
Altered Functional Connectivity during Mild Transient Respiratory Impairment Induced by a Resistive Load
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Induction and Measurement of Respiratory Impairment
2.3. Experimental Design
2.4. MRI Data Acquisition
2.5. Data Pre-Processing
2.6. ROI-to-ROI Analysis
2.7. Seed-to-Voxel Analysis
2.8. Graph Theoretical Analysis
2.9. Correlation with the Modified Borg Scale
3. Results
3.1. Participant Selection
3.2. Changes in the Modified Borg Scale and Vital Signs during Respiratory Training between Normal Breathing and Effortful Breathing
3.3. Adverse Events
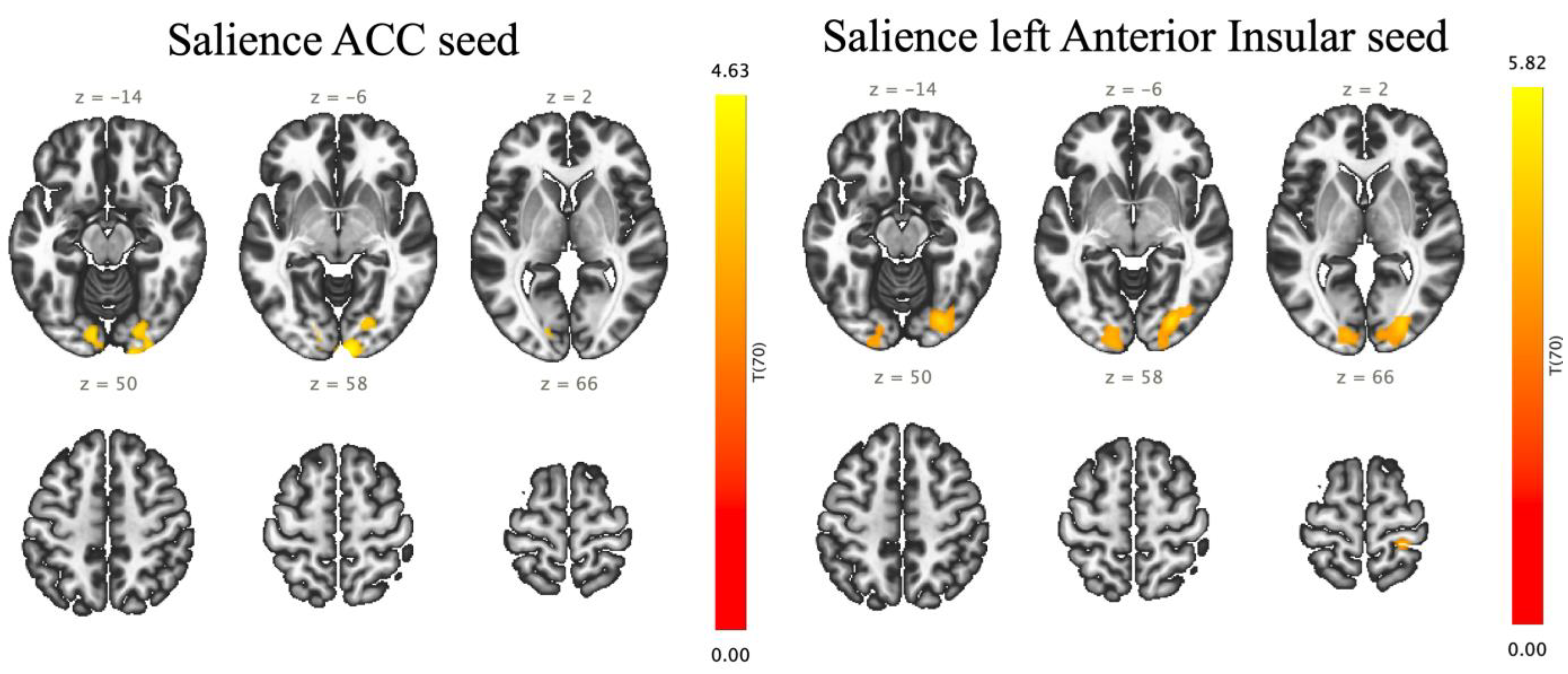
3.4. ROI-to-ROI Results
3.5. Seed-to-Voxel Results
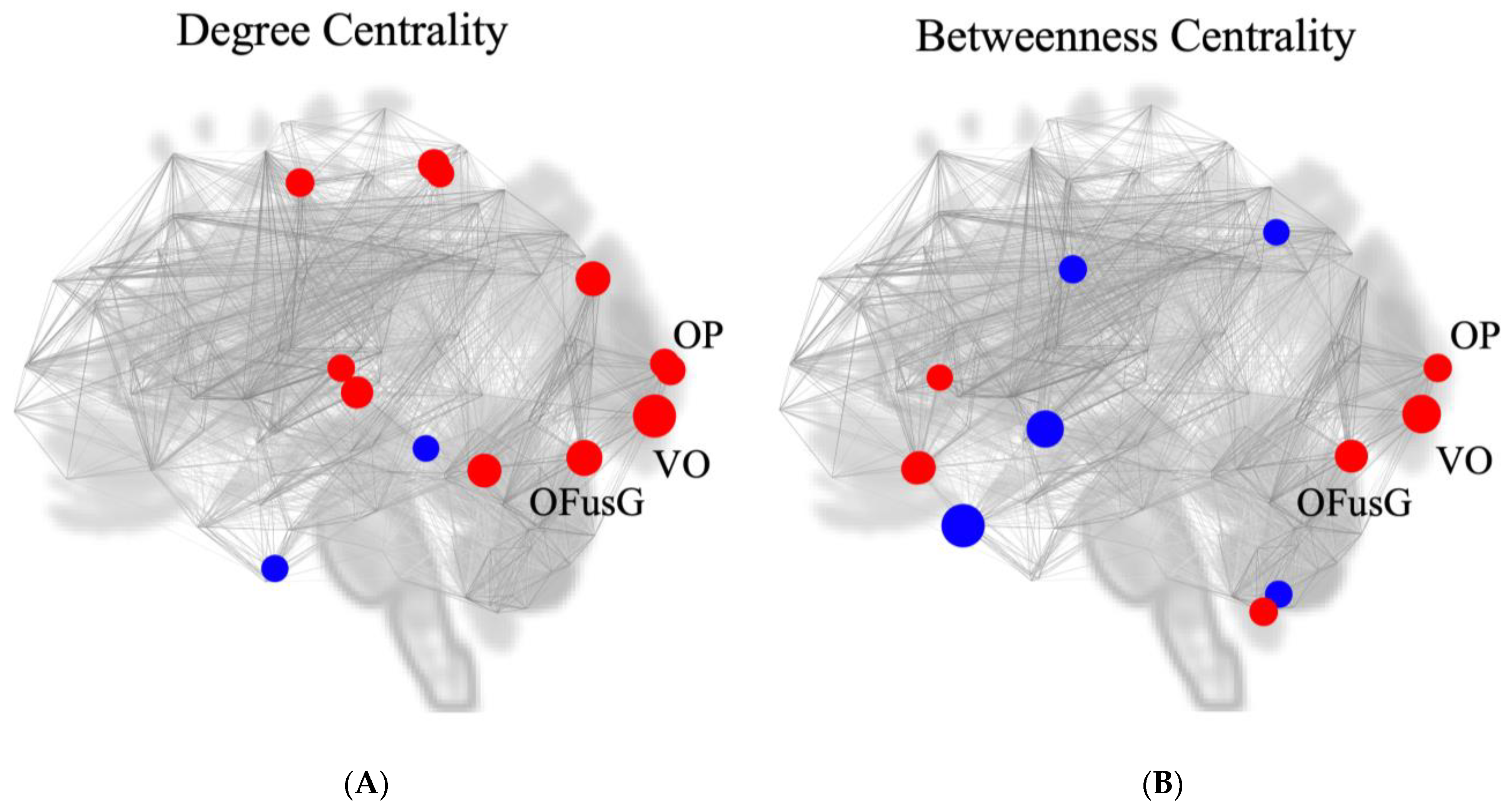
3.6. Measurement of Nodal Centrality
3.7. Correlation with the Modified Borg Scale
4. Discussion
4.1. Unique Point of Our Resistive Load
4.2. Functional Connectivity within the Respiratory-Associated Area
4.3. Functional Connectivity Outside of the Respiratory-Associated Area
4.4. Correlation with Clinical Score (Modified Borg Scale)
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, J.L.; Del Negro, C.A. Looking for inspiration: New perspectives on respiratory rhythm. Nat. Rev. Neurosci. 2006, 7, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.C. Cortico-limbic circuitry and the airways: Insights from functional neuroimaging of respiratory afferents and efferents. Biol. Psychol. 2010, 84, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Gore, J.C. Principles and practice of functional MRI of the human brain. J. Clin. Investig. 2003, 112, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Rosazza, C.; Minati, L. Resting-state brain networks: Literature review and clinical applications. Neurol. Sci. 2011, 32, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Srivastava, G.; Reiss, A.L.; Menon, V. Default mode network activity distinguishes Alzheimer’s disease from healthy aging: Evidence from functional MRI. Proc. Natl. Acad. Sci. USA 2004, 101, 4637–4642. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Flores, B.H.; Menon, V.; Glover, G.H.; Solvason, H.B.; Kenna, H.; Reiss, A.L.; Schatzberg, A.F. Resting-state functional connectivity in major depression: Abnormally increased contributions from subgenual cingulate cortex and thalamus. Biol. Psychiatry 2007, 62, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Garrity, A.G.; Pearlson, G.D.; McKiernan, K.; Lloyd, D.; Kiehl, K.A.; Calhoun, V.D. Aberrant “default mode” functional connectivity in schizophrenia. Am. J. Psychiatry 2007, 164, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, D.; Qin, W.; Li, Q.; Chen, B.; Zhang, Y.; Yu, C. Altered resting-state brain activity in obstructive sleep apnea. Sleep 2013, 36, 651–659B. [Google Scholar] [CrossRef]
- Li, H.; Xin, H.; Yu, J.; Yu, H.; Zhang, J.; Wang, W.; Peng, D. Abnormal intrinsic functional hubs and connectivity in stable patients with COPD: A resting-state MRI study. Brain Imaging Behav. 2020, 14, 573–585. [Google Scholar] [CrossRef]
- Walter, H.; Kausch, A.; Dorfschmidt, L.; Waller, L.; Chinichian, N.; Veer, I.; Hilbert, K.; Lüken, U.; Paulus, M.P.; Goschke, T.; et al. Self-control and interoception: Linking the neural substrates of craving regulation and the prediction of aversive interoceptive states induced by inspiratory breathing restriction. Neuroimage 2020, 215, 116841. [Google Scholar] [CrossRef]
- Barkhof, F.; Haller, S.; Rombouts, S.A.R.B. Resting-state functional MR imaging: A new window to the brain. Radiology 2014, 272, 29–49. [Google Scholar] [CrossRef] [PubMed]
- Macefield, V.G.; Gandevia, S.C.; Henderson, L.A. Neural sites involved in the sustained increase in muscle sympathetic nerve activity induced by inspiratory capacity apnea: A fMRI study. J. Appl. Physiol. 2006, 100, 266–273. [Google Scholar] [CrossRef]
- McKay, L.C.; Adams, L.; Frackowiak, R.S.; Corfield, D.R. A bilateral corticobulbar network associated with breath holding in humans, determined by functional magnetic resonance imaging. NeuroImage 2008, 40, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- Pattinson, K.T.S.; Mitsis, G.D.; Harvey, A.K.; Jbabdi, S.; Dirckx, S.; Mayhew, S.D.; Rogers, R.; Tracey, I.; Wise, R.G. Determination of the human brainstem respiratory control network and its cortical connections in vivo using functional and structural imaging. NeuroImage 2009, 44, 295–305. [Google Scholar] [CrossRef]
- Yorita, A.; Tokunaga, Y.; Kinoshita, T.; Nakakura, A.; Oda, H.; Imaoka, H.; Matsunaga, K.; Kakuma, T.; Hoshino, T.; Kawayama, T. Usefulness of a 4-grade novel mouthpiece device for increased mouth pressure reproducing artificial difficulty in breathing. Kurume Med. J. 2023, 68, 229–238. [Google Scholar] [CrossRef]
- Fink, G.R.; Corfield, D.R.; Murphy, K.; Kobayashi, I.; Dettmers, C.; Adams, L.; Frackowiak, R.S.; Guz, A. Human cerebral activity with increasing inspiratory force: A study using positron emission tomography. J. Appl. Physiol. 1996, 81, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, C.; Poline, J.-B.; Thivard, L.; Aubier, M.; Samson, Y. Neural substrates for the perception of acutely induced dyspnea. Am. J. Respr. Crit. Care Med. 2001, 163, 951–957. [Google Scholar] [CrossRef]
- Ramsay, S.C.; Adams, L.; Murphy, K.; Corfield, D.R.; Grootoonk, S.; Bailey, D.L.; Frackowiak, R.S.J.; Guz, A. Regional cerebral blood flow during volitional expiration in man: A comparison with volitional inspiration. J. Physiol. 1993, 461, 85–101. [Google Scholar] [CrossRef]
- Stoeckel, M.C.; Esser, R.W.; Gamer, M.; Buchel, C.; von Leupolt, A. Brain responses during the anticipation of dyspnea. Neural Plast. 2016, 2016, 6434987. [Google Scholar] [CrossRef]
- von Leupoldt, A.; Sommer, T.; Kegat, S.; Baumann, H.J.; Klose, H.; Dahme, B.; Buchel, C. The unpleasantness of perceived dyspnea is processed in the anterior insula and amygdala. Am. J. Respir. Crit. Care Med. 2008, 177, 1026–1032. [Google Scholar] [CrossRef]
- von Leupoldt, A.; Sommer, T.; Kegat, S.; Baumann, H.J.; Klose, H.; Dahme, B.; Buchel, C. Dyspnea and pain share emotion-related brain network. NeuroImage 2009, 48, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Borg, G. Subjective effort and physical activities. Scand. J. Rehab. Med. 1978, 6, 105–113. [Google Scholar]
- Wilson, R.C.; Jones, P.W. A comparison of the visual analogue scale and modified Borg scale for the measurement of dyspnoea during exercise. Clin. Sci. 1989, 76, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.W.; Kim, K.W.; Lee, D.Y.; Lee, S.B.; Park, J.H.; Choi, E.A.; Choe, J.Y.; Do, Y.J.; Ryang, J.S.; Roh, H.A.; et al. A normative study of the Revised Hasegawa Dementia Scale: Comparison of demographic influences between the Revised Hasegawa Dementia Scale and the Mini-MentalStatus Examination. Dement. Geriatr. Cogn. Disord. 2007, 24, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Porcu, M.; Craboledda, D.; Garofalo, P.; Barberini, L.; Sanfilippo, R.; Zaccagna, F.; Wintermark, M.; Montisci, R.; Saba, L. Reorganization of brain networks following carotid endarterectomy: An exploratory study using resting state functional connectivity with a focus on the changes in default mode network connectivity. Eur. J. Radiol. 2019, 110, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Podgorski, P.; Waliszewska-Prosol, M.; Zimny, A.; Sasiadek, M.; Bladowska, J. Resting-State functional connectivity of the ageing female brain-differences between young and elderly female adults on multislice short TR rs-fMRI. Front. Neurol. 2021, 12, 645974. [Google Scholar] [CrossRef] [PubMed]
- Amir, J.; Nari, J.K.R.; Del Carpio-O’Donovan, R.; Ptito, A.; Chen, J.-K.; Chankowsky, J.; Tinawi, S.; Lunkova, E.; Saluja, R.S. Atypical resting state functional connectivity in mild traumatic brain injury. Brain Behav. 2021, 11, e2261. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Segonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Frazier, J.A.; Chiu, S.; Breeze, J.L.; Makris, N.; Lange, N.; Kennedy, D.N.; Herbert, M.R.; Bent, E.K.; Koneru, V.K.; Dieterich, M.E.; et al. Structural brain magnetic resonance imaging of limbic and thalamic volumes in pediatric bipolar disorder. Am. J. Psychiatry 2005, 162, 1256–1265. [Google Scholar] [CrossRef]
- Goldstein, J.M.; Seidman, L.J.; Makris, N.; Ahern, T.; O’Brien, L.M.; Caviness Jr, V.S.; Kennedy, D.N.; Faraone, S.V.; Tsuang, M.T. Hypothalamic abnormalities in schizophrenia: Sex effects and genetic vulnerability. Biol. Psychiatry 2007, 61, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Goldstein, J.M.; Kennedy, D.; Hodge, S.M.; Caviness, V.S.; Faraone, S.V.; Tsuang, M.T.; Seidman, L.J. Decreased volume of left and total anterior insular lobule in schizophrenia. Schizophr. Res. 2006, 83, 155–171. [Google Scholar]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.J.; Perlson, G.D.; Stevens, M.; Calhoun, V.D. A method for functional network connectivity among spatially independent resting-state components in schizophrenia. NeuroImage 2008, 39, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Sporns, O. Network hubs in the human brain. Trends Cogn. Sci. 2013, 17, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 54, 1059–1069. [Google Scholar] [CrossRef]
- Malagutti, N.; Di Laora, A.; Barbetta, C.; Groppo, E.; Tugnoli, V.; Sette, E.; Astolfi, L.; Beswick, W.; Borin, M.; Ciorba, A.; et al. Is peripheral oxygen saturation a reliable predictor of upper airways air-flow limitation? J. Emer. Med. 2018, 55, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Higashimoto, Y.; Sano, A.; Nishiyama, O.; Sano, H.; Iwanaga, T.; Haraguchi, R.; Chiba, Y.; Fukuda, K.; Tohda, Y. Prefrontal cortex activation is associated with dyspnea during methacholine bronchial provocation tests in patients with bronchial asthma. Allergol. Int. 2020, 69, 453–454. [Google Scholar] [CrossRef]
- Ogawa, S.; Lee, T.M.; Kay, A.R.; Tank, D.W. Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proc. Natl. Acad. Sci. USA 1990, 87, 9868–9872. [Google Scholar] [CrossRef]
- Baker, C.M.; Burks, J.D.; Briggs, R.G.; Stafford, J.; Conner, A.K.; Glenn, C.A.; Sali, G.; McCoy, T.M.; Battiste, J.D.; O’Donoghue, D.L.; et al. A connectomic atlas of the human cerebrum-Chapter 4: The medial frontal lobe, anterior cingulate gyrus, and orbitofrontal cortex. Oper. Neurosurg. 2018, 15 (Suppl. 1), S122–S174. [Google Scholar] [CrossRef]
- Sturm, V.E.; Sollberger, M.; Seeley, W.W.; Rankin, K.P.; Ascher, E.A.; Rosen, H.J.; Miller, B.L.; Levenson, R.W. Role of right pregenual anterior cingulate cortex in self-conscious emotional reactivity. Soc. Cogn. Affect. Neurosci. 2013, 8, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Price, D.D. Psychological and neural mechanisms of the affective dimension of pain. Science 2000, 288, 1769–1772. [Google Scholar] [CrossRef] [PubMed]
- Tataranni, P.A.; Gautier, J.F.; Chen, K.; Uecker, A.; Bandy, D.; Salbe, A.D.; Pratley, R.E.; Lawson, M.; Reiman, E.M.; Ravussin, E. Neuroanatomical correlates of hunger and satiation in humans using positron emission tomography. Proc. Natl. Acad. Sci. USA 1999, 96, 4569–4574. [Google Scholar] [CrossRef] [PubMed]
- Phan, K.L.; Wager, T.; Taylor, S.F.; Liberzon, I. Functional neuroanatomy of emotion: A meta-analysis of emotion activation studies in PET and fMRI. Neuroimage 2002, 16, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D. How do you feel–now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Singer, T.; Critchley, H.D.; Preuschoff, K. A common role of insula in feelings, empathy and uncertainty. Trends Cogn. Sci. 2009, 13, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.M.; Cohen, J.D.; Carter, C.S. Conflict monitoring and anterior cingulate cortex: An update. Trends Cogn. Sci. 2004, 8, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Gaytan, S.P.; Pasaro, R. Connections of the rostral ventral respiratory neuronal cell group: An anterograde and retrograde tracing study in the rat. Brain Res. Bull. 1998, 47, 625–642. [Google Scholar] [CrossRef]
- Corfield, D.R.; Fink, G.R.; Ramsay, S.C.; Murphy, K.; Harty, H.R.; Watson, J.D.G.; Adams, L.; Frackowiak, R.S.; Guz, A. Evidence for limbic system activation during CO2-stimulated breathing in man. J. Physiol. 1995, 488, 77–84. [Google Scholar] [CrossRef]
- Evans, K.C.; Banzett, R.B.; Adams, L.; McKay, L.; Frackowiak, R.S.; Corfield, D.R. BOLD fMRI identifies limbic, paralimbic, and cerebellar activation during air hunger. J. Neurophysiol. 2002, 88, 1500–1511. [Google Scholar] [CrossRef] [PubMed]
- Sepulcre, J.; Sabuncu, M.R.; Yeo, T.B.; Liu, H.; Johnson, K.A. Stepwise connectivity of the modal cortex reveals the multimodal organization of the human brain. J. Neurosci. 2012, 32, 10649–10661. [Google Scholar] [CrossRef] [PubMed]
- Leech, R.; Braga, R.; Sharp, D.J. Echoes of the brain within the posterior cingulate cortex. J. Neurosci. 2012, 32, 215–222. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.I.; Turk, E.; Manning, J.H.; Hect, J.; Hernandez-Andrate, E.; Hassan, S.S.; Romero, R.; van den Heuvel, M.P.; Thomason, M.E. Hubs in the human fetal brain network. Dev. Cogn. Neurosci. 2018, 30, 108–115. [Google Scholar] [CrossRef]
- Wang, W.; Wang, P.; Li, Q.; Peng, Z.; Wang, X.; Wang, G.; Hou, J.; Fan, L.; Liu, S. Alterations of grey matter volumes and network-level functions in patients with stable chronic obstructive pulmonary disease. Neurosci. Lett. 2020, 720, 134748. [Google Scholar] [CrossRef]
- Tang, F.; Li, L.; Peng, D.; Yu, J.; Xin, H.; Tang, X.; Li, K.; Zeng, Y.; Xie, W.; Li, H. Abnormal static and dynamic functional network connectivity in stable chronic obstructive pulmonary disease. Front. Aging Neurosci. 2022, 14, 1009232. [Google Scholar] [PubMed]
- Xin, H.; Li, H.; Yu, H.; Yu, J.; Zhang, J.; Wang, W.; Peng, D. Disrupted resting-state spontaneous neural activity in stable COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2019, 14, 499–508. [Google Scholar] [CrossRef]
- Kendrick, K.R.; Baxi, S.C.; Smith, R.M. Usefullness of the modified 0-10 Borg scale in assessing the degree of dyspnea in patients with COPD and asthoma. J. Emerg. Nurs. 2000, 26, 216–222. [Google Scholar] [CrossRef]
- Mancini, I.; Body, J.J. Assessment of dyspnea in advanced cancer patients. Support Care Cancer 1999, 7, 229–232. [Google Scholar] [CrossRef]
- Mador, M.J.; Rodis, A.; Magalang, U.J. Reproducibility of Borg scale measurements of dyspnea during exercise in patients with COPD. Chest 1995, 107, 1590–1597. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Participants, n = 36 |
|---|---|
| Age, years | 35.0 ± 7.2 |
| Gender, women/men, n | 18/18 |
| Body mass index, kg/m2 | 22.6 ± 2.8 |
| Respiratory rates, /min | 15.5 ± 3.3 |
| Heartbeats, /min | 71.0 ± 8.4 |
| Blood pressure, mmHg | |
| Systolic | 116.0 ± 12.7 |
| Diastolic | 68.5 ± 10.6 |
| Saturation of percutaneous oxygen, % | 97.9 ± 1.0 |
| Hasegawa’s Dementia scale, points | 29.9 ± 0.5 |
| Lung function tests | |
| FVC, L | 4.0 ± 0.9 |
| %FVC predicted, % | 104.3 ± 12.3 |
| FEV1, L | 3.4 ± 0.7 |
| %FEV1 predicted, % | 102.5 ± 11.3 |
| FEV1/FVC ratio | 0.86 ± 0.06 |
| Blood tests | |
| White blood cell count, ×102/µL | 5.7 ± 1.1 |
| Red blood cell count, ×104/µL | 475 ± 39 |
| Haemoglobin, g/dL | 14.3 ± 1.6 |
| Haemoglobin A1c (NGSP), % | 5.2 ± 0.6 |
| Serum tests | |
| Thyroid-stimulating hormone, µIU/mL | 1.8 ± 0.9 |
| Blood urea nitrogen, mg/dL | 11.9 ± 2.8 |
| Creatinine, mg/dL | 0.7 ± 0.1 |
| Sodium, mEq/L | 140.4 ± 1.6 |
| Chloride, mEq/L | 104.1 ± 1.9 |
| Potassium, mEq/L | 4.1 ± 0.3 |
| Aspartate aminotransferase, U/L | 22.7 ± 8.4 |
| Alanine aminotransferase, U/L | 24.7 ± 14.1 |
| Lactate dehydrogenase, U/L | 177.2 ± 32.1 |
| Gamma-glutamyl transpeptidase, U/L | 35.6 ± 45.6 |
| Vitamin B1, ng/mL | 33.9 ± 7.1 |
| Analysis Unit (ROI-to-ROI) | Statistics | p-Uncorrected | p-FDR | |
|---|---|---|---|---|
| Cluster 1/325 F (3, 68) = 9.18 | 0.000035 | 0.011489 | ||
| Sal AInsula l | VO | T (70) = 4.51 | 0.000025 | 0.004139 |
| Sal AInsula l | OP r | T (70) = 4.26 | 0.000063 | 0.004326 |
| Sal AInsula l | OFusG r | T (70) = 4.19 | 0.000080 | 0.004326 |
| Sal ACC | OP r | T (70) = 3.92 | 0.000206 | 0.019694 |
| Sal ACC | OFusG r | T (70) = 3.78 | 0.000328 | 0.019694 |
| Sal ACC | VO | T (70) = 3.75 | 0.000362 | 0.019608 |
| Sal AInsula l | OP l | T (70) = 3.52 | 0.000752 | 0.028116 |
| Sal AInsula l | OFusG l | T (70) = 3.48 | 0.000862 | 0.028116 |
| Seed | Cluster Name | Clusters (x, y, z) | Size | Coverage | Size p-FEW | Size p-FDR | Size p-unc | Peak p-FEW | Peak p-unc |
|---|---|---|---|---|---|---|---|---|---|
| Sal. ACC | A | +06 −98 −04 | 858 | 303 voxels (35%) covering 12% of OP r | 0.000011 | 0.000018 | 0.000001 | 0.386005 | 0.000017 |
| 194 voxels (23%) covering 22% of OFusG r | |||||||||
| 76 voxels (9%) covering 3% of OP l | |||||||||
| 52 voxels (6%) covering 3% of LG l | |||||||||
| 31 voxels (4%) covering 3% of OFusG l | |||||||||
| 23 voxels (3%) covering 1% of LG r | |||||||||
| 18 voxels (2%) covering 1% of iLOC r | |||||||||
| 5 voxels (1%) covering 1% of ICC l | |||||||||
| 2 voxels (0%) covering 0% of Cereb 6 l | |||||||||
| 154 voxels (18%) covering 0% of not-labelled | |||||||||
| Sal. AInsular l | A | +26 −82 −04 | 1145 | 349 voxels (30%) covering 14% of OP r | 0.000000 | 0.000001 | 0.000000 | 0.071112 | 0.000002 |
| 317 voxels (28%) covering 36% of OFusG r | |||||||||
| 93 voxels (8%) covering 5% of iLOC r | |||||||||
| 12 voxels (1%) covering 2% of ICC r | |||||||||
| 5 voxels (0%) covering 0% of sLOC r | |||||||||
| 369 voxels (32%) covering 0% of non-labelled | |||||||||
| B | −14 −90 +04 | 572 | 329 voxels (58%) covering 12% of OP l | 0.000275 | 0.000291 | 0.000022 | 0.636977 | 0.000039 | |
| 36 voxels (6%) covering 4% of OFusG l | |||||||||
| 16 voxels (3%) covering 2% of ICC l | |||||||||
| 15 voxels (3%) covering 1% of LG l | |||||||||
| 176 voxels (31%) covering 0% of non-labelled | |||||||||
| C | +28 −34 +72 | 219 | 185 voxels (84%) covering 6% of PostCG r | 0.043271 | 0.031177 | 0.003464 | 0.009311 | 0.000000 | |
| 3 voxels (1%) covering 0% of PreCG r | |||||||||
| 31 voxels (14%) covering 0% of non-labelled |
| Analysis Unit | Degree Centrality | Betweenness Centrality | ||||
|---|---|---|---|---|---|---|
| t-Value | p-Uncorrected | Rank | t-Value | p-Uncorrected | Rank | |
| VO | 3.26 | 0.001706 | 1 | 2.93 | 0.004589 | 2 |
| OFusG l | 2.71 | 0.008518 | 2 | 2.49 | 0.015310 | 5 |
| OP l | 2.26 | 0.026699 | 7 | 2.14 | 0.035570 | 8 |
| OP r | 2.20 | 0.031157 | 8 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yorita, A.; Kawayama, T.; Inoue, M.; Kinoshita, T.; Oda, H.; Tokunaga, Y.; Tateishi, T.; Shoji, Y.; Uchimura, N.; Abe, T.; et al. Altered Functional Connectivity during Mild Transient Respiratory Impairment Induced by a Resistive Load. J. Clin. Med. 2024, 13, 2556. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092556
Yorita A, Kawayama T, Inoue M, Kinoshita T, Oda H, Tokunaga Y, Tateishi T, Shoji Y, Uchimura N, Abe T, et al. Altered Functional Connectivity during Mild Transient Respiratory Impairment Induced by a Resistive Load. Journal of Clinical Medicine. 2024; 13(9):2556. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092556
Chicago/Turabian StyleYorita, Akiko, Tomotaka Kawayama, Masayuki Inoue, Takashi Kinoshita, Hanako Oda, Yoshihisa Tokunaga, Takahisa Tateishi, Yoshihisa Shoji, Naohisa Uchimura, Toshi Abe, and et al. 2024. "Altered Functional Connectivity during Mild Transient Respiratory Impairment Induced by a Resistive Load" Journal of Clinical Medicine 13, no. 9: 2556. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092556