
Functional Characterization of the MeSSIII-1 Gene and Its Promoter from Cassava
 ,
,
Abstract
:1. Introduction
2. Results
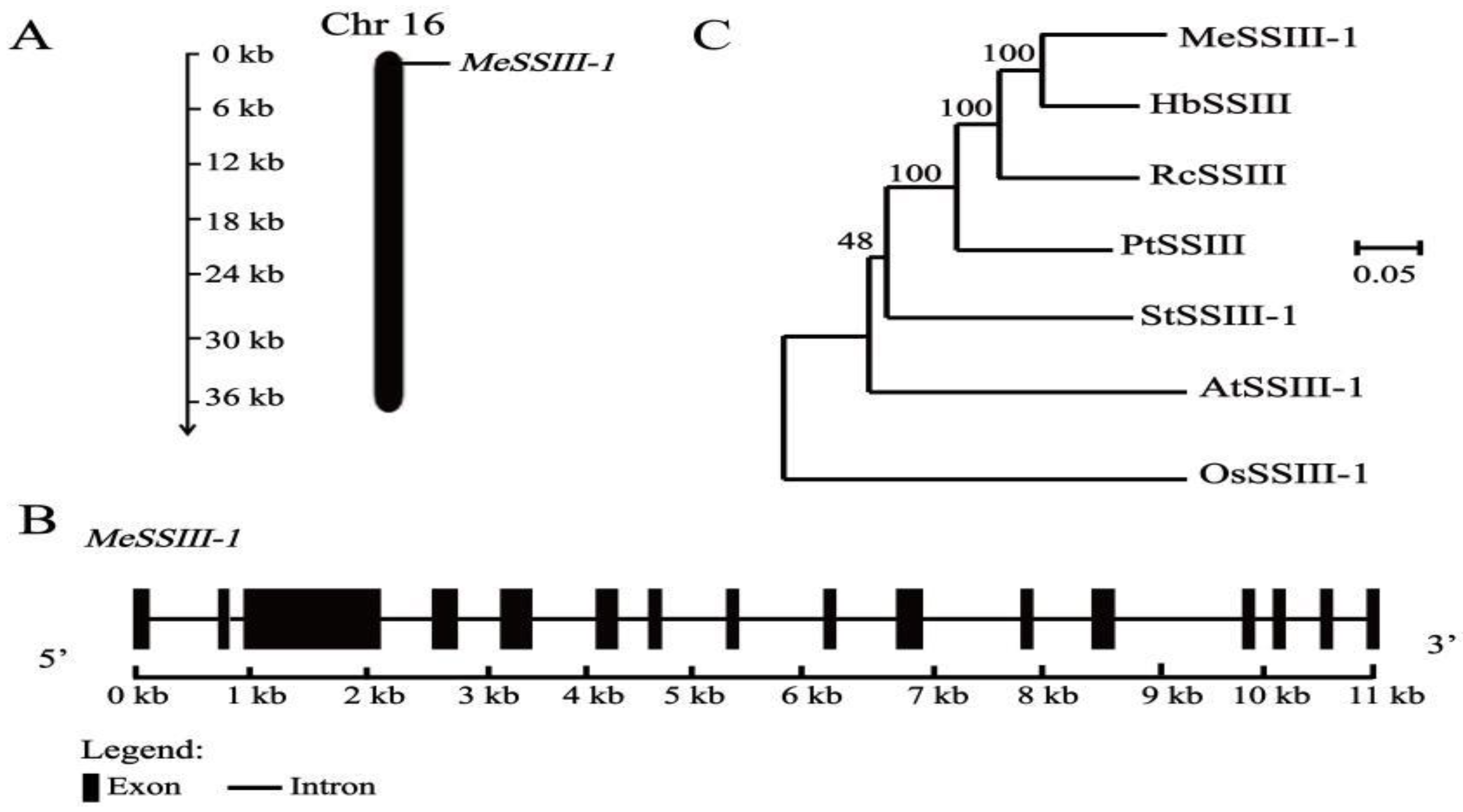
2.1. Isolation and Characterization of MeSSIII-1
2.2. Subcellular Localization of MeSSIII-1 Protein
2.3. Expression Patterns of MeSSIII-1 in Cassava Tissues
2.4. Isolation and Cis-Element Analysis of MeSSIII-1 Promoter
2.5. Expression Profiles of MeSSIII-1 Gene under Hormone Treatments
2.6. Analysis of MeSSIII-1 Promoter Activity
2.7. Histochemical Localization of proMeSSIII-1 in Cassava
2.8. Activities of MeSSIII-1 Promoter in Response to MeJA, ABA, and ET Hormones
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Bioinformatics Analysis of the MeSSIII-1 Gene and Promoter
4.3. Cloning of MeSSIII-1 Gene and Promoter of Cassava
4.4. Subcellular Localization of MeSSIII-1 Protein
4.5. Expression Pattern of MeSSIII-1 Gene
4.6. Construction of Truncated MeSSIII-1 Promoter Vectors and Tobacco Leaves Injection
4.7. GUS Staining and Determination of GUS Activity
4.8. Genetic Transformation and Identification of proMeSSIII-1 Promoter in Cassava
4.9. Hormone Treatments in proMeSSIII-1 Transgenic Cassava
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palavecino, C.E.; Carrasco-Véliz, N.; Quest, A.F.; Garrido, M.P.; Valenzuela-Valderrama, M. The 5′ untranslated region of the anti-apoptotic protein survivin contains an inhibitory upstream AUG codon. Biochem. Biophys. Res. Commun. 2020, 526, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.; Wang, W.Q.; Jiang, X.N.; Chen, X.M. Review on plant gene promoters. Acta Genet. Sin. 2004, 31, 1455–1464. [Google Scholar] [PubMed]
- Misra, S.; Ganesan, M. The impact of inducible promoters in transgenic plant production and crop improvement. Plant Gene 2021, 27, 100300. [Google Scholar] [CrossRef]
- Matsui, T.; Matsuura, H.; Sawada, K.; Takita, E.; Kinjo, S.; Takenami, S.; Ueda, K.; Nishigaki, N.; Yamasaki, S.; Hata, K. High level expression of transgenes by use of 5′-untranslated region of the Arabidopsis thaliana arabinogalactan-protein 21 gene in dicotyledons. Plant Biotechnol. 2012, 29, 319–322. [Google Scholar] [CrossRef]
- Yang, R.; Bai, J.; Li, R.; Chang, L. Research progress of the inducible promoters in plant genetic engineering. J. Shanxi Agric. Sci. 2018, 46, 292–298. [Google Scholar]
- Huang, L.C.; Li, Q.F.; Zhang, C.Q.; Chu, R.; Gu, Z.W.; Tan, H.Y.; Zhao, D.S.; Fan, X.L.; Liu, Q.Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.C.; Liu, T.L.; Ma, X.L.; Wang, B.; Zheng, Z.Y.; Zhang, Y.L.; Xie, X.R.; Yang, B.W.; Zhao, Z.; Zhu, Q.L.; et al. Quantitative regulation of Waxy expression by CRISPR/Cas9-based promoter and 5′UTR-intron editing improves grain quality in rice. Plant Biotechnol. J. 2020, 18, 2385. [Google Scholar] [CrossRef]
- Tetlow, I.J.; Morell, M.K.; Emes, M.J. Recent developments in understanding the regulation of starch metabolism in higher plants. J. Exp. Bot. 2004, 55, 2131–2145. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef]
- D’Hulst, C.; Wattebled, F.; Szydlowski, N. Starch biosynthesis in leaves and its regulation. In Starch: Metabolism and Structure; Nakamura, Y., Ed.; Springer: Tokyo, Japen, 2015; pp. 211–237. [Google Scholar]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Tuncel, A.; Okita, T.W. Improving starch yield in cereals by over-expression of ADPglucose pyrophosphorylase: Expectations and unanticipated outcomes. Plant Sci. 2013, 211, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef]
- Huang, L.C.; Tan, H.Y.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.B.; Kang, H.Q.; Wu, H.B.; Lian, X.J.; Wang, J.Y.; Zou, M.Z.; Li, Y.P. Analysis of visible and IR absorbance of maize, sweet potato and potato amylose before and after retrogradation. Food Sci. Technol. 2014, 39, 264–267. [Google Scholar]
- Zhong, Y.Y.; Liang, W.X.; Pu, H.Q.; Blennow, A.; Liu, X.X.; Guo, D.W. Short-time microwave treatment affects the multi-scale structure and digestive properties of high-amylose maize starch. Int. J. Biol. Macromol. 2019, 137, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.Y.; Liu, L.S.; Qu, J.; Blennow, A.; Hansen, A.R.; Wu, Y.X.; Guo, D.W.; Liu, X.X. Amylose content and specific fine structures affect lamellar structure and digestibility of maize starches. Food Hydrocoll. 2020, 108, 105994. [Google Scholar] [CrossRef]
- Blennow, A. Chapter 4-Starch Bioengineering. In Starch in Food, 2nd ed.; Sjöö, M., Nilsson, L., Eds.; Woodhead Publishing: Sawston, UK, 2018; pp. 179–222. [Google Scholar]
- Guan, S.; Wang, P.; Liu, H.; Liu, G.; Ma, Y.; Zhao, L. Production of high-amylose maize lines using RNA interference in sbe2a. Afr. J. Biotechnol. 2011, 10, 15229–15237. [Google Scholar] [CrossRef]
- Yangcheng, H.Y.; Jiang, H.X.; Blanco, M.; Jane, J.L. Characterization of normal and waxy corn starch for bioethanol production. J. Agric. Food Chem. 2013, 61, 379–386. [Google Scholar] [CrossRef]
- Tappiban, P.; Duncan, R.; Triwitayakorn, K.; Bao, J. Recent understanding of starch biosynthesis in cassava for quality improvement: A review. Trends Food Sci. Technol. 2019, 83, 167–180. [Google Scholar] [CrossRef]
- Zhao, S.S.; Dufour, D.; Sánchez, T.; Ceballos, H.; Zhang, P. Development of waxy cassava with different Biological and physico-chemical characteristics of starches for industrial applications. Biotechnol. Bioeng. 2011, 108, 1925–1935. [Google Scholar] [CrossRef]
- Bull, S.E.; Seung, D.; Chanez, C.; Mehta, D.; Kuon, J.-E.; Truernit, E.; Hochmuth, A.; Zurkirchen, I.; Zeeman, S.C.; Gruissem, W.; et al. Accelerated ex situ breeding of GBSS-and PTST1-edited cassava for modified starch. Sci. Adv. 2018, 4, eaat6086. [Google Scholar] [CrossRef]
- He, S.T.; Hao, X.M.; Wang, S.S.; Zhou, W.Z.; Ma, Q.X.; Lu, X.L.; Chen, L.N.; Zhang, P. Starch synthase II plays a crucial role in starch biosynthesis and the formation of multienzyme complexes in cassava storage roots. J. Exp. Bot. 2022, 73, 2540–2557. [Google Scholar] [CrossRef]
- Ying, Y.; Hu, Y.; Zhang, Y.; Tappiban, P.; Zhang, Z.; Dai, G.; Deng, G.; Bao, J.; Xu, F. Identification of a new allele of soluble starch synthase IIIa involved in the elongation of amylopectin long chains in a chalky rice mutant. Plant Sci. Int. J. Exp. Plant Biol. 2023, 328, 111567. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Fang, K.X.; Chen, X.; Ma, P.A.; Zhou, X.C.; Lu, C.; Wang, W.Q. Change of endogenous hormones and expression levels of genes for ABA synthesis, signaling and starch synthesis in leaves of cassava under exogenous ABA treatment. J. China Agric. Univ. 2015, 20, 100–107. [Google Scholar]
- Luo, X.L.; Pan, X.L.; Zhu, Y.M. The relationship between endogenous abscisic acid content and starch accumulation in root tuber of cassava. Chin. J. Trop. Crops 2018, 39, 472–479. [Google Scholar]
- Huang, B.; Keeling, P.L.; Hennen-Bierwagen, T.A.; Myers, A.M. Comparative in vitro analyses of recombinant maize starch synthases SSI, SSIIa, and SSIII reveal direct regulatory interactions and thermosensitivity. Arch. Biochem. Biophys. 2016, 596, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Myers, A.M.; James, M.G. Mutations affecting starch synthaseIII in arabidopsis alter leaf starch structure and increase the rate of starch synthesis. Plant Physiol. 2005, 138, 663–674. [Google Scholar] [CrossRef]
- Ohdan, T.; Francisco, P.B.; Sawada, T.; Hirose, T.; Terao, T.; Satoh, H.; Nakamura, Y. Expression profiling of genes involved in starch synthesis in sink and source organs of rice. J. Exp. Bot. 2005, 56, 3229–3244. [Google Scholar] [CrossRef] [PubMed]
- Fujita, N.; Yoshida, M.; Kondo, T.; Saito, K.; Utsumi, Y.; Tokunaga, T.; Nishi, A.; Satoh, H.; Park, J.H.; Jane, J.L.; et al. Characterization of SSIIIa-deficient mutants of rice: The function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm. Plant Physiol. 2007, 144, 2009–2023. [Google Scholar] [CrossRef]
- Wang, A.Q.; Jing, Y.H.; Cheng, Q.; Zhou, H.J.; Wang, L.J.; Gong, W.X.; Kou, L.Q.; Liu, G.F.; Meng, X.B.; Chen, M.J.; et al. Loss of function of SSIIIa and SSIIIb coordinately confers high RS content in cooked rice. Proc. Natl. Acad. Sci. USA 2023, 120, e2220622120. [Google Scholar] [CrossRef]
- Yan, W.; Wu, X.; Li, Y.; Liu, G.; Cui, Z.; Jiang, T.; Ma, Q.; Luo, L.; Zhang, P. Cell Wall Invertase 3 affects cassava productivity via regulating sugar allocation from source to sink. Front. Plant Sci. 2019, 10, 541. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Zhang, H.; Zhai, H.; Liu, Q.; He, S. A soluble starch synthase I gene, IbSSI, alters the content, composition, granule size and structure of starch in transgenic sweet potato. Sci. Rep. 2017, 7, 2315. [Google Scholar] [CrossRef] [PubMed]
- Szydlowski, N.; Ragel, P.; Raynaud, S.; Lucas, M.; Roldán, I.; Montero, M.; Muñoz, F.J.; Ovecka, M.; Bahaji, A.; Planchot, V.; et al. Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthases. Plant Cell 2009, 21, 2443–2457. [Google Scholar] [CrossRef] [PubMed]
- Gámez-Arjona, F.M.; Raynaud, S.; Ragel, P.; Mérida, Á. Starch synthase 4 is located in the thylakoid membrane and interacts with plastoglobule-associated proteins in Arabidopsis. Plant J. 2014, 80, 305–316. [Google Scholar] [CrossRef]
- Zhou, W.Z.; He, S.T.; Naconsie, M.; Ma, Q.X.; Zeeman, S.C.; Gruissem, W.; Zhang, P. Alpha-glucan, water dikinase 1 affects starch metabolism and storage root growth in cassava (Manihot esculenta Crantz). Sci. Rep. 2017, 7, 9863. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting transgene expression in research, agricultural, and environmentalapplications: Promoters used in plant transformation. In Vitro Cell. Dev. Biol. Plant 2004, 40, 1–22. [Google Scholar] [CrossRef]
- Shibata, M.; Gulden, F.O.; Sestan, N. From trans to cis: Transcriptional regulatory networks in neocortical development. Trends Genet. 2015, 31, 77–87. [Google Scholar] [CrossRef]
- Zou, C.; Sun, K.; Mackaluso, J.D.; Seddon, A.E.; Jin, R.; Thomashow, M.F.; Shiu, S.-H. Cis-regulatory code of stress-responsive transcription in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 14992–14997. [Google Scholar] [CrossRef]
- Dao, L.T.; Spicuglia, S. Transcriptional regulation by promoters with enhancer function. Transcription 2018, 9, 307–314. [Google Scholar] [CrossRef]
- Hu, Y.F.; Li, Y.P.; Zhang, J.J.; Liu, H.M.; Chen, Z.Y.; Huang, Y.B. PzsS3a, a novel endosperm specific promoter from maize (Zea mays L.) induced by ABA. Biotechnol. Lett. 2011, 33, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Xie, S.D.; Xiao, Q.L.; Wei, B.; Zheng, L.J.; Wang, Y.B.; Cao, Y.; Zhang, X.G.; Long, T.D.; Li, Y.P.; et al. Sucrose and ABA regulate starch biosynthesis in maize through a novel transcription factor, ZmEREB156. Sci. Rep. 2016, 6, 27590. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.B.; Torbjörn, L.; Örberg, H.; Wei, M.G.; Hedman, B.; Ren, J.W.; Xie, G.H.; Xiong, S.J. Cassava stems: A new resource to increase food and fuel production. GCB Bioenergy 2013, 7, 72–83. [Google Scholar] [CrossRef]
- Guan, Z.H.; Chen, X.; Xie, H.R.; Wang, W.Q. Promoter regulatory domain identification of cassava starch synthase IIb gene in transgenic tobacco. Plant Physiol. Biochem. 2016, 102, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.L. Studies on the effects of different plant growth regulators on the growth and starch accumulation of cassava. Chin. Agric. Sci. Bull. 2002, 3, 30–33. [Google Scholar]
- Takahashi, I.; Hara, M. Enhancement of starch accumulation in plants by exogenously applied methyl jasmonate. Plant Biotechnol. Rep. 2014, 8, 143–149. [Google Scholar] [CrossRef]
- Xing, C.; Liu, Y.; Zhao, L.; Zhang, S.; Huang, X. A novel MYB transcription factor regulates ascorbic acid synthesis and affects cold tolerance. Plant Cell Environ. 2019, 42, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, L.; Huang, Y.; Wu, H.; Zhang, X.; Hu, X.; Ma, Q. Identification and characterization of the core region of ZmDi19-5 promoter activity and its upstream regulatory proteins. Int. J. Mol. Sci. 2022, 23, 7390. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Lu, X.H.; Zhen, X.H.; Yang, H.; Che, Y.N.; Hou, J.Y.; Geng, M.T.; Liu, J.; Hu, X.W.; Li, R.M.; et al. A Transformation and genome editing system for cassava cultivar SC8. Genes 2022, 13, 1650. [Google Scholar] [CrossRef]
- Yan, Y.; Zhao, S.H.; Ye, X.X.; Tian, L.B.; Shang, S.; Tie, W.W.; Zeng, L.W.; Zeng, L.M.; Yang, J.H.; Li, M.Y.; et al. Abscisic acid signaling in the regulation of postharvest physiological deterioration of sliced cassava tuberous roots. J. Agric. Food Chem. 2022, 70, 12830–12840. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cis-Elements | Number | Function | Core Sequence |
|---|---|---|---|
| ABRE | 1 | cis-acting element involved in the abscisic acid responsiveness | ACGTG |
| ERE | 6 | ethylene-responsive element | ATTT TAAA |
| CGTCA-motif | 2 | cis-acting regulatory element involved in the MeJA-responsiveness | CGTCA |
| TGACG-motif | 2 | cis-acting regulatory element involved in the MeJA-responsiveness | TGACG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.-H.; Wang, Y.-J.; Zhen, X.-H.; Yu, H.; Pan, M.; Fu, D.-Q.; Li, R.-M.; Liu, J.; Luo, H.-Y.; Hu, X.-W.; et al. Functional Characterization of the MeSSIII-1 Gene and Its Promoter from Cassava. Int. J. Mol. Sci. 2024, 25, 4711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094711
Lu X-H, Wang Y-J, Zhen X-H, Yu H, Pan M, Fu D-Q, Li R-M, Liu J, Luo H-Y, Hu X-W, et al. Functional Characterization of the MeSSIII-1 Gene and Its Promoter from Cassava. International Journal of Molecular Sciences. 2024; 25(9):4711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094711
Chicago/Turabian StyleLu, Xiao-Hua, Ya-Jie Wang, Xing-Hou Zhen, Hui Yu, Mu Pan, Dong-Qing Fu, Rui-Mei Li, Jiao Liu, Hai-Yan Luo, Xin-Wen Hu, and et al. 2024. "Functional Characterization of the MeSSIII-1 Gene and Its Promoter from Cassava" International Journal of Molecular Sciences 25, no. 9: 4711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094711