakagawa, Y. Subdivision of the genus Gluconacetobacter Yamada, Hoshino and Ishikawa 1998: The proposal of Komagatabacter gen. nov., for strains accommodated to the Gluconacetobacter xylinus group in the α-Proteobacteria. Ann. Microbiol. 2012, 62, 849–859. [Google Scholar] [CrossRef]
- Federal Agency on Technical Regulating and Metrology of the Russian Federation. Available online: https://protect.gost.ru/v.aspx?control=8&baseC=-1&page=0&month=-1&year=-1&search=&RegNum=1&DocOnPageCount=15&id=218419 (accessed on 24 February 2024).
- Menas, A.L.; Yanamala, N.; Farcas, M.T.; Russo, M.; Friend, S.; Fournier, P.M.; Star, A.; Iavicoli, I.; Shurin, G.V.; Vogel, U.B.; et al. Fibrillar vs. crystalline nanocellulose pulmonary epithelial cell responses: Cytotoxicity or inflammation? Chemosphere 2017, 171, 671–680. [Google Scholar] [CrossRef]
- Adewuyi, A.; Otuechere, C.A.; Adebayo, O.L.; Anazodo, C.; Pereira, F.V. Renal toxicological evaluations of sulphonated nanocellulose from Khaya sengalensis seed in Wistar rats. Chem. Biol. Interact. 2018, 284, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Kong, F. The behavior of nanocellulose in gastrointestinal tract and its influence on food digestion. J. Food Eng. 2021, 292, 110346. [Google Scholar] [CrossRef]
- Liang, X.; Fu, Y.; Cao, W.T.; Wang, Z.; Zhang, K.; Jiang, Z.; Jia, X.; Liu, C.; Lin, H.; Zhong, H.; et al. Gut microbiome, cognitive function and brain structure: A multi-omics integration analysis. Transl. Neurodegener. 2022, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Rescigno, M. The gut-brain vascular axis in neuroinflammation. Semin. Immunol. 2023, 69, 101802. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.P.; Pipek, L.Z.; Donaghey, T.C.; DeLoid, G.M.; Demokritou, P.; Brain, J.D.; Molina, R.M. Effects of ingested nanomaterials on tissue distribution of co-ingested zinc and iron in normal and zinc-deficient mice. NanoImpact 2021, 21, 100279. [Google Scholar] [CrossRef]
- Beketova, N.A.; Vrzhesinskaya, O.A.; Kosheleva, O.V.; Pereverzeva, O.G.; Isaeva, V.A.; Rudoy, B.A.; Dikovsky, A.V.; Kodentsova, V.M. Assessment of the ability of some dietary fibers to adsorb vitamins A, E, C, B1 and B2 in vitro. Vopr. Pitan. 2010, 2, 47–53. (In Russian) [Google Scholar]
- Stepan, M.D.; Vintilescu, Ș.B.; Streață, I.; Podeanu, M.A.; Florescu, D.N. The role of vitamin D in obese children with non-alcoholic fatty liver disease and associated metabolic syndrome. Nutrients 2023, 15, 2113. [Google Scholar] [CrossRef]
- Melguizo-Rodríguez, L.; Costela-Ruiz, V.J.; García-Recio, E.; De Luna-Bertos, E.; Ruiz, C.; Illescas-Montes, R. Role of vitamin D in the metabolic syndrome. Nutrients 2021, 13, 830. [Google Scholar] [CrossRef]
- Ujianti, I.; Sianipar, I.R.; Prijanti, A.R.; Hasan, I.; Arozal, W.; Jusuf, A.A.; Wibowo, H.; Prihartono, J.; Amani, P.; Santoso, D.I.S. Effect of Roselle Flower extract (Hibiscus sabdariffa Linn.) on reducing steatosis and steatohepatitis in vitamin B12 deficiency rat model. Medicina 2023, 59, 1044. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Aksenov, I.V.; Nikitin, N.S.; Guseva, G.V.; Avrenyeva, L.I.; Trusov, N.V.; Balakina, A.S.; Tutelyan, V.A. Lipoic acid exacerbates oxidative stress and lipid accumulation in the liver of Wistar rats fed a hypercaloric choline-deficient diet. Nutrients 2021, 13, 1999. [Google Scholar] [CrossRef]
- Riger, N.A.; Trushina, E.N.; Mustafina, O.K.; Timonin, A.N.; Aksenov, I.V.; Guseva, G.V.; Tutelyan, V.A. Pathogenetic mechanisms for the development of hematological disorders in induced fatty liver disease in Wistar rats and assessment of the regulatory effects of carnosine and α-lipoic acid. Vopr. Pitan. 2021, 3, 6–19. (In Russian) [Google Scholar] [CrossRef]
- Ede, J.D.; Ong, K.J.; Mulenos, M.R.; Pradhan, S.; Gibb, M.; Sayes, C.M.; Shatkin, J.A. Physical, chemical, and toxicological characterization of sulfated cellulose nanocrystals for food-related applications using in vivo and in vitro strategies. Toxicol. Res. 2020, 9, 808–822. [Google Scholar] [CrossRef]
- Ong, K.J.; Ede, J.D.; Pomeroy-Carter, C.A.; Sayes, C.M.; Mulenos, M.R.; Shatkin, J.A. A 90-day dietary study with fibrillated cellulose in Sprague-Dawley rats. Toxicol. Rep. 2020, 7, 174–182. [Google Scholar] [CrossRef]
- Shimazaki, T.; Deshpande, A.; Hajra, A.; Thomas, T.; Muta, K.; Yamada, N.; Yasui, Y.; Shoda, T. Deep learning-based image-analysis algorithm for classification and quantification of multiple histopathological lesions in rat liver. J. Toxicol. Pathol. 2022, 35, 135–147. [Google Scholar] [CrossRef]
- Morgan, A.E.; Mc Auley, M.T. Cholesterol homeostasis: An in silico investigation into how aging disrupts its key hepatic regulatory mechanisms. Biology 2020, 9, 314. [Google Scholar] [CrossRef]
- Oberdörster, G.; Oberdörster, E.; Oberdörster, J. Concepts of nanoparticle dose metric and response metric. Environ. Health Perspect. 2007, 115, A290. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G1 | G2 | G3 | |
|---|---|---|---|
| Reads received (number per file) | 2,817,576 | 2,100,066 | 776,416 |
| Both surviving | 2,084,829 (73.99%) | 1,447,380 (68.92%) | 567,571 (73.10%) |
| Forward-only surviving | 15,7254 (5.58%) | 118,147 (5.63%) | 48,302 (6.22%) |
| Reverse-only surviving | 423,014 (15.01%) | 399,165 (19.01%) | 114,818 (14.79%) |
| Dropped | 152,479 (5.41%) | 135,374 (6.45%) | 45,725 (5.89%) |
| Alphaproteobacteria | Betaproteobacteria | Gammaproteobacteria | |
|---|---|---|---|
| G1 | 14 | 21 | 65 |
| G2 | 7 | 49 | 42 |
| G3 | 3 | 81 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shipelin, V.A.; Skiba, E.A.; Budayeva, V.V.; Shumakova, A.A.; Kolobanov, A.I.; Sokolov, I.E.; Maisaya, K.Z.; Guseva, G.V.; Trusov, N.V.; Masyutin, A.G.; et al. Toxicological Characteristics of Bacterial Nanocellulose in an In Vivo Experiment—Part 1: The Systemic Effects. Nanomaterials 2024, 14, 768. https://0-doi-org.brum.beds.ac.uk/10.3390/nano14090768
Shipelin VA, Skiba EA, Budayeva VV, Shumakova AA, Kolobanov AI, Sokolov IE, Maisaya KZ, Guseva GV, Trusov NV, Masyutin AG, et al. Toxicological Characteristics of Bacterial Nanocellulose in an In Vivo Experiment—Part 1: The Systemic Effects. Nanomaterials. 2024; 14(9):768. https://0-doi-org.brum.beds.ac.uk/10.3390/nano14090768
Chicago/Turabian StyleShipelin, Vladimir A., Ekaterina A. Skiba, Vera V. Budayeva, Antonina A. Shumakova, Alexey I. Kolobanov, Ilya E. Sokolov, Kirill Z. Maisaya, Galina V. Guseva, Nikita V. Trusov, Alexander G. Masyutin, and et al. 2024. "Toxicological Characteristics of Bacterial Nanocellulose in an In Vivo Experiment—Part 1: The Systemic Effects" Nanomaterials 14, no. 9: 768. https://0-doi-org.brum.beds.ac.uk/10.3390/nano14090768