
Sarcopenia as a Risk Factor for Alzheimer’s Disease: Genetic and Epigenetic Perspectives

1
Centre for Health and Life Sciences, Coventry University, Coventry CV1 5FB, UK
2
School of Life, Health and Chemical Sciences, Open University, Milton Keynes MK7 6AA, UK
*
Author to whom correspondence should be addressed.
Genes 2024, 15(5), 561; https://0-doi-org.brum.beds.ac.uk/10.3390/genes15050561
Submission received: 28 March 2024
/
Revised: 24 April 2024
/
Accepted: 25 April 2024
/
Published: 27 April 2024
(This article belongs to the Section Molecular Genetics and Genomics)
Abstract
:Sarcopenia, defined as the age-associated loss of muscle mass and increased fragility with age, is increasing worldwide. The condition often precedes the development of Alzheimer’s disease, thereby decreasing the levels of mobility and physical activity in those affected. Indeed, the loss of muscle mass has, in some studies, been associated with an increased risk of Alzheimer’s disease and other dementias. However, a detailed understanding of the interplay between both conditions is not available and needs to be thoroughly addressed. In the following review, we focus on several genes, specifically APOE, BDNF, ACE, FTO, and FNDC5, that have been associated with both conditions. We also discuss the epigenetic regulation of each of these genes along with non-coding RNAs (ncRNAs) that may have a role in the development of both the sarcopenic and Alzheimer’s disease phenotypes. Finally, we assert that the application of systems biology will unravel the relationship between sarcopenia and Alzheimer’s disease and believe that the prevention of muscle loss in older age will reduce the incidence of debilitating cognitive decline.
1. Background
Sarcopenia, defined as the age-related loss of skeletal muscle and function [1], is known to predispose to an increased risk of falls and progressive loss of mobility with advancing years [2,3]. The condition is typically divided into two sub-categories whereby primary sarcopenia is an age-associated malady with no other evident causes and secondary sarcopenia is linked to causal factors, such as inflammatory disease, organ failure, and reduced physical activity [4,5].
The estimated prevalence of sarcopenia varies considerably and has been shown to range from 1–29% in community-dwelling older adults, 14–33% in elders within long-term care facilities, and 10% in those in an acute care setting [6]. Due to an aging population, the prevalence of sarcopenia is likely to increase as the percentage of the population over the age of 60 increases. For example, the World Health Organization (WHO) has projected that, by 2050, 2.1 billion people will pass their 60th year, inferring that over 200 million people will become sarcopenic in the next 40 years [1].
In addition to an increased fall risk and reduced mobility, evidence suggests that sarcopenia is associated with both Alzheimer’s disease (AD) and cognitive decline [7]. For example, Liu and coworkers recently showed that measures of reduced muscle function, including both lean mass and grip strength, correlate with declines in both cognition and memory. They also showed that lower muscle mass is associated with an increased degree of medial temporal lobe atrophy [8]. Likewise, sarcopenia was recently found to be prevalent in both AD patients and those with Lewy body dementia, where it manifested itself as both reduced muscle strength and slower gait speed [9]. Lower scores for the Mini-Mental State Examination (MMSE) and activities of daily living (ADL) were also evident in AD patients with sarcopenia compared to non-sarcopenic AD patients [10]. Furthermore, findings from a large UK Biobank-based study involving data from more than 190, 000 participants, showed that poor hand grip strength directly associates with reduced neurocognitive health and risk of dementia [11]. The link between sarcopenia and/or muscle mass (or function) and AD is, however, ambiguous in studies where fat-to-muscle ratios (FMRs) have been studied. For example, high whole-body FMRs apparently lower AD risk in men but have no effect on the risk of vascular dementia in both sexes [12]. Interestingly, leg FMRs exert a more pronounced inverse association on all causes of dementia risk in men and women [12]. Such data seem somewhat perplexing, as high FMRs are linked to an increased risk of type 2 diabetes [12], which is an independent risk factor for AD [13].
If sarcopenia has an impact on predisposition to AD and the associated dementias, then it is plausible to assume that genetic risk factors for sarcopenia might also exacerbate the risk of AD. In the paragraphs below, we highlight five genes that have been associated with both sarcopenia and AD. We discuss how genetic and epigenetic variability within these genes might modify the risk of both conditions but also spotlight areas where the association is less clear. We also discuss some ncRNAs that appear dysregulated in sarcopenia and AD, the emerging role of network biology, and how this will likely explain overlapping pathways in both conditions.
2. Genetics and Epigenetics
The human APOE gene resides on chromosome 19 and encodes a protein of 299 amino acids [14], which acts as a component of circulating lipoproteins and is crucial for the transport of lipids and cholesterol between cells of different organs [15]. The APOE gene is polymorphic with three well-studied allelic forms (ε2, ε3, and ε4) that lead to amino acid substitutions at positions 112 and 158. These changes result in three common isoforms of APOE, designated as apoE2, apoE3, and apoE4, respectively [16]. The APOE ε4 variant is a strong risk factor for both late-stage AD [17] and low-grade inflammation [17]. Interestingly, inflammation is also a risk factor for sarcopenia [18]. Although the exact mechanism whereby the APOE ε4 variant predisposes to AD is unclear, recent work suggests a paradoxical process for a neurodegenerative disease whereby the APOE4 isoform increases certain brain signaling pathways and synapse density, presumably by a mechanism involving enhanced precursor protein expression [15]. The APOE ε4 variant also predisposes to slower walking speed in individuals with mild cognitive impairments (MCI) [19], and slow walking speed is an underlying feature of sarcopenia [20]. However, and somewhat surprisingly, there is evidence that, in individuals with normal BMI, the APOE ε4 variant predisposes to higher lean body mass, and in young individuals’ certain aspects of the condition appear enhanced in APOE ε4 carriers [21]. Collectively, these data do not support a firm role for the APOE ε4 variant as predisposing to AD in sarcopenic individuals. However, it is important to mention that the influence of the APOE ε4 variant on human phenotypes appears to work through an epigenetic mechanism, as the presence of the ε4 allele changes the DNA methylation pattern of the APOE gene [22]. Specifically, the ε4 allele adds an additional CpG site to a 12 bp region that already contains 4 CpG sites. Conversely, the ε2 allele removes a CpG site and opens a 33 bp region free of CpG sites [22]. Furthermore, there appears to be a role for CpG islands within the APOE gene to act as signals for protein binding and chromatin remodeling [23], and this might also affect the risk of both sarcopenia and AD with advancing age. Therefore, we suggest that studies into the effect of the ε4 allele on muscle- and/or neurocognitive-related conditions should focus more on its epigenetic influences as opposed to a simple fixation on possession of (or lack of) the ε4 allele itself.
Brain-derived neurotrophic factor (BDNF) is a member of the neurotrophin family, with a complex role in supporting the health and longevity of neurons [24]. BDNF is expressed in various brain regions, particularly the hippocampus [25], along with the prostate [26] and skeletal muscle [27]. BDNF has an important role in long-term memory formation and storage [28], and evidence shows that exercise increases the expression of BDNF. For example, intense resistance exercise in both sexes has been shown to increase circulating BDNF levels immediately and 30 min after the intervention [29]. Furthermore, Lira and colleagues [30] found that lower-body resistance exercise was associated with higher BDNF levels compared to full- and upper-body exercise. However, some investigations have not found an increase in BDNF following resistance exercise [31]. For aerobic exercise, even in older-age subjects (65–74 years), significant increases in BDNF have been observed following 12 weeks of treadmill walking [32] and low-intensity prolonged cycling also upregulates BDNF [33].
Interestingly, a link between lower levels of BDNF and sarcopenia is evident in the literature. For example, Roh and co-workers [34] showed that reduced plasma BDNF levels were associated with both sarcopenia and dementia in a cohort of community-dwelling adults aged 70–84. Likewise, in sarcopenic patients with Parkinson’s disease [35] or comorbid with respiratory dysfunction, lower levels of BDNF are reported [36]. A recent meta-analysis showed that serum BDNF was lower in those with Alzheimer’s disease compared to healthy controls [37]. However, in peripheral blood samples, Kim and colleagues found that BDNF was lower in AD patients with the lowest MMSE scores, but in early AD, BDNF was higher than in controls [38].
Although, in general, it seems that lower BDNF is associated with both sarcopenia and AD, it should be noted that expression of BDNF is modified by the presence of the rs6265 (G196A or Val66Met) variant within exon XI of the gene [39]. The rs6265 variant is known to influence several downstream functions of BDNF, such as hippocampal function and episodic memory [39]. Furthermore, the presence of the A allele removes a CpG binding site within the gene that impacts expression [39]. Importantly, the extent of DNA methylation within promoter regions of the BDNF gene has been shown to impact cognitive performance in individuals with major depressive disorder [40]. The role of BDNF methylation in the development of AD has also been demonstrated by the work of Xie and co-workers. These authors conducted a 5-year longitudinal study on epigenetic changes in patients with mild cognitive impairment who converted to AD. They measured CpG site methylation at four locations within two promoter regions of the BDNF gene and found that elevated methylation of a site within the promoter IV area significantly (HR = 3.51, p = 0.013) associated with the conversion of MCI to AD [41].
The Angiotensin-converting enzyme (ACE) gene is found on chromosome 17q23 and consists of 26 exons with 25 introns. The gene itself contains at least 160 variant sites and encodes two isoforms with an important role in regulating blood pressure. Specifically, ACE converts angiotensin I to angiotensin II, and this has a vasoconstrictive effect on blood vessels and, therefore, reverses the effect of low blood pressure [42]. The common ACE I/D variant that resides within intron 17 of the gene has been associated with a broad spectrum of phenotypes and has long been considered an important factor related to both strength and endurance [43]. However, although the I/D variant has been associated with sarcopenia, the direction of association is still unclear. For example, the II genotype has been found to predispose to sarcopenia in a Brazilian population [44] but in elderly Indonesians, it was the DD genotype that was associated with the same condition [45]. Furthermore, in women, the DD genotype seems to protect from sarcopenic obesity [46]. As with sarcopenia, the role of the ACE I/D variant as a risk factor for AD is somewhat opaque, but a recent meta-analysis, using data from 65 studies, found some evidence to suggest that the I allele increased risk [47]. This finding is consistent with an older, large-scale study that also found that the I allele increased the risk of AD across diverse populations [48]. Interestingly, and perhaps inconsistent with the role of the I allele as a risk factor for AD, ACE D allele carriers with mild cognitive impairments (MCI) perform worse on auditory–verbal learning tests compared to those without a D allele [49]. A similar association between D allele carriage and recall in MCI patients was also found by Zhang and colleagues [50].
It is important to recognize that the regulation of the ACE gene is under strong epigenetic control via DNA methylation of CpG sites within its promoter. ACE expression is also controlled by histone modifications [51]. Indeed, the methylation status of the ACE gene has an indirect and possibly profound effect on the stress response and predisposition to depression, as it can modulate the levels of cortisol [52]. Elevated cortisol has been linked to both sarcopenia [53] and AD [54]. Hence it seems that the methylation status of the ACE gene may be more important than certain sequence variants in influencing a range of physiological phenotypes [55].
The FTO (fat mass and obesity-associated protein) gene resides on chromosome 16 in humans and encodes a non-heme dioxygenase Fe(II) and 2-oxoglutarate-dependent dioxygenase [56], which functions as a mRNA demethylase [57]. In recent years, variation within the FTO gene has been studied in a wide variety of phenotypes related to weight gain [58,59] cardiovascular fitness [60], and elite sporting ability [61]. Shu Ran and colleagues (2020) used a genome-wide association (GWAS) approach in a large Caucasian cohort to identify 29 single nucleotide polymorphisms (SNPs) associated with sarcopenia [62]. Furthermore, the role of FTO SNPs in sarcopenia has also been demonstrated in COPD patients by Attaway and coworkers using a GWAS design followed up with in vitro FTO knockdown experiments. In their studies, they found that FTO knockdown in mice yielded a sarcopenic profile that was also exacerbated by hypoxia [63]. Interestingly, Khanal and colleagues (2020) have shown that the FTO rs9939609 variant seems to be associated with sarcopenia, but the association was dependent on the definition used to describe the condition. Specifically, the variant modified the risk of sarcopenia when defined by both the percentage of muscle mass and the skeletal muscle mass index, but the association disappeared when using a definition for sarcopenia based on the European Working Group on Sarcopenia in Older People (EWGSOP) [64]. This clearly shows the necessity of using a standardized set of criteria for disease definition across all studies.
Variation within the FTO gene seems also to impact on the risk of AD. Using a cohort of nearly 2000 Caucasian cases and controls, the rs6499640 SNP within intron 1 of the gene was found to significantly associate with AD, while the rs10852521, rs16945088, and rs8044769 SNPs approached a significant p value for association [65]. The same authors also found that in Hispanics, the rs17219084, rs11075996, and rs11075997 SNPs were significantly associated with the disease. Likewise, a significant association between the FTO rs9939609 variant (AA genotype) and both AD and dementia was demonstrated by Keller and colleagues as part of the Kungsholmen project [66]. Furthermore, these authors also discovered an interaction effect between the FTO AA and APOE ϵ4 genotypes that increased the risk of both AD and dementia independent of other potentially confounding factors, such as diabetes, physical activity, and BMI [66].
It seems then that variation within the FTO gene can modify the risk of both sarcopenia and AD. However, although most studies have focused on population-based cohort studies, the effect of epigenetic modification of the FTO gene and the risk of both conditions has largely been ignored. Interestingly, the FTO gene is regulated at the epigenetic level by increased CpG site methylation/demethylation within the promoter by such factors as smoking, dietary triglycerides, curcumin, and Vitamin B12 [67]. Furthermore, promoter FTO methylation levels were shown to inversely correlate with birth weight in a Chinese population [68]. Indeed, a detailed understanding of how FTO-promoter methylation impacts on lean muscle mass and risk of AD, as a function of age, would prove useful.
Although a detailed commentary on the myokines is beyond the scope of this paper, we assert that these molecules and their aberrant expression may have a significant role to play in the development of both sarcopenia and AD. The myokines, which consist of a large group of small cytokine-based peptides, are secreted by muscle cells during the process of muscle contraction [69]. Indeed, this phenomenon is one way in which muscles communicate with other tissues of the body and points also towards an endocrine role for muscle that perhaps has previously been overlooked [69]. Irisin, formally discovered in 2012 [70], is a myokine that activates a range of signaling pathways in a variety of different tissues, including the mitogen-activated protein kinase (MAPK) pathway in neuronal tissue, the ERK1/2 pathways in cardiac microvascular endothelial cells [71], and the activation of telomerase in hepatocytes [72]. Circulating irisin levels have been shown to be lower in sarcopenic patients [73,74], and resistance training upregulates irisin via the AMPK/PGC-1α/irisin signaling pathway [75]. There is now growing interest in the role of Irisin in AD as the molecule has been shown to potentially protect from the deposition of amyloid-β (Aβ) protein in the brain, a key molecular signature of AD pathology, by upregulating astrocytic release of Aβ-degrading enzymes [76]. Indeed, this may be a mechanism whereby exercise protects from the risk of AD [76]. Furthermore, elevated irisin has been associated with improved global cognition and memory in adults at risk of dementia following ten weeks of physical or cognitive training [77].
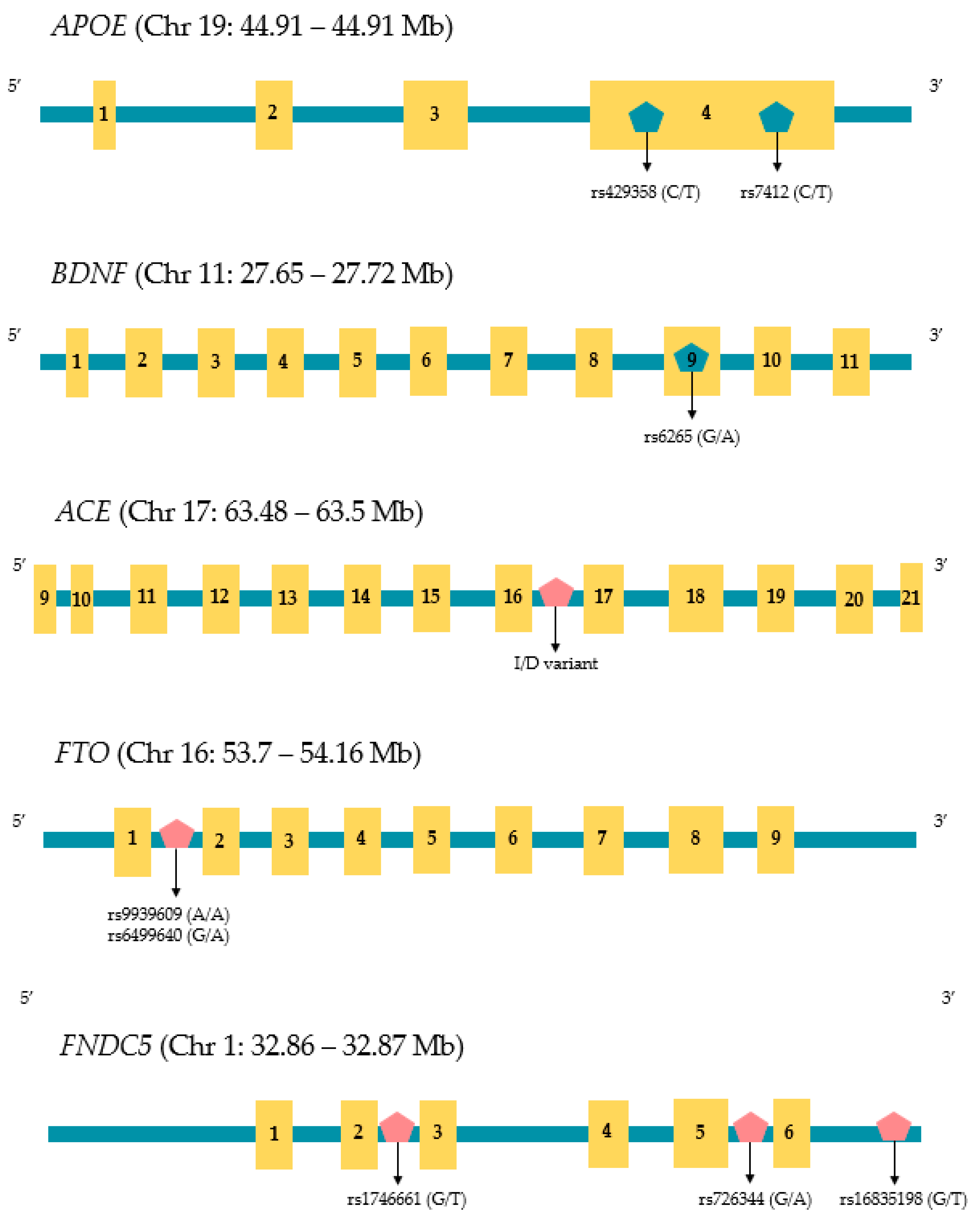
In humans, irisin is encoded by the FNDC5 (Fibronectin type III Domain Containing 5) gene that is found on chromosome 1. There are three variants of FNDC5 distinguished by sequences that code for different signal peptides and C-terminal domain regions [78] Variation within the FNDC5 promoter has been associated with a variety of phenotypic changes. For example, Lima Filho and colleagues reported that the rs1746661 variant (T allele) influences glucose metabolism within the brains of elderly individuals, which affects the risk of AD [79]. Using a mouse model, Madhu and coworkers have shown that FNDC5 knockout reduces the benefits of exercise-induced cognitive enhancement, which supports the role of irisin in protecting from AD [80]. Furthermore, two polymorphisms within the FNDC5 promoter (rs16835198 and rs726344) are both known to impact insulin sensitivity [81], and insulin sensitivity is a contributing factor for developing sarcopenia [82,83]. In addition to FNDC5 sequence variation, the gene is controlled epigenetically by a large CpG site, located at −52~−442 bp within two of the FNDC5 genes (variant 2 and 3), and methylation of this region has a significant effect on the expression of the gene. [78]. However, to our knowledge, a detailed understanding of the methylation status of this CpG site and its influence on both sarcopenia and AD risk has yet to be established. The structure, size, and location of relevant SNPs discussed in this review are shown in Figure 1.
Noncoding RNAs (ncRNAs), such as microRNAs (miRNAs) and long noncoding RNAs (lncRNAs), do not code for proteins but regulate gene expression through mRNA binding and transcriptional silencing, in the case of miRNAs [84]. In addition to mRNA binding, lncRNAs also regulate the overall process of gene expression through multiple regulatory processes, including the regulation of both histone modification and DNA methylation [85]. Lnc RNAs also act as protein scaffolds that can bring together multiple proteins to enhance biological functions [85]. There are multiple studies that show associations between ncRNAs and sarcopenia. For example, Faraldi and coworkers recently reported the downregulation of hsa-miR-221-3p, hsa-miR-374b-5p, hsa-miR-146a-5p, hsa-miR-126-5p, and hsa-miR-425-5p and the upregulation of hsa-miR-145-5p and hsa-miR-25-3p in post-menopausal women with muscle wasting [86]. Sarcopenic patients have also been shown to have downregulated levels of miR-133a, miR-133b, miR-206, miR-208b, and miR-499 [87]. Indeed, the role of ncRNAs in sarcopenia is further strengthened by the observation that certain ncRNAs, such as miR-155-5p, miR-421-3p, miR-425-5p, and miR-495-3p, appear repressed in those with the condition, but upon appropriate exercise intervention, expression levels return to normal [88]. Altered expression of ncRNAs maybe useful for the diagnosis and management of sarcopenia. For instance, the levels of circulating miR-1-3p, which can activate components of the Akt/mTOR pathway, are elevated in sarcopenic patients with heart disease and may be useful as a biomarker for the prediction of sarcopenia in these individuals [89]. Likewise, Valášková and colleagues [86] have shown that a profile of multiple ncRNAs, including miRNA-1, miRNA-29a and miRNA-29b, are increased in older individuals (aged 55-80 years) with poor muscle performance, whereas they detected decreased expression of miRNA-206, miRNA-133a, miRNA-133b, miRNA-208b, and miRNA-499 correlated with the same phenotype [86]. Indeed, the authors speculated that their data may be useful in the development of miRNA biomarkers for sarcopenia.
It seems likely then that ncRNAs may have a role to play in sarcopenia. However, it is more difficult to ascertain whether these molecules have a causative role in the condition or whether they are simply an artifact of muscle atrophy or other degenerative process during the aging process. Furthermore, is it possible that changes in ncRNAs that might predispose to sarcopenia may also increase the risk of AD? Indeed, many of the ncRNAs mentioned above (that have a role in sarcopenia) are also associated with AD. For example, the levels of hsa-miR-126-5p were shown to be altered in neutrophil-derived microvesicles of AD patients compared to controls, leading to possible blood–brain barrier dysfunctions that may predispose to AD [90]. Yuan and colleagues used mechanistic studies to establish that upregulation of hsa-miR-425-5p promotes tau phosphorylation and apoptosis in AD [91]. Indeed, tau hyperphosphorylation is a hallmark of AD pathology [92]. Interestingly, upregulation of has-miR-145p was recently shown to be a predictor of both AD and mild cognitive impairment (MCI) in a small cohort of patients. The same study found that hsa-miR-145 also improved the predicted performance of AD patients on the Mini-Mental State Examination (MMSE) [93]. Furthermore, both hsa-miR-29a and hsa-miR-29b have been shown to be altered in AD cohorts. Specifically, Peña-Bautista and coworkers [94] found upregulated levels of hsa-miR-29a that, in combination with several other ncRNAs, tentatively associated with AD. However, hsa-miR-29a was found to be downregulated in Egyptian patients with AD [95]. Studies into the interaction of lncRNAs with miRNAs have shown that hsa-miR-155-5p may play a role in neuron projection development and neuron morphogenesis in AD [96]. Furthermore, the authors infer that hsa-miR-155-5p may have potential as a clinical biomarker for AD [96]. As with sarcopenia, we must also be cautious in inferring direct causative roles for ncRNAs in AD. Furthermore, although we have highlighted overlaps between ncRNAs that may appear dysregulated in both conditions, definitive mechanisms for how these molecules enhance the risk of both are not yet understood. It is important to remember that ncRNAs, for example, miRNAs, often have multiple target mRNAs that they can regulate, and it is possible that some observations of their altered expression in disease processes (like sarcopenia and AD) may be coincidental findings or due to confounding factors within patient cohorts. A summary of risk genes, non-coding RNAs, and microRNAs discussed in this review is shown in Table 1.
In the above paragraphs, we have limited discussion to how variability (both genetic and epigenetic) within five genes (APOE, BDNF, ACE, FNDC5, and FTO) might impact on both sarcopenia and AD risk. We have also included a commentary on the role of ncRNAs that overlap in both conditions. However, these are complex conditions and unlikely to be solely due to changes within a small number of genes per se. To support this view, it has now been demonstrated that 270 differentially methylated regions of the genome exist between patients with AD compared to controls. Furthermore, the methylation changes were shown to profoundly impact protein networks and transcriptomic, proteomic, and chromatin accessibility of specific regions of the brain in AD patients [97]. However, overlapping epigenetic and interatomic changes between AD and sarcopenia are only just starting to emerge. For example, it was recently shown, using protein-interaction networks, that the dysregulated expression of five overlapping mitochondrial hub genes (NDUFAB1, UQCRC1, UQCRFS1, NDUFS3, and MRPL15) and associated networks may be important in both musculoskeletal aging (sarcopenia) and AD [98]. Indeed, it is the authors’ assertion that a systems-biology-based approach to identify common aspects, and potentially new biomarkers for both conditions, should now be a priority. This approach has been adopted by Yedigaran and colleagues where divergent and shared miRNA regulatory pathways in both cachexia and sarcopenia were identified [99] but extending this work to overlapping miRNA networks between sarcopenia and AD pathways, especially in relation to physical activity, might also prove beneficial. Finally, when it comes to both sarcopenia and AD, we are faced with a possible cause-and-effect question. Specifically, does sarcopenia, which is postulated to be due, at least in part, to systemic inflammation [100], promote a cascade of events that leads to AD, or does AD lead to sarcopenia purely by a reduction in physical activity, and subsequent loss of muscle mass, in those affected? Whichever of these two scenarios is at play, an integrated genetic, epigenetic, and systems-biology-based signature would help to identify at-risk individuals and allow for personalized interventions.
Author Contributions
S.M.R. and K.J.A.O. researched the content of this article, S.M.R. drafted the initial manuscript and both S.M.R. and K.J.A.O. edited the initial draft and agreed to the final version. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Santilli, V.; Bernetti, A.; Mangone, M.; Paoloni, M. Clinical definition of sarcopenia. Clin. Cases Miner. Bone Metab. 2014, 11, 177–180. [Google Scholar] [PubMed]
- Han, D.; Wu, W.; Hsu, P.; Chang, H.; Huang, K.; Chang, K. Sarcopenia is associated with increased risks of rotator cuff tendon diseases among community-dwelling elders: A cross-sectional quantitative ultrasound study. Front. Med. 2021, 8, 630009. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.H.-T.; U, K.P.; Yiu, T.; Ong, M.T.-Y.; Lee, W.Y.-W. Sarcopenia: Current treatments and new regenerative therapeutic approaches. J. Orthop. Transl. 2020, 23, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyere, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Writing Group for the European Working Group on Sarcopenia in Older People 2 (EWGSOP2), and the Extended Group for EWGSOP2. Sarcopenia: Revised European Consensus on Definition and Diagnosis. Age Ageing 2019, 48, 601. [Google Scholar] [CrossRef] [PubMed]
- Avgerinou, C. Sarcopenia: Why it matters in general practice. Br. J. Gen. Pract. 2020, 70, 200–201. [Google Scholar] [CrossRef]
- Delmonico, M.J.; Beck, D.T. The current understanding of sarcopenia. Am. J. Lifestyle Med. 2016, 11, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Beeri, M.S.; Leugrans, S.E.; Delbono, O.; Bennett, D.A.; Buchman, A.S. Sarcopenia is associated with incident Alzheimer’s dementia, mild cognitive impairment, and cognitive decline. J. Am. Geriatr. Soc. 2021, 69, 1826–1835. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.W.; Li, M.; Zhu, J.T.; Zhang, Y.C.; Wu, Y.H.; Liu, C.F.; Hu, H. Correlation of muscle strength with cognitive function and medial temporal lobe atrophy in patients with mild to moderate Alzheimer’s disease. Zhonghua Yi Zue Za Zhi 2022, 102, 2786–2792. [Google Scholar] [CrossRef] [PubMed]
- Dost, F.S.; Bulut, E.A.; Dokuzlar, O.; Kaya, D.; Mutlay, F.; Gurel, B.H.Y.; Isik, A.T. Sarcopenia is as common in older patients with dementia with Lewy bodies as it is in those with Alzheimer’s disease. Geriatr. Gerontol. Int. 2022, 22, 418–424. [Google Scholar] [CrossRef]
- Dost, F.S.; Erken, N.; Ontan, M.S.; Bulut, E.A.; Kaya, D.; Kocyigit, S.E.; Dokuzlar, O.; Aydin, A.E.; Isik, A.T. Muscle strength Seems to be Related to The Functional Status and Severity of Dementia in Older Adults with Alzheimer’s Disease. Curr. Aging Sci. 2023, 16, 75–83. [Google Scholar] [CrossRef]
- Duchowny, K.A.; Ackley, S.F.; Brenowitz, W.D.; Wang, J.; Zimmerman, S.C.; Caunca, M.R.; Glymour, M.M. Associations between Handgrip strength and dementia risk, cognition, and neuroimaging outcomes in the UK biobank cohort study. JAMA Netw. Open 2022, 5, 75–83. [Google Scholar] [CrossRef]
- Wang, W.; Luo, Y.; Zhuang, Z.; Song, Z.; Huang, N.; Li, Y.; Dong, X.; Xiao, W.; Zhao, Y.; Huang, T. Total and regional fat-to-muscle mass ratio and risks of incident all-cause dementia, Alzheimer’s disease, and vascular dementia. J. Cachexia Sarcopenia Muscle 2022, 13, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Čater, M.; Hölter, S.M. A pathophysiological intersection of diabetes and Alzheimer’s disease. Int. J. Mol. Sci. 2022, 23, 11562. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.A.; Laurent, B.; Plourde, M. APOE and Alzheimer’s disease: From lipid transport to physiopathology and therapeutics. Front. Neurosci. 2021, 15, 630502. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Mahley, R.W. Apolipoprotein E: Structure and function in lipid metabolism, neurobiology, and Alzheimer’s diseases. Neurobiol. Dis. 2014, 72, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Ghebranious, N.; Ivacic, L.; Mallum, J.; Dokken, C. Detection of ApoE E2, E3 and E4 alleles using MALDI-TOF mass spectrometry and the homogeneous mass-extend technology. Nucleic Acids Res. 2005, 33, e149. [Google Scholar] [CrossRef] [PubMed]
- Di Battista, A.M.; Heinsinger, N.M.; Rebeck, G.W. Alzheimer’s disease genetic risk factor APOE-ε4 also affects normal brain function. Curr. Alzheimer Res. 2016, 13, 1200–1207. [Google Scholar] [CrossRef] [PubMed]
- Dalle, S.; Rossmeislova, L.; Koppo, K. The role of inflammation in age-related sarcopenia. Front. Physiol. 2017, 8, 1045. [Google Scholar] [CrossRef] [PubMed]
- Doi, T.; Shimada, H.; Makizako, H.; Tsutsumimoto, K.; Uemura, K.; Suzuki, T. Apolipoprotein E genotype and physical function among older people with mild cognitive impairment. Geriatr. Gerontol. Int. 2014, 15, 422–427. [Google Scholar]
- Bhasin, S.; Travison, T.G.; Manini, T.M.; Patel, S.; Pencina, K.M.; Fielding, R.A.; Magaziner, J.M.; Newman, A.B.; Kiel, D.P.; Cooper, C.; et al. Sarcopenia definition: The position statements of the sarcopenia definition and outcomes consortium. J. Am. Geriatr. Soc. 2020, 68, 1410–1418. [Google Scholar] [CrossRef]
- Matura, S.; Prvulovic, D.; Hartmann, D.; Scheibe, M.; Sepanski, B.; Butz, M.; Oertel-Knöchel, V.; Knöchel, C.; Karakaya, T.; Fußer, F.; et al. Age-Related Effects of the Apolipoprotein E gene on Brain Function. J. Alzheimer’s Dis. 2016, 52, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Foraker, J.; Millard, S.P.; Leong, L.; Thomson, Z.; Chen, S.; Keene, C.D.; Bekris, L.M.; Yu, C.E. The APOE Gene is Differentially Methylated in Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 48, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, M.; Dibello, V.; Sardone, R.; Castellana, F.; Zupo, R.; Lampignano, L.; Bortone, I.; Daniele, A.; Bellomo, A.; Solfrizzi, V.; et al. The Impact of Apolipoprotein E (APOE) Epigenetics on Aging and Sporadic Alzheimer’s Disease. Biology 2023, 12, 1529. [Google Scholar] [CrossRef] [PubMed]
- Acheson, A.; Conover, J.C.; Fandl, J.P.; DeChiara, T.M.; Russell, M.; Thadani, A.; Squinto, S.P.; Yancopoulos, G.D.; Lindsay, R.M. A BDNF autocrine loop in adult sensory neurons prevents cell death. Nature 1995, 374, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Nabeshima, T. Brain-derived neurotrophic factor/TrkB signaling in memory processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Malekan, M.; Nezamabadi, S.S.; Samami, E.; Mohebalizadeh, M.; Saghazadeh, A.; Rezaei, N. BDNF and its signaling in cancer. J. Cancer Res. Clin. Oncol. 2022, 149, 2621–2636. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Edgett, B.A.; Tschakovsky, M.E.; Gurd, B.J. Fasting and exercise differentially regulate BDNF mRNA expression in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2015, 40, 96–98. [Google Scholar] [CrossRef] [PubMed]
- Bekinschtein, P.; Cammarota, M.; Katche, C.; Slipczuk, L.; Rossato, J.I.; Goldin, A.; Izquierdo, I.; Medina, J.H. BDNF is essential to promote persistence of long-term memory storage. Proc. Natl. Acad. Sci. USA 2008, 105, 2711–2716. [Google Scholar] [CrossRef] [PubMed]
- Marston, K.J.; Newton, M.J.; Brown, B.M.; Rainey-Smith, S.R.; Bird, S.; Martins, R.N.; Peiffer, J.J. Intense resistance exercise increases peripheral brain-derived neurotrophic factor. J. Sci. Med. Sports 2017, 20, 899–903. [Google Scholar] [CrossRef]
- Lira, F.S.; de Freitas, M.C.; Gerosa-Neto, J.; Cholewa, J.M.; Rossi, F.E. Comparison between Full-Body vs. Split-Body Resistance Exercise on the Brain-Derived Neurotrophic Factor and Immunometabolic Response. J. Strength Cond. Res. 2020, 34, 3094–3102. [Google Scholar] [CrossRef]
- Babiarz, M.; Laskowski, R.; Grzywacz, T. Effects of strength training on BDNF in healthy young adults. Int. J. Environ. Res. Public Health 2022, 19, 13795. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Roh, H.T. Effects of Exercise Training on Neurotrophic Factors and Blood–Brain Barrier Permeability in Young-old and Old-old Women. Int. J. Environ. Res. Public Health 2022, 19, 16896. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, T.D.; Cotter, J.D.; Ainslie, P.N.; Abraham, W.C.; Mockett, B.G.; Campbell, H.A.; Jones, E.M.; Jenkins, E.J.; Thomas, K.N. Fasting for 20 h does not affect exercise-induced increases in circulating BDNF in humans. J. Physiol. 2023, 601, 2121–2137. [Google Scholar] [CrossRef] [PubMed]
- Roh, E.; Hwang, S.Y.; Song, E.; Park, J.M.; Yoo, J.H.; Baik, H.S.; Kim, M.; Won, W.C.; Choi, M.K. Association of plasma brain-derived neurotrophic factor levels and frailty in community-dwelling older adults. Sci. Rep. 2022, 12, 18605. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Iqbal, M.S.; Muhammad, T.; Qaisar, R. Evaluation of Sarcopenia Using Biomarkers of the Neuromuscular Junction in Parkinson’s Disease. J. Mol. Neurosci. 2022, 72, 820–829. [Google Scholar] [CrossRef]
- Lage, V.K.; de Paula, F.A.; Lima, L.P.; Santos, J.N.; dos Santos, J.M.; Viegas, Â.A.; da Silva, G.P.; de Almeida, H.C.; Rodrigues, A.L.; Leopoldino, A.A.; et al. Plasma levels of myokines and inflammatory markers are related with functional and respiratory performance in older adults with COPD and sarcopenia. Exp. Gerontol. 2022, 164, 111834. [Google Scholar] [CrossRef]
- Ng, T.K.S.; Ho, C.S.H.; Tam, W.W.S.; Kua, E.H.; Ho, R.C. Decreased serum brain-derived neurotrophic factor (BDNF) levels in patients with Alzheimer’s disease (AD): A systematic review and meta-analysis. Int. J. Mol. Sci. 2019, 20, 257. [Google Scholar] [CrossRef]
- Kim, B.Y.; Lee, S.H.; Graham, P.L.; Angelucci, F.; Lucia, A.; Pareja-Galeano, H.; Leyhe, T.; Turana, Y.; Lee, I.R.; Yoon, J.H.; et al. Peripheral brain-derived neurotrophic factor levels in Alzheimer’s disease and mild cognitive impairment: A comprehensive systematic review and meta-analysis. Mol. Neurobiol. 2017, 54, 7297–7311. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Hill, B.; Sims, N.; Heck, A.; Negron, M.; Lusk, C.; Galindo, C.L. Brain-derived neurotrophic factor RS6265 (val66met) single nucleotide polymorphism as a master modifier of human pathophysiology. Neural Regen. Res. 2023, 18, 102. [Google Scholar] [CrossRef]
- Ferrer, A.; Labad, J.; Salvat-Pujol, N.; Barrachina, M.; Costas, J.; Urretavizcaya, M.; de Arriba-Arnau, A.; Crespo, M.J.; Soriano-Mas, C.; Carracedo, A.; et al. BDNF genetic variants and methylation: Effects on cognition in major depressive disorder. Transl. Psychiatry 2019, 9, 265. [Google Scholar] [CrossRef]
- Xie, B.; Xu, Y.; Liu, Z.; Liu, W.; Jiang, L.; Zhang, R.; Cui, D.; Zhang, Q.; Xu, S. Elevation of peripheral BDNF promoter methylation predicts conversion from amnestic mild cognitive impairment to Alzheimer’s disease: A 5-year longitudinal study. J. Alzheimer’s Dis. 2017, 56, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Sharma, V.; Kaur, D.; Purkait, P. Angiotensin-converting enzyme (ACE) insertion/deletion (I/d) polymorphism as a conjoint regulator of coagulation, fibrinolytic, and raas pathway in infertility and associated pregnancy complications. J. Renin-Angiotensin-Aldosterone Syst. 2022, 2022, 1695769. [Google Scholar] [CrossRef] [PubMed]
- Puthucheary, Z.; Skipworth, J.R.A.; Rawal, J.; Loosemore, M.; Van Someren, K.; Montgomery, H.E. The Ace Gene and human performance: 12 years on. Sports Med. 2011, 41, 433–448. [Google Scholar] [CrossRef]
- da Silva, J.R.; Freire, I.V.; Ribeiro, Í.J.S.; dos Santos, C.S.; Casotti, C.A.; dos Santos, D.B.; Barbosa, A.A.; Pereira, R. Improving the comprehension of sarcopenic state determinants: An multivariate approach involving hormonal, nutritional, lifestyle and genetic variables. Mech. Ageing Dev. 2018, 173, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Handayani, M.D.N.; Sadewa, A.H.; Farmawati, A.; Rochmah, W. Deletion Polymorphism of Angiotensin-Converting Enzyme Gene Is Associated with Low Muscle Mass in Elderly People in Jakarta, Indonesia. Kobe J. Med. Sci. 2018, 64, 119–125. [Google Scholar] [PubMed Central]
- Blanco, C.R.; González, M.J.A.; Cruz, E.C.; Mallén, J.A.C.; Royo, I.A.; Laín, S.A. Sarcopenic Obesity in Elderly Women: Influence of I/D Polymorphism of the Angiotension Converter Enzyme. Nutr. Hosp. 2017, 34, 1099–1104. [Google Scholar] [CrossRef]
- Xin, X.-Y.; Lai, Z.-H.; Ding, K.-Q.; Zeng, L.-L.; Ma, J.-F. Angiotensin-converting enzyme polymorphisms and Alzheimer’s disease susceptibility: An updated meta-analysis. PLoS ONE 2021, 16, e0260498. [Google Scholar] [CrossRef]
- Lehmann, D.J.; Cortina-Borja, M.; Warden, D.R.; Smith, A.D.; Sleegers, K.; Prince, J.A.; van Duijn, C.M.; Kehoe, P.G. Large meta-analysis establishes the ACE insertion-deletion polymorphism as a marker of Alzheimer’s disease. Am. J. Epidemiol. 2005, 162, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Deng, L.; Bai, F.; Shi, Y.; Yu, H.; Xie, C.; Yuan, Y.; Zhang, Z. Genetic variation in angiotensin converting-enzyme affects the white matter integrity and cognitive function of amnestic mild cognitive impairment patients. J. Neurol. Sci. 2017, 380, 177–181. [Google Scholar] [CrossRef]
- Zhang, Z.; Deng, L.; Yu, H.; Shi, Y.; Bai, F.; Xie, C.; Yuan, Y.; Jia, J.; Zhang, Z. Association of angiotensin-converting enzyme functional gene I/D polymorphism with amnestic mild cognitive impairment. Neurosci. Lett. 2017, 514, 131–135. [Google Scholar] [CrossRef]
- Rivière, G.; Lienhard, D.; Andrieu, T.; Vieau, D.; Frey, B.M.; Frey, F.J. Epigenetic regulation of somatic angiotensin-converting enzyme by DNA methylation and histone acetylation. Epigenetics 2011, 6, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.; Ancelin, M.-L.; Ritchie, K.; Saffery, R.; Ryan, J. DNA methylation and genetic variation of the angiotensin converting enzyme (ACE) in depression. Psychoneuroendocrinology 2018, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.G.; Marques-Vidal, P.; Aubry-Rozier, B.; Papadakis, G.; Preisig, M.; Kuehner, C.; Vollenweider, P.; Waeber, G.; Hans, D.; Lamy, O. Diurnal salivary cortisol in sarcopenic postmenopausal women: The osteolaus cohort. Calcif. Tissue Int. 2021, 109, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Ouanes, S.; Popp, J. High cortisol and the risk of dementia and alzheimer’s disease: A review of the literature. Front. Aging Neurosci. 2019, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, S.M. Epigenetic regulation of the ACE gene might be more relevant to endurance physiology than the I/D polymorphism. J. Appl. Physiol. 2012, 112, 1082–1083. [Google Scholar] [CrossRef]
- Sanchez-Pulido, L.; Andrade-Navarro, M.A. The FTO (fat mass and obesity associated) gene codes for a novel member of the non-heme dioxygenase superfamily. BMC Biochem. 2007, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.; Girard, C.A.; Tung, Y.-C.L.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.H.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Bryl, E.; Szcześniewska, P.; Dutkiewicz, A.; Słopień, A.; Dmitrzak-Węglarz, M.; Hanć, T. FTO and MC4R polymorphisms, and selected pre-, peri- and postnatal factors as determinants of body mass index and fatness in children: A thorough analysis of the associations. J. Physiol. Anthr. 2023, 42, 29. [Google Scholar] [CrossRef] [PubMed]
- Usategui-Martín, R.; Pérez-Castrillón, J.L.; Briongos-Figuero, L.; Abadía-Otero, J.; Lara-Hernandez, F.; García-Sorribes, S.; Martín-Vallejo, J.; García-García, A.B.; Chaves, F.J.; Martín-Escudero, J.C. Genetic variants in obesity-related genes and the risk of osteoporotic fracture. Hortega Follow-Up Study. Front. Biosci. (Landmark Ed.) 2022, 27, 32. [Google Scholar] [CrossRef]
- Mier-Mota, J.; Ponce-González, J.G.; Perez-Bey, A.; Cabanas-Sánchez, V.; Veiga-Núñez, O.; Santiago-Dorrego, C.; Gómez-Gallego, F.; Castro-Piñero, J. Longitudinal effects of FTO gene polymorphism on body composition, cardiorespiratory fitness, physical activity, inflammatory markers, and cardiovascular risk in children and adolescents. “The UP & DOWN study”. Scand. J. Med. Sci. Sports 2023, 33, 2261–2272. [Google Scholar] [CrossRef]
- Heffernan, S.M.; Stebbings, G.K.; Kilduff, L.P.; Erskine, R.M.; Day, S.H.; Morse, C.I.; McPhee, J.S.; Cook, C.J.; Vance, B.; Ribbans, W.J.; et al. Fat mass and obesity associated (FTO) gene influences skeletal muscle phenotypes in non-resistance trained males and elite rugby playing position. BMC Genet. 2017, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Ran, S.; Jiang, Z.-X.; He, X.; Liu, Y.; Zhang, Y.-X.; Zhang, L.; Pei, Y.-F.; Zhang, M.; Hai, R.; Gu, G.-S.; et al. Replication of FTO Gene associated with lean mass in a Meta-Analysis of Genome-Wide Association Studies. Sci. Rep. 2020, 10, 5057. [Google Scholar] [CrossRef] [PubMed]
- Attaway, A.H.; Bellar, A.; Welch, N.; Sekar, J.; Kumar, A.; Mishra, S.; Hatipoğlu, U.; McDonald, M.; Regan, E.A.; Smith, J.D.; et al. Gene polymorphisms associated with heterogeneity and senescence characteristics of sarcopenia in chronic obstructive pulmonary disease. J. Cachexia Sarcopenia Muscle 2023, 14, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Khanal, P.; He, L.; Stebbings, G.; Onambele-Pearson, G.L.; Degens, H.; Williams, A.; Thomis, M.; Morse, C.I. Prevalence and association of single nucleotide polymorphisms with sarcopenia in older women depends on definition. Sci. Rep. 2020, 10, 2913. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Tosto, G.; Mayeux, R.; Luchsinger, J.A.; Alzheimer’s Disease Neuroimaging the NIA-LOAD/NCRAD Family Study Group; Alzheimer’s Disease Neuroimaging Initiative. Genetic Variants in the Fat and Obesity Associated (FTO) Gene and Risk of Alzheimer’s Disease. PLoS ONE 2012, 7, e50354. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.; Xu, W.; Wang, H.-X.; Winblad, B.; Fratiglioni, L.; Graff, C. The obesity related gene, FTO, interacts with APOE, and is associated with Alzheimer’s disease risk: A prospective cohort study. J. Alzheimer’s Dis. 2011, 23, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Popović, A.-M.; Turković, A.H.; Žuna, K.; Bačun-Družina, V.; Rubelj, I.; Matovinović, M. FTO Gene Polymorphisms at the Crossroads of Metabolic Pathways of Obesity and Epigenetic Influences. Food Technol. Biotechnol. 2023, 61, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-W.; Zhang, J.-T.; Cai, Q.-Y.; Zhang, H.-X.; Wang, Y.-H.; Yan, H.-T.; Wu, H.-M.; Yang, X.-J. Birth weight is associated with placental fat mass- and obesity-associated gene expression and promoter methylation in a Chinese population. J. Matern. Fetal Neonatal Med. 2016, 29, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Grzeszczuk, M.; Dzięgiel, P.; Nowińska, K. The Role of FNDC5/Irisin in Cardiovascular Disease. Cells 2024, 13, 277. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Yang, L.; Wang, T.; Zhang, J.; Li, T.; Ren, Y.; Wang, M.; Chen, X.; Lv, Y.; Wu, R. Irisin Improves Autophagy of Aged Hepatocytes via Increasing Telomerase Activity in Liver Injury. Oxid. Med. Cell Longev. 2020, 2020, 6946037. [Google Scholar] [CrossRef] [PubMed]
- Demir, L.; Oflazoğlu, U. The relationship between sarcopenia and serum irisin and TNF-α levels in newly diagnosed cancer patients. Support. Care Cancer 2023, 31, 586. [Google Scholar] [CrossRef] [PubMed]
- de Luis, D.; Primo, D.; Izaola, O.; Gómez, J.J.L. Role of irisin and myostatin on sarcopenia in malnourished patients diagnosed with GLIM criteria. Nutrition 2023, 120, 112348. [Google Scholar] [CrossRef] [PubMed]
- Zuo, C.; Ma, X.; Ye, C.; Zheng, Z.; Bo, S. Acute and chronic functional and traditional resistance training improve muscular fitness in young males via the AMPK/PGC-1α/irisin signaling pathway. Environ. Health Prev. Med. 2023, 28, 69. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kim, H.; Jedrychowski, M.P.; Bakiasi, G.; Park, J.; Kruskop, J.; Choi, Y.; Kwak, S.S.; Quinti, L.; Kim, D.Y.; et al. Irisin reduces amyloid-β by inducing the release of neprilysin from astrocytes following downregulation of ERK-STAT3 signaling. Neuron 2023, 111, 3619–3633. [Google Scholar] [CrossRef] [PubMed]
- Küster, O.C.; Laptinskaya, D.; Fissler, P.; Schnack, C.; Zügel, M.; Nold, V.; Thurm, F.; Pleiner, S.; Karabatsiakis, A.; von Einem, B.; et al. Novel Blood-Based Biomarkers of Cognition, Stress, and Physical or Cognitive Training in Older Adults at Risk of Dementia: Preliminary Evidence for a Role of BDNF, Irisin, and the Kynurenine Pathway. J. Alzheimers Dis. 2017, 59, 1097–1111. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Jeong, Y.J.; Song, I.-S.; Noh, Y.H.; Seo, K.W.; Kim, M.; Han, J. Glucocorticoid receptor positively regulates transcription of FNDC5 in the liver. Sci. Rep. 2017, 7, 43296. [Google Scholar] [CrossRef]
- Lima-Filho, R.; Benedet, A.L.; De Bastiani, M.A.; Povala, G.; Cozachenco, D.; Ferreira, S.T.; De Felice, F.G.; Rosa-Neto, P.; Zimmer, E.R.; Lourenco, M.V.; et al. Alzheimer’s Disease Neuroimaging Initiative. Association of the fibronectin type III domain-containing protein 5 rs1746661 single nucleotide polymorphism with reduced brain glucose metabolism in elderly humans. Brain Commun. 2023, 5, fcad216. [Google Scholar] [CrossRef]
- Madhu, L.N.; Somayaji, Y.; Shetty, A.K. Promise of irisin to attenuate cognitive dysfunction in aging and Alzheimer’s disease. Ageing Res. Rev. 2022, 78, 101637. [Google Scholar] [CrossRef]
- Staiger, H.; Böhm, A.; Scheler, M.; Berti, L.; Machann, J.; Schick, F.; Machicao, F.; Fritsche, A.; Stefan, N.; Weigert, C.; et al. Common genetic variation in the human FNDC5 locus, encoding the novel muscle-derived ‘browning’ factor irisin, determines insulin sensitivity. PLoS ONE 2013, 8, e61903. [Google Scholar] [CrossRef]
- Miao, Y.; Xie, L.; Song, J.; Cai, X.; Yang, J.; Ma, X.; Chen, S.; Xie, P. Unraveling the causes of sarcopenia: Roles of neuromuscular junction impairment and mitochondrial dysfunction. Physiol. Rep. 2024, 12, e15917. [Google Scholar]
- Guo, M.; Yao, J.; Li, J.; Zhang, J.; Wang, D.; Zuo, H.; Zhang, Y.; Xu, B.; Zhong, Y.; Shen, F.; et al. Irisin ameliorates age-associated sarcopenia and metabolic dysfunction. J. Cachexia Sarcopenia Muscle 2023, 14, 391–405. [Google Scholar] [CrossRef]
- Raleigh, S.M. Chapter 2: Noncoding RNAs. In Epigenetics of Exercise and Sports; Concepts, Methods and Current Research, 1st ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 25, ISBN 9780128206829. [Google Scholar]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef]
- Faraldi, M.; Sansoni, V.; Vitale, J.; Perego, S.; Gomarasca, M.; Verdelli, C.; Messina, C.; Sconfienza, L.M.; Banfi, G.; Corbetta, S.; et al. Plasma microRNA signature associated with skeletal muscle wasting in post-menopausal osteoporotic women. J. Cachexia Sarcopenia Muscle 2024, 25, 690–701. [Google Scholar] [CrossRef]
- Valášková, S.; Gažová, A.; Vrbová, P.; Koller, T.; Šalingová, B.; Adamičková, A.; Chomaničová, N.; Hulajová, N.; Payer, J.; Kyselovič, J. The severity of muscle performance deterioration in sarcopenia correlates with circulating muscle tissue-specific miRNAs. Physiol. Res. 2021, 70, S91–S98. [Google Scholar] [CrossRef]
- Agostini, S.; Mancuso, R.; Costa, A.S.; Guerini, F.R.; Trecate, F.; Miglioli, R.; Menna, E.; Arosio, B.; Clerici, M. Sarcopenia associates with SNAP-25 SNPs and a miRNAs profile which is modulated by structured rehabilitation treatment. J. Transl. Med. 2021, 19, 315. [Google Scholar] [CrossRef]
- Xu, R.; Cui, S.; Chen, L.; Chen, X.-C.; Ma, L.-L.; Yang, H.-N.; Wen, F.-M. Circulating miRNA-1-3p as Biomarker of Accelerated Sarcopenia in Patients Diagnosed with Chronic Heart Failure. Rev. Investig. Clin. 2022, 74, 276–283. [Google Scholar] [CrossRef]
- Vázquez-Villaseñor, I.; Smith, C.I.; Thang, Y.J.R.; Heath, P.R.; Wharton, S.B.; Blackburn, D.J.; Ridger, V.C.; Simpson, J.E. RNA-Seq Profiling of Neutrophil Derived Microvesicles in Alzheimer’s Disease Patients Identifies a miRNA Signature That May Impact Blood-Brain Barrier Integrity. Int. J. Mol. Sci. 2022, 23, 5913. [Google Scholar] [CrossRef]
- Yuan, J.; Wu, Y.; Li, L.; Liu, C. MicroRNA-425-5p promotes tau phosphorylation and cell apoptosis in Alzheimer’s disease by targeting heat shock protein B8. J. Neural Transm. 2020, 127, 339–346. [Google Scholar] [CrossRef]
- Singh, A.; Ansari, V.A.; Mahmood, T.; Hasan, S.M.; Wasim, R.; Maheshwari, S.; Akhtar, J.; Sheikh, S.; Vishwakarma, V.K. Targeting Abnormal Tau Phosphorylation for Alzheimer’s Therapeutics. Horm. Metab. Res. 2024, 13, a-2238-1384. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Wittens, M.M.J.; Engelborghs, S.; van Herwijnen, M.H.M.; Tsamou, M.; Roggen, E.; Smeets, B.; Krauskopf, J.; Briedé, J.J. Beyond CSF and Neuroimaging Assessment: Evaluating Plasma miR-145-5p as a Potential Biomarker for Mild Cognitive Impairment and Alzhei-mer’s Disease. ACS Chem. Neurosci. 2024, 15, 1042–1054. [Google Scholar] [CrossRef]
- Peña-Bautista, C.; Tarazona-Sánchez, A.; Braza-Boils, A.; Balaguer, A.; Ferré-González, L.; Cañada-Martínez, A.J.; Baquero, M.; Cháfer-Pericás, C. Plasma microRNAs as potential biomarkers in early Alzheimer disease expression. Sci. Rep. 2022, 12, 15589. [Google Scholar] [CrossRef] [PubMed]
- Abuelezz, N.Z.; Nasr, F.E.; Aal, W.M.A.; Molokhia, T.; Zaky, A. Sera miR-34a, miR-29b and miR-181c as potential novel diagnostic bi-omarker panel for Alzheimers in the Egyptian population. Exp. Gerontol. 2022, 169, 111961. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, Z.; Yu, Y.; Cao, J.; Qiao, Y.; Qiao, H.; Suo, G. Comprehensive analysis of the lncRNA-associated ceRNA network identifies neuroinflammation biomarkers for Alzheimer’s disease. Mol. Omics 2019, 15, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wang, M.; Guo, L.; Fullard, J.F.; Micallef, C.; Bendl, J.; Song, W.; Ming, C.; Huang, Y.; Li, Y.; et al. Genome-wide methylomic regulation of multiscale gene networks in Alzheimer’s disease. Alzheimer’s Dement. 2023, 19, 3472–3495. [Google Scholar] [CrossRef] [PubMed]
- Giannos, P.; Prokopidis, K.; Raleigh, S.M.; Kelaiditi, E.; Hill, M. Altered mitochondrial microenvironment at the spotlight of musculoskeletal aging and Alzheimer’s disease. Sci. Rep. 2022, 12, 11290. [Google Scholar] [PubMed]
- Yedigaryan, L.; Gatti, M.; Marini, V.; Maraldi, T.; Sampaolesi, M. Shared and divergent epigenetic mechanisms in cachexia and sarcopenia. Cells 2022, 11, 2293. [Google Scholar] [CrossRef]
- Antuña, E.; Cachán-Vega, C.; Bermejo-Millo, J.C.; Potes, Y.; Caballero, B.; Vega-Naredo, I.; Coto-Montes, A.; Garcia-Gonzalez, C. Inflammaging: Implications in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 15039. [Google Scholar] [CrossRef]
Figure 1.
Chromosomal locations and approximate sizes of the genes review in this article. Horizontal bars show intronic regions and the yellow boxes represent exons which are numbered according to their position. Relevant single nucleotide polymorphisms (SNPs) and the ACE indel are also shown along with their relative positions. Gene structure and information on each SNP were obtained by reference to the University of California Santa Cruz Genome Browser (hg38) at: https://genome.ucsc.edu/ (accessed on 24 April 2024).
Figure 1.
Chromosomal locations and approximate sizes of the genes review in this article. Horizontal bars show intronic regions and the yellow boxes represent exons which are numbered according to their position. Relevant single nucleotide polymorphisms (SNPs) and the ACE indel are also shown along with their relative positions. Gene structure and information on each SNP were obtained by reference to the University of California Santa Cruz Genome Browser (hg38) at: https://genome.ucsc.edu/ (accessed on 24 April 2024).


{kind=link}
Table 1.
Summary of the role of risk genes, non-coding RNAs, and microRNAs discussed in this review with relevance to both sarcopenia and Alzheimer’s disease.
Table 1.
Summary of the role of risk genes, non-coding RNAs, and microRNAs discussed in this review with relevance to both sarcopenia and Alzheimer’s disease.
| Risk Genes, Non-Coding RNAs, and MicroRNAs | Potential Epigenetic Regulation | Sarcopenia | Alzheimer’s Disease |
|---|---|---|---|
| APOE ε4 variant | DNA methylation | Inflammation is a risk factor for sarcopenia, and the APOE ε4 variant is associated with low-grade inflammation [17]. | APOE ε4 variant is a strong risk factor, associated with amyloid deposition increase and an earlier stage of onset [17]. |
| BDNF | DNA methylation | Reduced plasma BDNF associated with sarcopenia and dementia in adults aged 70–84 [34]. Lower levels of BDNF reported in patients with Parkinson’s [35]. | Serum BDNF lower in patients with Alzheimer’s compared to healthy controls [37]. BDNF lower in peripheral blood samples of patients with the lowest MMSE sores but higher in early Alzheimer’s disease [38]. In patients with mild cognitive impairment (MCI) that progressed to Alzheimer’s disease; elevated methylation of a CpG site within the promoter of BDNF was significantly associated with the conversion of MCI to Alzheimer’s [41]. |
| ACE I/D variant | DNA methylation Histone Modifications | In a Brazilian population, the II genotype of the ACE I/D variant predisposes to sarcopenia [44]. In an elderly Indonesian population, the DD genotype was found to be associated with sarcopenia [45]. | Meta-analysis suggests that the I allele of the ACE I/D variant increased the risk of Alzheimer’s Disease, which is supported by Lehmann and colleagues [47,48]. |
| Methylation of ACE has an indirect effect on the stress response and predisposition to depression, as it can modulate the levels of cortisol, and elevated levels of cortisol have been associated with sarcopenia and Alzheimer’s Disease [53,54]. | |||
| FTO | DNA methylation | GWAS identified 29 SNPs in a Caucasian cohort that were associated with sarcopenia [61]. GWAS followed by FTO knockdown in mice produced a sarcopenic profile [63]. The FTO rs9939609 variant is associated with sarcopenia when defining sarcopenia using percentage of muscle mass and skeletal muscle mass index but not when using the EWGSOP definition of sarcopenia [64]. | The rs649960 SNP within intron 1 of FTO significantly associates with Alzheimer’s in a large Caucasian cohort, and rs10852521, rs16945088, and rs8044769 were close to significance [65]. In a Hispanic population, rs17219084, rs11075996, and rs11075997 are significantly associated with Alzheimer’s disease [65]. Significant association found between the rs9939609 variant of FTO (AA genotype) and Alzheimer’s disease and dementia [66]. |
| FNDC5 (Irisin) | DNA methylation | Circulating Irisin levels are lower in patients with sarcopenia [73,74]. The rs16835198 and rs726344 variants of FNDC5 can impact insulin sensitivity, which is a contributing factor to the development of sarcopenia [82,83]. | The rs1746661 variant of FNDC5 influences glucose metabolism in elderly individuals which affects the risk of Alzheimer’s disease [79]. FNDC5 knockout in a mice model reduces the benefits of exercise-induced cognitive enhancement, which supports the role of Irisin in Alzheimer’s disease protection [80]. |
| hsa-miR-221-3p | Non-coding RNA | Downregulated in post-menopausal women with muscle wasting [86]. | N/A |
| hsa-miR-374b-5p | Non-coding RNA | N/A | |
| hsa-miR-146a-5p | Non-coding RNA | N/A | |
| hsa-miR-126-5p | Non-coding RNA | Altered levels in neutrophil-derived microvesicles of patients with Alzheimer’s disease [90]. | |
| hsa-miR-425-5p | Non-coding RNA | Upregulation promotes tau phosphorylation, which is a hallmark of Alzheimer’s disease pathology [92], and apoptosis in Alzheimer’s disease [91]. Upregulation also shown to be a predictor of both Alzheimer’s disease and mild cognitive impairment [93]. | |
| hsa-miR-145-5p | Non-coding RNA | Upregulated in post-menopausal women with muscle wasting [86]. | N/A |
| hsa-miR-25-3p | Non-coding RNA | N/A | |
| miR-133a | Non-coding RNA | Downregulated in patients with sarcopenia [87]. | N/A |
| miR-133b | Non-coding RNA | N/A | |
| miR-206 | Non-coding RNA | N/A | |
| miR-208b | Non-coding RNA | N/A | |
| miR-499 | Non-coding RNA | N/A | |
| miR-155-5p | Non-coding RNA | Repressed in sarcopenia, but expression levels return to normal upon exercise intervention [88]. | May play a role in neuron projection development and neuron morphogenesis in Alzheimer’s disease and could also act as a clinical biomarker for Alzheimer’s disease [96]. |
| miR-421-3p | Non-coding RNA | N/A | |
| miR-495-3p | Non-coding RNA | N/A | |
| miR-495-5p | Non-coding RNA | N/A | |
| miR-1-3p | Non-coding RNA | Elevated expression in patients with sarcopenia and heart disease may be beneficial to use as a biomarker to predict sarcopenia in patients with heart disease [89]. | N/A |
| miRNA-1 | Non-coding RNA | Increased expression in patients aged 55 to 80 with poor muscle performance [87]. | N/A |
| miRNA-29a | Non-coding RNA | Upregulated levels tentatively associated with Alzheimer’s disease [94]. | |
| miR-29b | Non-coding RNA | Downregulated in Egyptian patients with Alzheimer’s disease [95]. | |
| miRNA-206 | Non-coding RNA | Decreased expression in patients aged 55 to 80 with poor muscle performance [87]. | N/A |
| miRNA-133a | Non-coding RNA | N/A | |
| miRNA-133b | Non-coding RNA | N/A | |
| miRNA-208b | Non-coding RNA | N/A | |
| miRNA-499 | Non-coding RNA | N/A | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Raleigh, S.M.; Orchard, K.J.A. Sarcopenia as a Risk Factor for Alzheimer’s Disease: Genetic and Epigenetic Perspectives. Genes 2024, 15, 561. https://0-doi-org.brum.beds.ac.uk/10.3390/genes15050561
AMA Style
Raleigh SM, Orchard KJA. Sarcopenia as a Risk Factor for Alzheimer’s Disease: Genetic and Epigenetic Perspectives. Genes. 2024; 15(5):561. https://0-doi-org.brum.beds.ac.uk/10.3390/genes15050561
Chicago/Turabian StyleRaleigh, Stuart M., and Kayleigh J. A. Orchard. 2024. "Sarcopenia as a Risk Factor for Alzheimer’s Disease: Genetic and Epigenetic Perspectives" Genes 15, no. 5: 561. https://0-doi-org.brum.beds.ac.uk/10.3390/genes15050561
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.