

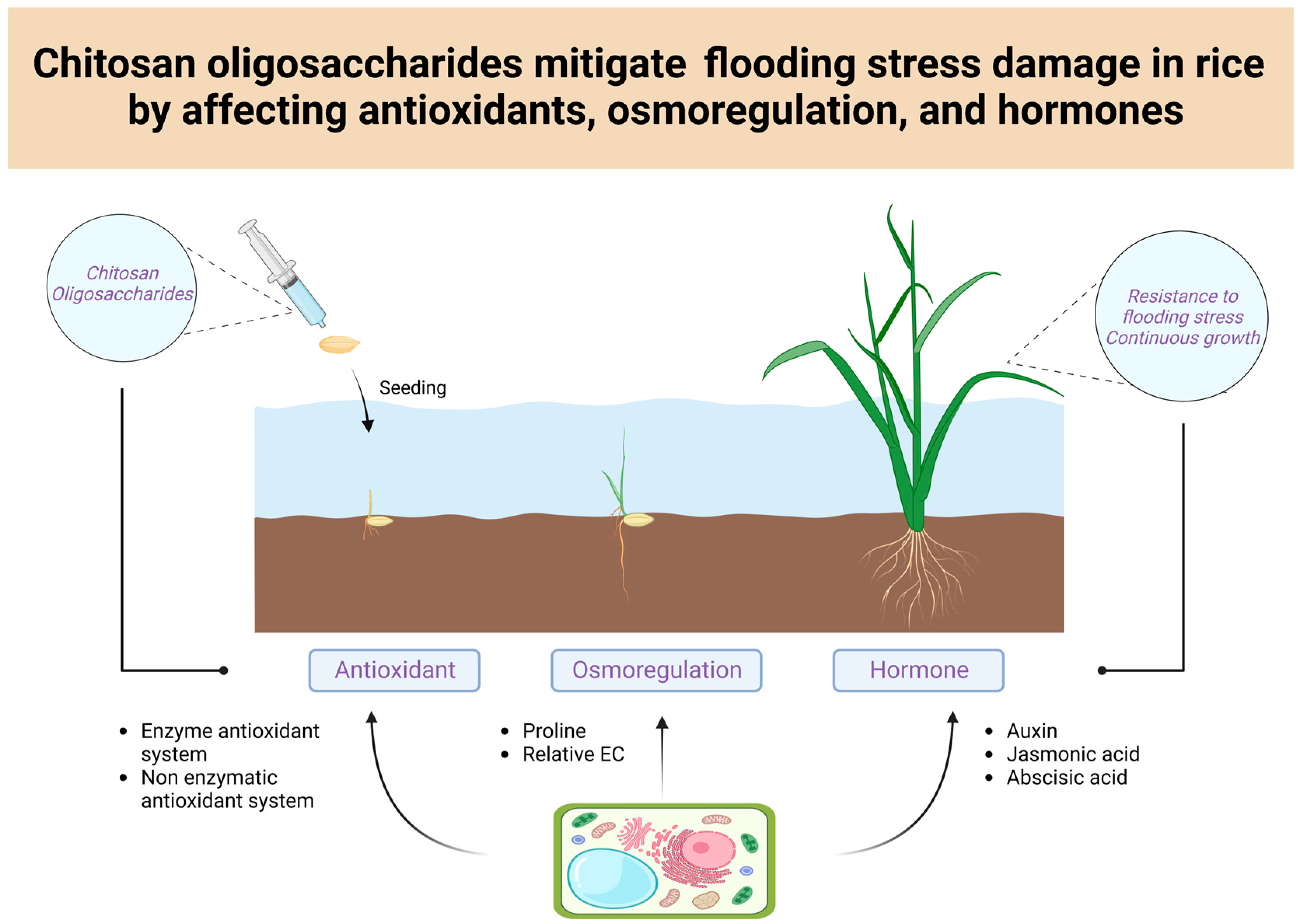
Chitosan Oligosaccharides Mitigate Flooding Stress Damage in Rice by Affecting Antioxidants, Osmoregulation, and Hormones
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Treatments
2.2. Biochemical Assays
2.3. RNA Extraction, Library Construction, RNA Sequencing, and Data Analysis
2.4. Metabolite Profiling and Data Analysis
2.5. Field Trial Verification
2.6. Data Analysis
3. Results
3.1. Seed Soaking with Chitosan Oligosaccharides Can Alleviate the Inhibitory Effect of Flooding Stress
3.2. Physiological Characteristics of Seed Soaking with Chitosan Oligosaccharides to Improve the Flooding Tolerance of Rice
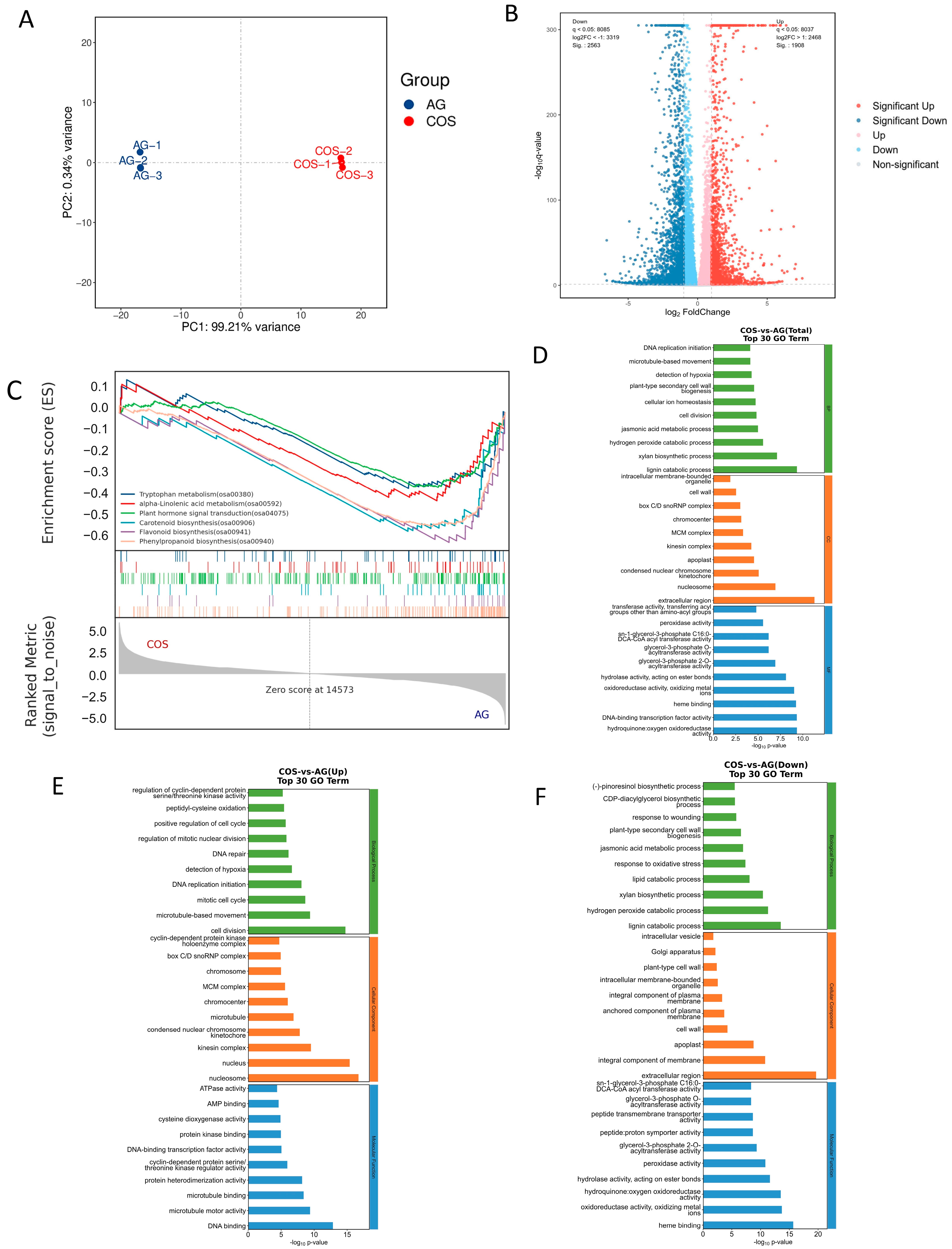
3.3. RNA-Seq Analysis of Effect of Seed Soaking with Chitosan Oligosaccharides on Improving Rice’s Submergence Tolerance
3.4. Metabolomic Differences of Seed Soaking with Chitosan Oligosaccharides in Improving the Flooding Tolerance of Rice
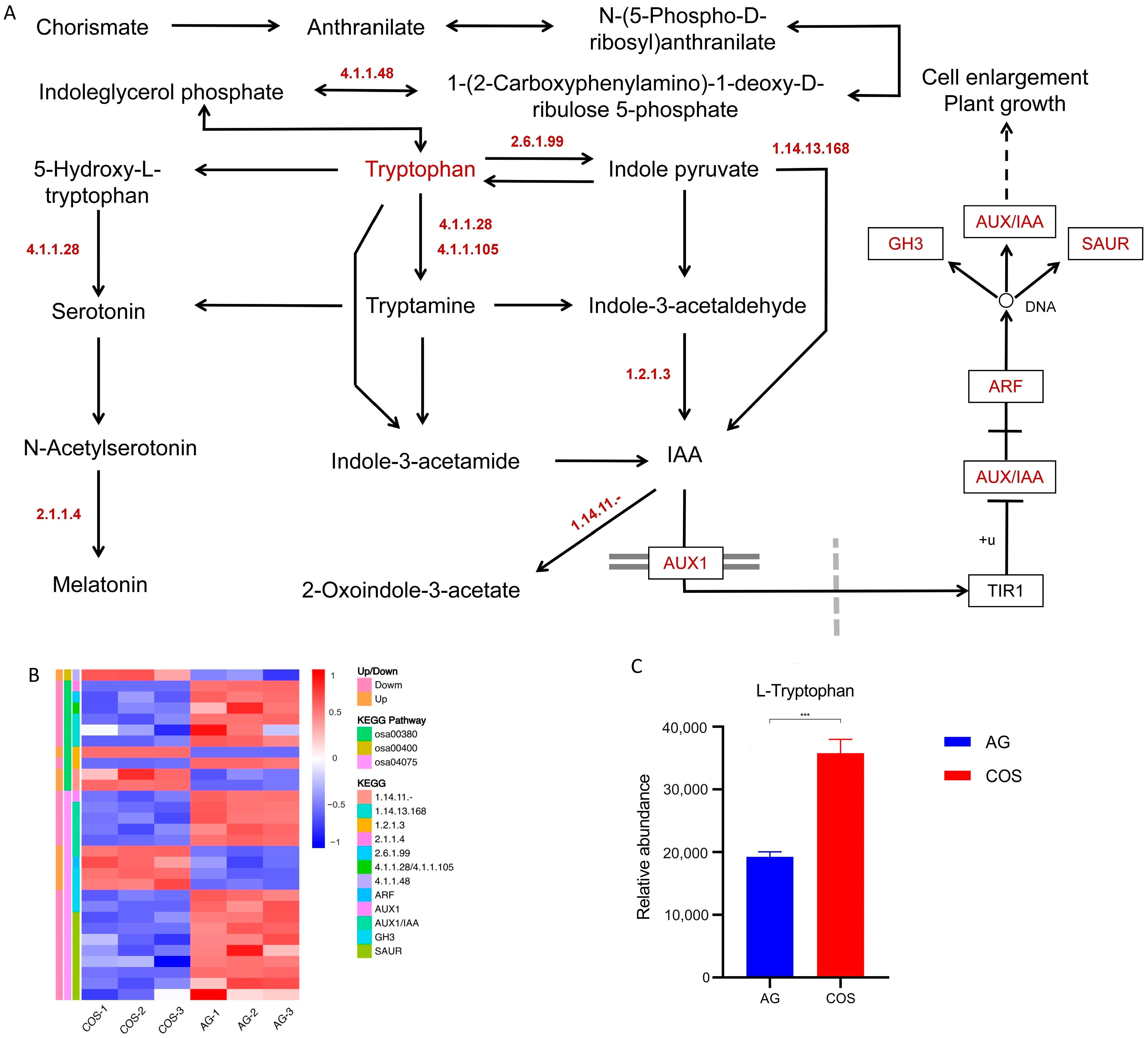
3.5. Combined Analysis of Transcriptome and Metabolome of Chitosan Oligosaccharides Seed Soaking’s Improvement Effect on the Flooding Tolerance of Rice Seedlings
3.5.1. Tryptophan Synthesis and Metabolism–Auxin Signaling Pathway
3.5.2. Jasmonic Acid Biosynthesis and Signal Transduction
3.5.3. β-Carotene Biosynthesis–Abscisic Acid Biosynthesis and Signal Transduction
3.6. Field Effect Verification of the Effect of Seed Soaking with Chitosan Oligosaccharides on Improving the Submergence Tolerance of Rice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef]
- Farooq, M.K.; Siddique, K.H.M.; Rehman, H.; Aziz, T.; Lee, D.-J.; Wahid, A. Rice direct seeding: Experiences, challenges and opportunities. Soil. Tillage Res. 2011, 111, 87–98. [Google Scholar] [CrossRef]
- Yang, Z.; Yu, Z.; Zhang, X.; Liao, Q.; Fu, H.; Cheng, Q.; Chen, Z.; Sun, Y.; Ma, J.; Zhang, J.; et al. Unmanned aerial vehicle direct seeding or integrated mechanical transplanting, which will be the next step for mechanized rice production in China?—A comparison based on energy use efficiency and economic benefits. Energy 2023, 273, 127223. [Google Scholar] [CrossRef]
- Turra, G.M.; Cutti, L.; Angonese, P.S.; Sulzbach, E.; Mariot, C.H.P.; Markus, C.; Merotto Junior, A. Variability to flooding tolerance in barnyardgrass and early flooding benefits on weed management and rice grain yield. Field Crops Res. 2023, 300, 108999. [Google Scholar] [CrossRef]
- Lu, H.; Wang, M.; Li, W.; Chen, Z.; Li, S.; Yi, Z.; Zhang, Y. Superior Antioxidant Capacity and Auxin Production Promote Seedling Formation of Rice Seeds under Submergence Stress. Agronomy 2023, 13, 171. [Google Scholar] [CrossRef]
- Ye, N.H.; Wang, F.Z.; Shi, L.; Chen, M.X.; Cao, Y.Y.; Zhu, F.Y.; Wu, Y.Z.; Xie, L.J.; Liu, T.Y.; Su, Z.Z.; et al. Natural variation in the promoter of rice calcineurin B-like protein10 (OsCBL10) affects flooding tolerance during seed germination among rice subspecies. Plant J. 2018, 94, 612–625. [Google Scholar] [CrossRef]
- Galvin, L.B.; İnci, D.; Mesgaran, M.B.; Brim-DeForest, W.; Al-Khatib, K. Flooding depths and burial effects on seedling emergence of five California weedy rice (Oryza sativa spontanea) accessions. Weed Sci. 2022, 70, 213–219. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.-J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef]
- He, Y.; Sun, S.; Zhao, J.; Huang, Z.; Peng, L.; Huang, C.-W.; Tang, Z.; Huang, Q.; Wang, Z. UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination. Nat. Commun. 2023, 14, 2296. [Google Scholar] [CrossRef]
- Rawyler, A. Impact of Oxygen Stress and Energy Availability on Membrane Stability of Plant Cells. Ann. Bot. 2002, 90, 499–507. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Song, X.; Furukawa, S.; Sakakibara, H.; Mori, H.; Matsuoka, M.; Ashikari, M. Analyses for Snorkel1 and Snorkel2, the responsible genes of deepwater rice character. Plant Cell Physiol. Suppl. 2009, 2009, 0352. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, A.K.; Hanjagi, P.S.; Kumari, K.; Vijayakumar, S.; Mohanty, S.; Tripathi, R.; Panneerselvam, P. Submergence stress in rice: Adaptive mechanisms, coping strategies and future research needs. Environ. Exp. Bot. 2021, 186, 104448. [Google Scholar] [CrossRef]
- Miro, B.; Ismail, A.M. Tolerance of anaerobic conditions caused by flooding during germination and early growth in rice (Oryza sativa L.). Front. Plant Sci. 2013, 4, 269. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.E.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Miftahul, A.; Siwi, I.; Christanti, S. RICE SEED TREATMENTS BY SOAKING IN WATER AND THREE PESTICIDES (CARBOSULFAN, BENOMYL, AND IMIDACLOPRID) TO CONTROL FOLIAR NEMATODE Aphelenchoides besseyi. J. Agroteknologi 2023, 14, 19–24. [Google Scholar] [CrossRef]
- Gai, D.; Liu, W.; Liang, J.; Liu, Y.; Geng, Y.; Zhang, Q.; Du, J.; Gao, J.; Shao, X. The Effects of Paclobutrazol Seed Soaking on Biomass Production and Yield Formation in Direct-Seeded Rice. Agronomy 2023, 13, 1402. [Google Scholar] [CrossRef]
- Lizansari, K.N.; Machmud, M.; Ilyas, S. Seed treatments and seedling root soaking in biological agents improved plant growth and reduced bacterial leaf blight disease incidence in rice. IOP Conf. Ser. 2023, 1133, 012031. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Nafees, M.; Chen, J.; Darras, A.I.; Ferrante, A.; Hancock, J.T.; Ashraf, M.; Zaid, A.; Latif, N.; Corpas, F.J.; et al. Chemical priming enhances plant tolerance to salt stress. Front. Plant Sci. 2022, 13, 946922. [Google Scholar] [CrossRef]
- Khaleduzzaman, M.; Hossain, M.A.; Bhuiyan, M.J.H.; Mahmud, S.; Tahjib-UI-Arif, M.; Murata, Y. Chitosan mitigates salt stress in rice by enhancing antioxidant defense system. Fundam. Appl. Agric. 2021, 6, 336–348. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif; Brestic, M.; Afrin, S.; Sakil, M.A.; Hossain, M.T.; Hossain, M.A.; Hossain, M.A. Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil. Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef]
- Violita, V.; Azhari, S.N. Effect of PEG-8000 imposed drought stress on rice varieties germination. J. Phys. 2021, 1940, 012071. [Google Scholar] [CrossRef]
- Xie, Z.; Wang, J.; Wang, W.; Wang, Y.; Xu, J.; Li, Z.; Zhao, X.; Fu, B. Integrated Analysis of the Transcriptome and Metabolome Revealed the Molecular Mechanisms Underlying the Enhanced Salt Tolerance of Rice Due to the Application of Exogenous Melatonin. Front. Plant Sci. 2021, 11, 618680. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Qu, D.; Chen, X.; Zeng, H.; Li, X.; Hu, C.Y. Metabolomics Reveals 5-Aminolevulinic Acid Improved the Ability of Tea Leaves (Camellia sinensis L.) against Cold Stress. Metabolites 2022, 12, 392. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, H.; Wen, F.; Bao, L.; Zhao, Z.; Zhong, Z. Chitooligosaccharide-induced plant stress resistance. Carbohydr. Polym. 2023, 302, 120344. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shu, R.; Shao, J.; Xu, G.; Gu, X. Radical scavenging activity of chitooligosaccharide with different molecular weights. Eur. Food Res. Technol. 2006, 222, 36–40. [Google Scholar] [CrossRef]
- El Gueddari, N.E.; Rauchhaus, U.; Moerschbacher, B.M.; Deising, H.B. Developmentally regulated conversion of surface-exposed chitin to chitosan in cell walls of plant pathogenic fungi. New Phytol. 2002, 156, 103–112. [Google Scholar] [CrossRef]
- Kim, S.W.; Park, J.K.; Lee, C.H.; Hahn, B.-S.; Koo, J.C. Comparison of the Antimicrobial Properties of Chitosan Oligosaccharides (COS) and EDTA against Fusarium fujikuroi Causing Rice Bakanae Disease. Curr. Microbiol. 2016, 72, 496–502. [Google Scholar] [CrossRef]
- Tang, W.; Lei, X.; Liu, X.; Yang, F. Nutritional Improvement of Bean Sprouts by Using Chitooligosaccharide as an Elicitor in Germination of Soybean (Glycine max L.). Appl. Sci. 2021, 11, 7695. [Google Scholar] [CrossRef]
- Benchamas, G.; Huang, G.; Huang, S.; Huang, H. Preparation and biological activities of chitosan oligosaccharides. Trends Food Sci. Technol. 2021, 107, 38–44. [Google Scholar] [CrossRef]
- Eshpulatov, N.; Khalmuradov, T.; Khalilov, R.; Obidov, A.; Allanazarov, M. Electrical conductivity of whole and shredded plant tissue. E3S Web Conf. 2021, 264, 04072. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, L.; Ma, J.; Fu, Q.; Zheng, Z.; Du, E.; Xu, Y.; Zhang, Z. Biotransformation of graphene oxide within lung fluids could intensify its synergistic biotoxicity effect with cadmium by inhibiting cellular efflux of cadmium. Environ. Pollut. 2022, 306, 119421. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zong, Y.; Li, W.; Guo, G.; Zhou, L.; Xu, H.; Gao, R.; Liu, C. Transcriptomics integrated with metabolomics reveals the effect of cold stress on rice microspores. BMC Plant Biol. 2023, 23, 521. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Du, F.; Wang, J.; Li, Y.; Zhang, Y.; Zhao, X.; Zheng, T.; Li, Z.; Xu, J.; Wang, W.; et al. Molecular Dissection of the Gene OsGA2ox8 Conferring Osmotic Stress Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 9107. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Striker, G.G. Plant Responses to Hypoxia: Signaling and Adaptation. Plants 2020, 9, 1704. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, G.; Xu, H.; Jing, S.; Jiang, Y.; Liu, Z.; Zhang, H.; Wang, F.; Hu, X.; Zhu, Y. Transcriptome Analysis of Rice Embryo and Endosperm during Seed Germination. Int. J. Mol. Sci. 2023, 24, 8710. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Park, P.-J.; Kim, S.-K. Free radical scavenging properties of hetero-chitooligosaccharides using an ESR spectroscopy. Food Chem. Toxicol. 2004, 42, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Kittur, F.S.; Vishu Kumar, A.B.; Varadaraj, M.C.; Tharanathan, R.N. Chitooligosaccharides—Preparation with the aid of pectinase isozyme from Aspergillus niger and their antibacterial activity. Carbohydr. Res. 2005, 340, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Muanprasat, C.; Chatsudthipong, V. Chitosan oligosaccharide: Biological activities and potential therapeutic applications. Pharmacol. Ther. 2017, 170, 80–97. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, S.; Malde, A.K.; Tan, X.; Gilbert, R.G. Effects of chickpea protein fractions on α-amylase activity in digestion. Food Hydrocoll. 2022, 133, 108005. [Google Scholar] [CrossRef]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedlings to Tolerate Flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Mora-Poblete, F.; Arnao, M.B.; Naz, S.; Anwar, M.; Altaf, M.M.; Shahid, S.; Shakoor, A.; et al. Phytomelatonin: An overview of the importance and mediating functions of melatonin against environmental stresses. Physiol. Plant. 2021, 172, 820–846. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. Auxin biosynthesis: A simple two-step pathway converts tryptophan to indole-3-acetic acid in plants. Mol. Plant 2012, 5, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Liu, M.; Wang, X.; Liu, L.; Wu, H.; Chen, X.; Wang, H.; Shen, Q.; Chen, G.; Wang, Y. Seed-Soaking with Melatonin for the Improvement of Seed Germination, Seedling Growth, and the Antioxidant Defense System under Flooding Stress. Agronomy 2022, 12, 1918. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, Y.; Sun, Y.; Xie, Z.; Luo, Y.; Long, Q.; Feng, J.; Liu, X.; Wang, B.; He, D.; et al. Natural variations of OsAUX5, a target gene of OsWRKY78, control the neutral essential amino acid content in rice grains. Mol. Plant 2023, 16, 322–336. [Google Scholar] [CrossRef]
- Chen, S.-H.; Zhou, L.-J.; Xu, P.; Xue, H.-W. SPOC domain-containing protein Leaf inclination3 interacts with LIP1 to regulate rice leaf inclination through auxin signaling. PLoS Genet. 2018, 14, e1007829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, R.; Xing, J.; Yan, L.; Wang, R.; Zhao, Y. The YUCCA-Auxin-WOX11 Module Controls Crown Root Development in Rice. Front. Plant Sci. 2018, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, Z.; Li, X.; Lv, T.; Liu, H.; Wang, L.; Niu, H.; Bu, Q. Characterization and Functional Analysis of Pyrabactin Resistance-Like Abscisic Acid Receptor Family in Rice. Rice 2015, 8, 28. [Google Scholar] [CrossRef]
- Chehab, E.W.; Raman, G.; Walley, J.W.; Perea, J.V.; Banu, G.; Theg, S.; Dehesh, K. Rice HYDROPEROXIDE LYASES with Unique Expression Patterns Generate Distinct Aldehyde Signatures in Arabidopsis. Plant Physiol. 2006, 141, 121–134. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Yang, Z.; Wang, C.; Li, S.; Cao, C.; Yao, T.; Wei, Z.; Li, Y.; Chen, J.; et al. Independently evolved viral effectors convergently suppress DELLA protein SLR1-mediated broad-spectrum antiviral immunity in rice. Nat. Commun. 2022, 13, 6920. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, Y.; Charnikhova, T.; Mulder, P.P.J.; Heijmans, J.; Hoogenboom, A.; Agalou, A.; Michel, C.; Morel, J.-B.; Dreni, L.; et al. OsJAR1 is required for JA-regulated floret opening and anther dehiscence in rice. Plant Mol. Biol. 2014, 86, 19–33. [Google Scholar] [CrossRef]
- Min, M.K.; Kim, R.; Hong, W.-J.; Jung, K.-H.; Lee, J.-Y.; Kim, B.-G. OsPP2C09 Is a Bifunctional Regulator in Both ABA-Dependent and Independent Abiotic Stress Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 393. [Google Scholar] [CrossRef] [PubMed]
- Balusamy, S.R.; Rahimi, S.; Sukweenadhi, J.; Sunderraj, S.; Shanmugam, R.; Thangavelu, L.; Mijakovic, I.; Perumalsamy, H. Chitosan, chitosan nanoparticles and modified chitosan biomaterials, a potential tool to combat salinity stress in plants. Carbohydr. Polym. 2022, 284, 119189. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Category | Term | ES | NES | p-Value | FDR |
|---|---|---|---|---|---|
| GO-BP | glycolytic process (GO:0006096) | 0.50 | 2.06 | 0.00 | 0.00 |
| gluconeogenesis (GO:0006094) | 0.64 | 1.99 | 0.00 | 0.00 | |
| response to hypoxia (GO:0001666) | 0.58 | 1.91 | 0.00 | 0.00 | |
| response to oxidative stress (GO:0006979) | −0.48 | −2.18 | 0.00 | 0.00 | |
| jasmonic acid metabolic process (GO:0009694) | −0.72 | −2.24 | 0.00 | 0.00 | |
| response to abscisic acid (GO:0009737) | −0.30 | −1.39 | 0.00 | 0.17 | |
| response to reactive oxygen species (GO:0000302) | 0.42 | 1.55 | 0.02 | 0.08 | |
| auxin polar transport (GO:0009926) | 0.38 | 1.46 | 0.02 | 0.12 | |
| carotenoid biosynthetic process (GO:0016117) | −0.47 | −1.47 | 0.04 | 0.13 | |
| GO-MF | oxidoreductase activity (GO:0016491) | −0.38 | −1.72 | 0.00 | 0.02 |
| methyl indole-3-acetate esterase activity (GO:0080030) | −0.71 | −1.98 | 0.00 | 0.00 | |
| peroxidase activity (GO:0004601) | −0.55 | −2.39 | 0.00 | 0.00 | |
| oxidoreductase activity, oxidizing metal ions (GO:0016722) | −0.73 | −2.44 | 0.00 | 0.00 | |
| UDP-glucosyltransferase activity (GO:0035251) | −0.52 | −1.76 | 0.00 | 0.01 | |
| UDP-glycosyltransferase activity (GO:0008194) | −0.39 | −1.53 | 0.02 | 0.06 |
| Variety | Treatment | Seedling Height (cm) | Root Length (cm) | Number of Roots | Stem Base Width (mm) | Fresh Weight (g) | Basic Number of Seedlings |
|---|---|---|---|---|---|---|---|
| YLY948 | AG | 4.70 ± 0.25 b | 2.95 ± 0.49 b | 3.20 ± 0.22 b | 16.40 ± 0.11 b | 0.62 ± 0.01 b | 22.33 ± 0.48 c |
| COS | 5.12 ± 0.13 a | 4.24 ± 0.31 a | 3.83 ± 0.35 a | 16.27 ± 0.18 b | 0.66 ± 0.01 a | 26.79 ± 0.43 b | |
| CK | 4.07 ± 0.07 c | 4.83 ± 0.40 a | 3.93 ± 0.43 a | 16.72 ± 0.07 a | 0.65 ± 0.01 a | 31.65 ± 1.51 a | |
| NX42 | AG | 3.24 ± 0.16 b | 2.27 ± 0.19 c | 2.0 ± 0.29 b | 14.71 ± 0.45 b | 0.61 ± 0.02 a | 15.92 ± 1.07 c |
| COS | 3.65 ± 0.19 a | 3.42 ± 0.53 b | 2.2 ± 0.08 ab | 15.11 ± 0.21 b | 0.62 ± 0.02 a | 17.85 ± 0.31 b | |
| CK | 3.03 ± 0.12 b | 3.97 ± 0.15 a | 2.4 ± 0.18 a | 16.66 ± 0.08 a | 0.63 ± 0.01 a | 23.31 ± 1.32 a | |
| Variety (V) | ** | ** | ** | ** | ** | ** | |
| Treatment (T) | ** | ** | ** | ** | ** | ** | |
| V × T | * | ns | ns | ** | ns | * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, H.; Wang, M.; Zhou, S.; Chen, K.; Wang, L.; Yi, Z.; Bai, L.; Zhang, Y. Chitosan Oligosaccharides Mitigate Flooding Stress Damage in Rice by Affecting Antioxidants, Osmoregulation, and Hormones. Antioxidants 2024, 13, 521. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050521
Lu H, Wang M, Zhou S, Chen K, Wang L, Yi Z, Bai L, Zhang Y. Chitosan Oligosaccharides Mitigate Flooding Stress Damage in Rice by Affecting Antioxidants, Osmoregulation, and Hormones. Antioxidants. 2024; 13(5):521. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050521
Chicago/Turabian StyleLu, Haoyu, Mei Wang, Shangfeng Zhou, Ke Chen, Lifeng Wang, Zhenxie Yi, Lianyang Bai, and Yuzhu Zhang. 2024. "Chitosan Oligosaccharides Mitigate Flooding Stress Damage in Rice by Affecting Antioxidants, Osmoregulation, and Hormones" Antioxidants 13, no. 5: 521. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050521