
In Vitro Gastrointestinal Digestion of Various Sweet Potato Leaves: Polyphenol Profiles, Bioaccessibility and Bioavailability Elucidation
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Chemical Composition Analysis
2.2.1. Ascorbic Acid Content (AAC) and Total Carbohydrate Content
2.2.2. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
2.3. Polyphenol Profile Analysis
2.4. Simulated Digestion
2.5. Bioactivity Evaluation
2.5.1. Antioxidant Capacity
2.5.2. Antihyperglycemic Activity
2.6. Statistical Analysis
3. Results
3.1. Chemical Composition Analysis
3.2. Phenolic Identification
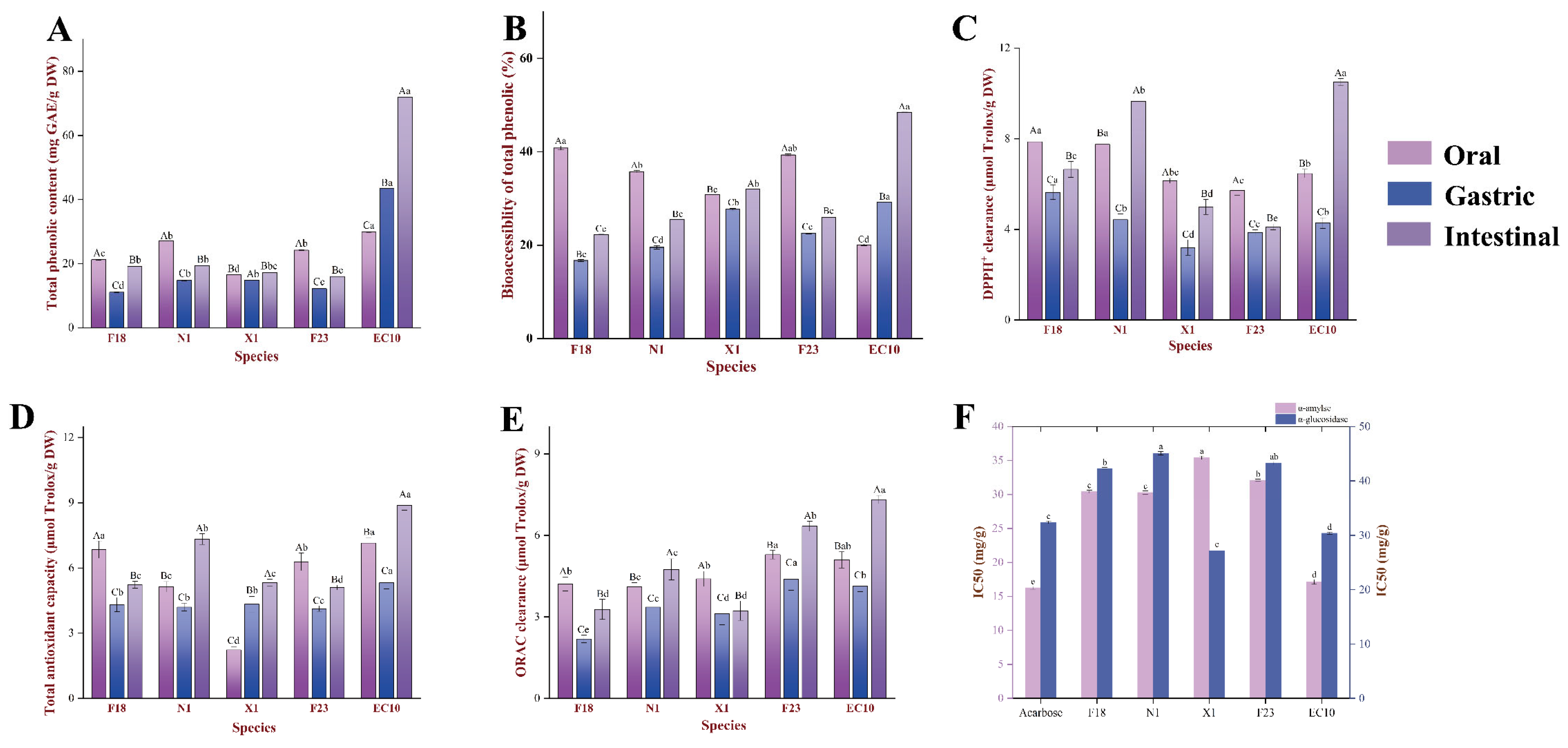
3.3. Bioaccessibility and Bioavailability Variations during Digestion In Vitro
3.3.1. Bioaccessibility of TPC
3.3.2. Bioaccessibility and Bioavailability of Phenolic Profile after Digestion
3.4. Antioxidant Activity Analysis
3.5. Anti-Glycemic Activity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hong, J.; Mu, T.; Sun, H.; Richel, A.; Blecker, C. Valorization of the green waste parts from sweet potato (Impoea batatas L.): Nutritional, phytochemical composition, and bioactivity evaluation. Food Sci. Nutr. 2020, 8, 4086–4097. [Google Scholar] [CrossRef]
- Tylewicz, U.; Oliveira, G.; Alminger, M.; Nohynek, L.; Dalla Rosa, M.; Romani, S. Antioxidant and antimicrobial properties of organic fruits subjected to PEF-assisted osmotic dehydration. Innov. Food Sci. Emerg. 2020, 62, 102341. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Chen, C.C.; Lin, K.H.; Chao, P.Y.; Lin, H.H.; Huang, M.Y. Bioactive Compounds, Antioxidants, and Health Benefits of Sweet Potato Leaves. Molecules 2021, 26, 1820. [Google Scholar] [CrossRef]
- Toy, J.Y.H.; Song, Z.; Huang, D. Resin glycosides in aerial parts of Ipomoea batatas are potent lipase inhibitors: Potential upcycling of sweet potato by-products to combat obesity. Food Funct. 2022, 13, 5353–5364. [Google Scholar] [CrossRef]
- Xi, L.; Mu, T.; Sun, H. Preparative purification of polyphenols from sweet potato (Ipomoea batatas L.) leaves by AB-8 macroporous resins. Food Chem. 2015, 172, 166–174. [Google Scholar] [CrossRef]
- Toy, J.Y.H.; See, J.W.; Huang, D. Physicochemical and functional characterisation of pectin from margarita sweet potato leaves. Food Chem. 2022, 385, 132684. [Google Scholar] [CrossRef]
- Sun, H.; Mu, B.; Song, Z.; Ma, Z.; Mu, T. The In Vitro Antioxidant Activity and Inhibition of Intracellular Reactive Oxygen Species of Sweet Potato Leaf Polyphenols. Oxid. Med. Cell. Longev. 2018, 2018, 9017828. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Paterson, J.; Bucknall, M.; Arcot, J. Interactions between phytochemicals from fruits and vegetables: Effects on bioactivities and bioavailability. Crit. Rev. Food Sci. Nutr. 2018, 58, 1310–1329. [Google Scholar] [CrossRef]
- Jagadeesan, G.; Muniyandi, K.; Manoharan, A.L.; Nataraj, G.; Thangaraj, P. Understanding the bioaccessibility, α-amylase and α-glucosidase enzyme inhibition kinetics of Allmania nodiflora (L.) R.Br. ex Wight polyphenols during in vitro simulated digestion. Food Chem. 2022, 372, 131294. [Google Scholar] [CrossRef]
- Murugan, R.; Chandran, R.; Parimelazhagan, T. Effect of in vitro simulated gastrointestinal digestion of Phoenix loureirii on polyphenolics, antioxidant and acetylcholinesterase inhibitory activities. LWT 2016, 74, 363–370. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of dietary flavonoids towards improved bioactivity: An update on structure-activity relationship. Crit. Rev. Food Sci. Nutr. 2018, 58, 513–527. [Google Scholar] [CrossRef]
- Izzo, L.; Rodríguez-Carrasco, Y.; Pacifico, S.; Castaldo, L.; Narváez, A.; Ritieni, A. Colon Bioaccessibility under In Vitro Gastrointestinal Digestion of a Red Cabbage Extract Chemically Profiled through UHPLC-Q-Orbitrap HRMS. Antioxidants 2020, 9, 955. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Yáñez, R.; Parajó, J.C.; Alonso, J.L. Pectic oligosacharides from lemon peel wastes: Production, purification, and chemical characterization. J. Agric. Food Chem. 2013, 61, 10043–10053. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G.; Yang, Z.; Hou, Y. Antioxidant activity of Momordica charantia polysaccharide and its derivatives. Int. J. Biol. Macromol. 2019, 138, 673–680. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of Total Phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar] [CrossRef]
- Sun, H.; Mu, T.; Xi, L.; Song, Z. Effects of domestic cooking methods on polyphenols and antioxidant activity of sweet potato leaves. J. Agric. Food Chem. 2014, 62, 8982–8989. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Huarte, E.; Serra, G.; Monteagudo-Mera, A.; Spencer, J.; Cid, C.; de Peña, M.P. Raw and Sous-Vide-Cooked Red Cardoon Stalks (Cynara cardunculus L. var. altilis DC): (Poly)phenol Bioaccessibility, Anti-inflammatory Activity in the Gastrointestinal Tract, and Prebiotic Activity. J. Agric. Food Chem. 2021, 69, 9270–9286. [Google Scholar] [CrossRef]
- Rufino, M.d.S.M.; Alves, R.E.; de Brito, E.S.; Pérez-Jiménez, J.; Saura-Calixto, F.; Mancini-Filho, J. Bioactive compounds and antioxidant capacities of 18 non-traditional tropical fruits from Brazil. Food Chem. 2010, 121, 996–1002. [Google Scholar] [CrossRef]
- Flores, F.P.; Singh, R.K.; Kerr, W.L.; Pegg, R.B.; Kong, F. Antioxidant and Enzyme Inhibitory Activities of Blueberry Antho-cyanins Prepared Using Different Solvents. J. Agric. Food. Chem. 2013, 61, 4441–4447. [Google Scholar] [CrossRef]
- Zeng, L.; Zhang, G.; Lin, S.; Gong, D. Inhibitory Mechanism of Apigenin on α-Glucosidase and Synergy Analysis of Flavonoids. J. Agric. Food. Chem. 2016, 64, 6939–6949. [Google Scholar] [CrossRef]
- Jeng, T.L.; Lai, C.C.; Liao, T.C.; Lin, S.Y.; Sung, J.M. Effects of drying on caffeoylquinic acid derivative content and antioxidant capacity of sweet potato leaves. J. Food Drug Anal. 2015, 23, 701–708. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Luthria, D.L. Drying affects artemisinin, dihydroartemisinic acid, artemisinic acid, and the antioxidant capacity of Artemisia annua L. leaves. J. Agric. Food Chem. 2010, 58, 1691–1698. [Google Scholar] [CrossRef]
- Mashitoa, F.M.; Shoko, T.; Shai, J.L.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Influence of Different Types of Drying Methods on Color Properties, Phenolic Metabolites and Bioactivities of Pumpkin Leaves of var. Butternut squash (Cucurbita moschata Duchesne ex Poir). Front. Nutr. 2021, 8, 694649. [Google Scholar] [CrossRef]
- Nallan Chakravartula, S.S.; Moscetti, R.; Farinon, B.; Vinciguerra, V.; Merendino, N.; Bedini, G.; Neri, L.; Pittia, P.; Massantini, R. Stinging Nettles as Potential Food Additive: Effect of Drying Processes on Quality Characteristics of Leaf Powders. Foods 2021, 10, 1152. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MSn. J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef]
- Gong, E.S.; Gao, N.; Li, T.; Chen, H.; Wang, Y.; Si, X.; Tian, J.; Shu, C.; Luo, S.; Zhang, J.; et al. Effect of In Vitro Digestion on Phytochemical Profiles and Cellular Antioxidant Activity of Whole Grains. J. Agric. Food Chem. 2019, 67, 7016–7024. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, B.; Xia, Y.; Li, H.; Shi, X.; Wang, J.; Deng, Z. Bioaccessibility and transformation pathways of phenolic compounds in processed mulberry (Morus alba L.) leaves after in vitro gastrointestinal digestion and faecal fermentation. J. Funct. Foods 2019, 60, 103406. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Fu, X.; Brennan, M.; Abbasi, A.M.; Zheng, B.; Liu, R.H. The use of an enzymatic extraction procedure for the enhancement of highland barley (Hordeum vulgare L.) phenolic and antioxidant compounds. Int. J. Food Sci. Technol. 2016, 51, 1916–1924. [Google Scholar] [CrossRef]
- Ferreira, D.M.; de Oliveira, N.M.; Chéu, M.H.; Meireles, D.; Lopes, L.; Oliveira, M.B.; Machado, J. Updated Organic Composition and Potential Therapeutic Properties of Different Varieties of Olive Leaves from Olea europaea. Plants 2023, 12, 688. [Google Scholar] [CrossRef]
- Baba, W.N.; McClements, D.J.; Maqsood, S. Whey protein-polyphenol conjugates and complexes: Production, characterization, and applications. Food Chem. 2021, 365, 130455. [Google Scholar] [CrossRef]
- Paull, D.H.; Weatherwax, A.; Lectka, T. Catalytic, asymmetric reactions of ketenes and ketene enolates. Tetrahedron 2009, 65, 3771–6803. [Google Scholar] [CrossRef]
- Lim, D.W.; Park, J.; Jung, J.; Kim, S.H.; Um, M.Y.; Yoon, M.; Kim, Y.T.; Han, D.; Lee, C.; Lee, J. Dicaffeoylquinic acids alleviate memory loss via reduction of oxidative stress in stress-hormone-induced depressive mice. Pharmacol. Res. 2020, 161, 105252. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, H.; Zeng, H.; Prinyawiwatukul, W.; Xu, W.; Xu, Z. Increases in Phenolic, Fatty Acid, and Phytosterol Contents and Anticancer Activities of Sweet Potato after Fermentation by Lactobacillus acidophilus. J. Agric. Food Chem. 2018, 66, 2735–2741. [Google Scholar] [CrossRef]
- Tong, J.; Ma, B.; Ge, L.; Mo, Q.; Zhou, G.; He, J.; Wang, Y. Dicaffeoylquinic Acid-Enriched Fraction of Cichorium glandulosum Seeds Attenuates Experimental Type 1 Diabetes via Multipathway Protection. J. Agric. Food Chem. 2015, 63, 10791–10802. [Google Scholar] [CrossRef]
- Xie, M.; Chen, G.; Wan, P.; Dai, Z.; Zeng, X.; Sun, Y. Effects of Dicaffeoylquinic Acids from Ilex kudingcha on Lipid Metabolism and Intestinal Microbiota in High-Fat-Diet-Fed Mice. J. Agric. Food Chem. 2019, 67, 171–183. [Google Scholar] [CrossRef]
- Gligor, O.; Clichici, S.; Moldovan, R.; Decea, N.; Vlase, A.M.; Fizeșan, I.; Pop, A.; Virag, P.; Filip, G.A.; Vlase, L.; et al. An In Vitro and In Vivo Assessment of Antitumor Activity of Extracts Derived from Three Well-Known Plant Species. Plants 2023, 12, 1840. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, Y.; Tan, Y.; Zhang, Z.; McClements, D.J. Digestibility and gastrointestinal fate of meat versus plant-based meat analogs: An in vitro comparison. Food Chem 2021, 364, 130439. [Google Scholar] [CrossRef]
- Managa, M.G.; Akinola, S.A.; Remize, F.; Garcia, C.; Sivakumar, D. Physicochemical Parameters and Bioaccessibility of Lactic Acid Bacteria Fermented Chayote Leaf (Sechium edule) and Pineapple (Ananas comosus) Smoothies. Front. Nutr. 2021, 8, 649189. [Google Scholar] [CrossRef]
- Oszmiański, J.; Lachowicz, S.; Nowicka, P.; Rubiński, P.; Cebulak, T. Evaluation of Innovative Dried Purée from Jerusalem Artichoke—In Vitro Studies of Its Physicochemical and Health-Promoting Properties. Molecules 2021, 26, 2644. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Bresciani, L.; Dall’Asta, M.; Mena, P.; Del Rio, D.; Cid, C.; de Peña, M.-P. Bioaccessibility of (poly)phenolic compounds of raw and cooked cardoon (Cynara cardunculus L.) after simulated gastrointestinal digestion and fermentation by human colonic microbiota. J. Funct. 2017, 32, 195–207. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-d’Arce, M.A.; Biasoto, A.C.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef]
- Jakobek, L.; Strelec, I.; Kenjerić, D.; Šoher, L.; Tomac, I.; Matić, P. Simulated Gastric and Intestinal Fluid Electrolyte Solutions as an Environment for the Adsorption of Apple Polyphenols onto β-Glucan. Molecules 2022, 27, 6683. [Google Scholar] [CrossRef]
- Laurent, C.; Besançon, P.; Caporiccio, B. Flavonoids from a grape seed extract interact with digestive secretions and intestinal cells as assessed in an in vitro digestion/Caco-2 cell culture model. Food Chem. 2007, 100, 1704–1712. [Google Scholar] [CrossRef]
- Xue, M.; Shi, H.; Zhang, J.; Liu, Q.Q.; Guan, J.; Zhang, J.Y.; Ma, Q. Stability and Degradation of Caffeoylquinic Acids under Different Storage Conditions Studied by High-Performance Liquid Chromatography with Photo Diode Array Detection and High-Performance Liquid Chromatography with Electrospray Ionization Collision-Induced Dissociation Tandem Mass Spec-trometry. Molecules 2016, 21, 948. [Google Scholar] [CrossRef]
- Wang, D.; Wang, Y.; Zhang, Z.; Qiu, S.; Yuan, Y.; Song, G.; Li, L.; Yuan, T.; Gong, J. Degradation, isomerization and stabilization of three dicaffeoylquinic acids under ultrasonic treatment at different pH. Ultrason. Sonochem. 2023, 95, 106401. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, X.; Liu, P.; Li, M.; Dong, H.; Qiao, X. Semi-Preparative Separation of 10 Caffeoylquinic Acid Derivatives Using High Speed Counter-Current Chromatogaphy Combined with Semi-Preparative HPLC from the Roots of Burdock (Arctium lappa L.). Molecules 2018, 23, 429. [Google Scholar] [CrossRef]
- Wu, L.; Liu, Y.; Qin, Y.; Wang, L.; Wu, Z. HPLC-ESI-qTOF-MS/MS Characterization, Antioxidant Activities and Inhibitory Ability of Digestive Enzymes with Molecular Docking Analysis of Various Parts of Raspberry (Rubus ideaus L.). Antioxidants 2019, 8, 274. [Google Scholar] [CrossRef]
- Guo, R.; Chang, X.; Guo, X.; Brennan, C.S.; Li, T.; Fu, X.; Liu, R.H. Phenolic compounds, antioxidant activity, antiproliferative activity and bioaccessibility of Sea buckthorn (Hippophaë rhamnoides L.) berries as affected by in vitro digestion. Food Funct. 2017, 8, 4229–4240. [Google Scholar] [CrossRef]
- Kumar, Y.; Tarafdar, A.; Kumar, D.; Saravanan, C.; Badgujar, P.C.; Pharande, A.; Pareek, S.; Fawole, O.A. Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient. Antioxidants 2022, 11, 993. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, Y.; Gao, J.; Feng, J.; Shang, Y.; Wei, Z. Phenolics and antioxidant activity of bamboo leaves soup as affected by in vitro digestion. Food Chem. Toxicol. 2020, 135, 110941. [Google Scholar] [CrossRef]
- Wang, L.; Luo, Y.; Wu, Y.; Xia, F.; Wu, Z. Quickly verifying the antioxidant contribution of the individual composition in natural antioxidants by HPLC-free radical scavenging detection. LWT 2018, 96, 461–468. [Google Scholar] [CrossRef]
- Cuvas-Limon, R.B.; Ferreira-Santos, P.; Cruz, M.; Teixeira, J.A.; Belmares, R.; Nobre, C. Effect of Gastrointestinal Digestion on the Bioaccessibility of Phenolic Compounds and Antioxidant Activity of Fermented Aloe vera Juices. Antioxidants 2022, 11, 2479. [Google Scholar] [CrossRef]
- Lachowicz, S.; Wiśniewski, R.; Ochmian, I.; Drzymała, K.; Pluta, S. Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry. Antioxidants 2019, 8, 397. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, L.; Liu, Y.; Zhang, Q.; Li, Y.; Wu, Z. Release of phenolics compounds from Rubus idaeus L. dried fruits and seeds during simulated in vitro digestion and their bio-activities. J. Funct. 2018, 46, 57–65. [Google Scholar] [CrossRef]
- Silva, C.P.D.; Soares-Freitas, R.A.M.; Sampaio, G.R.; Santos, M.C.B.; do Nascimento, T.P.; Cameron, L.C.; Ferreira, M.S.L.; Arêas, J.A.G. Identification and action of phenolic compounds of Jatobá-do-cerrado (Hymenaea stignocarpa Mart.) on α-amylase and α-glucosidase activities and flour effect on glycemic response and nutritional quality of breads. Food Res. Int. 2019, 116, 1076–1083. [Google Scholar] [CrossRef]
- Sun, L.; Warren, F.J.; Gidley, M.J.; Guo, Y.; Miao, M. Mechanism of binding interactions between young apple polyphenols and porcine pancreatic α-amylase. Food Chem. 2019, 283, 468–474. [Google Scholar] [CrossRef]
- Fu, Z.F.; Tu, Z.C.; Zhang, L.; Wang, H.; Wen, Q.H.; Huang, T. Antioxidant activities and polyphenols of sweet potato (Ipomoea batatas L.) leaves extracted with solvents of various polarities. Food Biosci. 2016, 15, 11–18. [Google Scholar] [CrossRef]
- Lu, X.F.; Zhou, Y.; Ren, Y.P.; Zhang, J. Improved sample treatment for the determination of flavonoids and polyphenols in sweet potato leaves by ultra performance convergence chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2019, 169, 245–253. [Google Scholar] [CrossRef]
- Sun, R.; Kan, J.; Cai, H.; Hong, J.; Jin, C.; Zhang, M. In vitro and in vivo ameliorative effects of polyphenols from purple potato leaves on renal injury and associated inflammation induced by hyperuricemia. J. Food Biochem. 2022, 46, e14049. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.C.; Wang, H.; Fu, Z.F.; Wen, Q.H.; Chang, H.X.; Huang, X.Q. Comparison of different methods for extracting polyphenols from Ipomoea batatas leaves, and identification of antioxidant constituents by HPLC-QTOF-MS2. Food Res. Int. 2015, 70, 101–109. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CQAs | Phenolic | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3-CQA | 4-CQA | 5-CQA | 3,4-CQA | 3,5-CQA | 4,5-CQA | 3,4,5-CQA | CA | Quinine Acid | ||
| Oral (Bioaccessibility %) | F18 | 0.58 b | nd | nd | 0.72 c | 0.34 c | 1.34 b | nd | 15.41 b | 100.00 a |
| N1 | 0.41 c | nd | 1.90 c | 0.97 b | 0.34 c | 0.74 c | 0.62c | 274.16 a | nd | |
| X1 | 0.51 b | nd | 2.52 b | nd | 2.70 a | 2.54 b | nd | 4.81 d | nd | |
| F23 | 0.86 a | 3.54 a | 3.99 a | 1.22 a | 0.39 b | 4.08 a | 7.45 a | 3.25 e | nd | |
| EC10 | 0.55 c | nd | 2.48 bc | nd | 0.32 c | 2.73 b | 4.38 b | 12.17 c | nd | |
| Gastric (Bioaccessibility %) | F18 | 0.27 d | nd | 3.75 b | nd | 0.14 d | 1.48 d | 3.89 c | 49.01 b | 100.00 a |
| N1 | 0.81 b | 3.45 a | 3.36 b | 1.09 a | 0.29 c | 2.49 b | 6.54 b | 1.25 e | nd | |
| X1 | nd | nd | nd | nd | 0.45 a | 3.99 a | 9.01 a | 23.19 c | nd | |
| F23 | nd | nd | nd | 0.99 a | 0.43 a | 1.81 c | nd | 16.15 d | nd | |
| EC10 | 0.97 a | nd | 6.57 a | nd | 0.34 b | 3.84 a | 6.83 b | 69.01 a | 100.00 a | |
| Intestinal (Bioaccessibility %) | F18 | 0.97 c | nd | 3.70 b | 2.20 b | 0.25 c | 3.22 c | 6.88 a | 4.16 c | nd |
| N1 | 0.67 d | 1.32 c | 1.81 d | 1.30 c | 0.47 a | 3.79 b | 6.88 a | 12.90 b | nd | |
| X1 | 0.52 e | 3.40 b | 2.96 c | 1.20 d | 0.36 b | 3.86 b | nd | nd | nd | |
| F23 | 1.34 b | 3.54 b | 3.26 bc | 1.08 d | 0.47 a | 1.76 d | nd | nd | nd | |
| EC10 | 9.75 a | 57.39 a | 79.37 a | 6.55 a | 0.27 c | 13.18 a | 5.12 b | 383.68 a | nd | |
| Bioavailability (%) | F18 | 45.34 b | nd | 61.08 b | 39.54 d | nd | 36.62 c | 47.25 b | 78.36 a | nd |
| N1 | 57.67 a | 62.16 b | 80.74 a | 40.78 d | 34.07 c | 78.88 b | 60.12 ab | 73.20 b | nd | |
| X1 | 55.85 ab | 66.23 b | 83.11 a | 68.31 b | 41.68 b | 69.43 c | nd | nd | nd | |
| F23 | 46.47 b | 75.32 a | 86.24 a | 60.90 c | 40.39 b | 73.31 bc | nd | nd | nd | |
| EC10 | 60.01 a | 68.86 b | 62.12 b | 79.13 a | 62.98 a | 89.47 a | 67.36 a | 76.35 a | nd | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.; Sui, Y.; Shi, J.; Cai, S.; Xiong, T.; Cai, F.; Zhou, L.; Li, S.; Mei, X. In Vitro Gastrointestinal Digestion of Various Sweet Potato Leaves: Polyphenol Profiles, Bioaccessibility and Bioavailability Elucidation. Antioxidants 2024, 13, 520. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050520
Wen J, Sui Y, Shi J, Cai S, Xiong T, Cai F, Zhou L, Li S, Mei X. In Vitro Gastrointestinal Digestion of Various Sweet Potato Leaves: Polyphenol Profiles, Bioaccessibility and Bioavailability Elucidation. Antioxidants. 2024; 13(5):520. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050520
Chicago/Turabian StyleWen, Junren, Yong Sui, Jianbin Shi, Sha Cai, Tian Xiong, Fang Cai, Lei Zhou, Shuyi Li, and Xin Mei. 2024. "In Vitro Gastrointestinal Digestion of Various Sweet Potato Leaves: Polyphenol Profiles, Bioaccessibility and Bioavailability Elucidation" Antioxidants 13, no. 5: 520. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050520