
Effects of Microtopography on Soil Microbial Community Structure and Abundance in Permafrost Peatlands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Selected Study Area
2.2. Experimental Design and Soil Collection
2.3. Determination of Soil Properties
2.4. Microbial DNA Extraction, Amplification, and Sequencing
2.5. Quantitative PCR (qPCR)
2.6. Statistical Analysis
3. Results
3.1. Physicochemical Properties of the Soil
3.2. Diversity of the Soil Bacterial and Fungal Communities
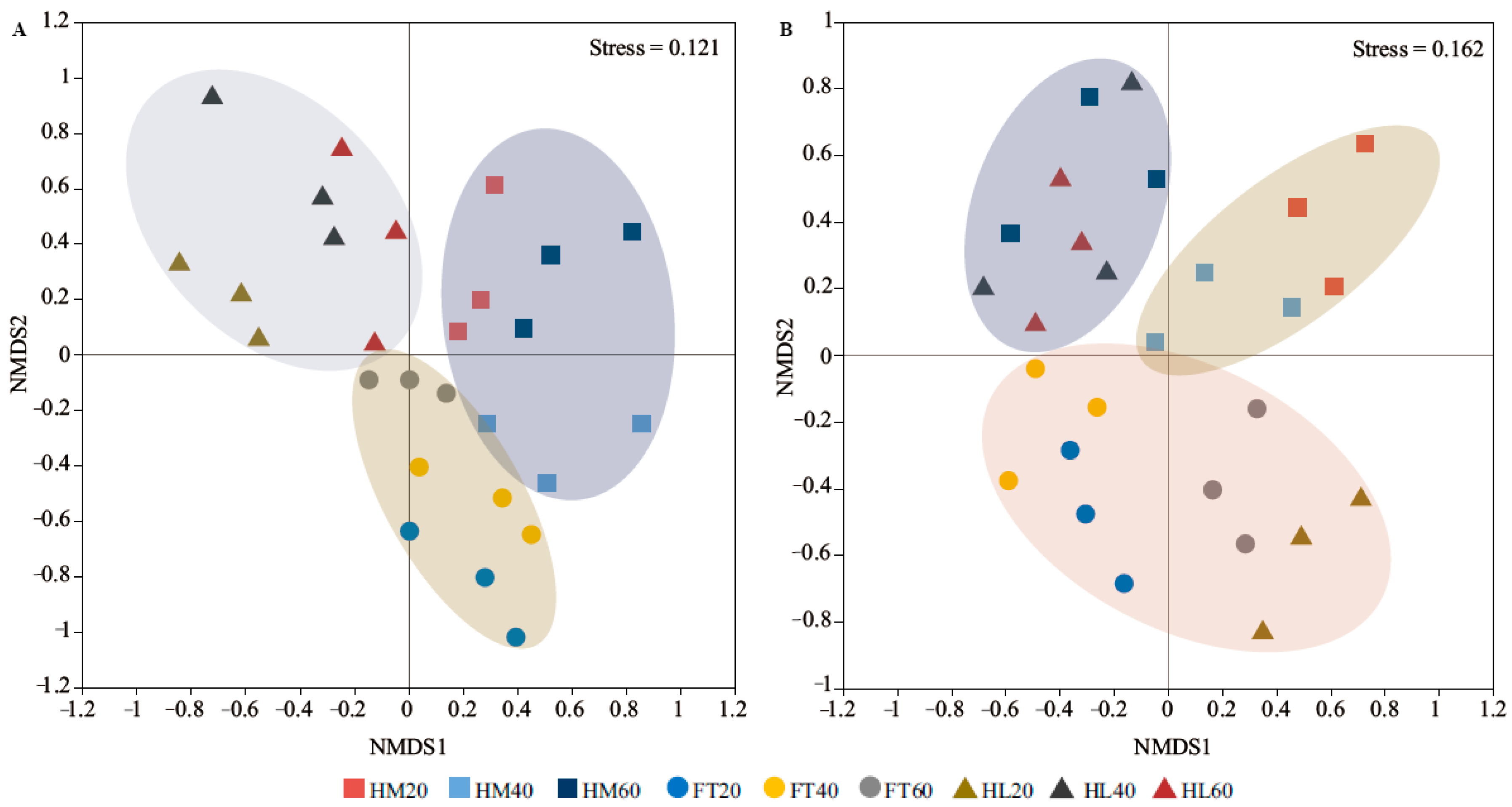
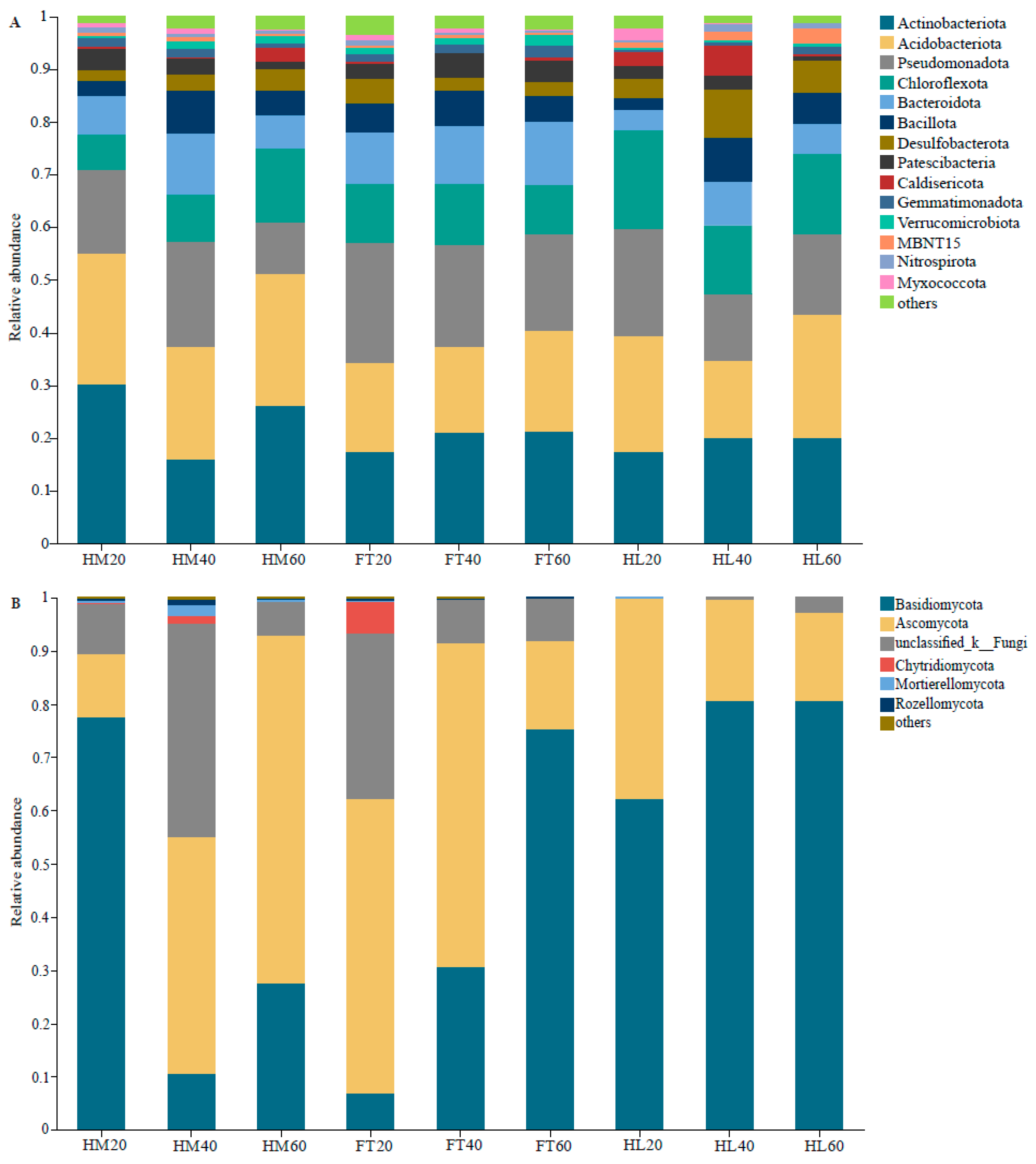
3.3. Structures and Compositions of the Soil Bacterial and Fungal Communities
3.4. Bacterial and Fungal Abundances
3.5. Functional Metabolic Pathways
3.6. Effects of Environmental Factors on the Community Structures and Abundances
4. Discussion
4.1. Effects of Microtopography and Depth on Microbial Diversity
4.2. Changes in Microbial Community Structures in Different Positions
4.3. Variations in Microbial Abundances Response to Microtopography
4.4. Impacts of Environmental Factors on Microbial Community Structures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Qiu, C.J.; Zhu, D.; Ciais, P.; Guenet, B.; Peng, S.S. The role of northern peatlands in the global carbon cycle for the 21st century. Glob. Ecol. Biogeogr. 2020, 29, 956–973. [Google Scholar] [CrossRef]
- Yu, Z.C.; Beilman, D.W.; Frolking, S.; MacDonald, G.M.; Roulet, N.T.; Camill, P.; Charman, D.J. Peatlands and their role in the global carbon cycle. EoS Trans. 2011, 92, 97–98. [Google Scholar] [CrossRef]
- Frolking, S.; Talbot, J.; Jones, M.C.; Treat, C.C.; Kauffman, J.B.; Tuittila, E.S.; Roulet, N. Peatlands in the Earth’s 21st century climate system. Environ. Rev. 2011, 19, 371–396. [Google Scholar] [CrossRef]
- Treat, C.C.; Jones, M.C.; Alder, J.; Frolking, S. Hydrologic controls on peat permafrost and carbon processes: New insights from past and future modeling. Front. Environ. Sci. 2022, 10, 892925. [Google Scholar] [CrossRef]
- Li, Y.; Wan, Z.M.; Sun, L. Simulation of carbon exchange from a permafrost peatland in the Great Hing’an Mountains based on coupmodel. Atmosphere 2021, 13, 44. [Google Scholar] [CrossRef]
- Wang, D.; Zang, S.Y.; Wu, X.W.; Ma, D.L.; Li, M.; Chen, Q.; Liu, X.R.; Zhang, N.N. Soil organic carbon stabilization in permafrost peatlands. Saudi J. Biol. Sci. 2021, 28, 7037–7045. [Google Scholar] [CrossRef]
- Carbognani, M.; Tomaselli, M.; Petraglia, A. Interactions and covariation of ecological drivers control CO2 fluxes in an alpine peatland. Wetlands 2023, 43, 44. [Google Scholar] [CrossRef]
- Voigt, C.; Marushchak, M.E.; Mastepanov, M.; Lamprecht, R.E.; Christensen, T.R.; Dorodnikov, M.; Jackowicz-Korczyński, M.; Lindgren, A.; Lohila, A.; Nykänen, H.; et al. Ecosystem carbon response of an arctic peatland to simulated permafrost thaw. Glob. Chang. Biol. 2019, 25, 1746–1764. [Google Scholar] [CrossRef]
- Lipson, D.A.; Zona, D.; Raab, T.K.; Bozzolo, F.; Mauritz, M.; Oechel, W.C. Water-table height and microtopography control biogeochemical cycling in an arctic coastal tundra ecosystem. Biogeosciences 2012, 9, 577–591. [Google Scholar] [CrossRef]
- Chaudhary, N.; Miller, P.A.; Smith, B. Biotic and abiotic drivers of peatland growth and microtopography: A model demonstration. Ecosystems 2018, 21, 1196–1214. [Google Scholar] [CrossRef]
- Wang, X.W.; Sun, X.X.; Sun, L.; Chen, N.; Du, Y. Small-scale variability of soil quality in permafrost peatland of the Great Hing’an Mountains, northeast China. Water 2022, 14, 2597. [Google Scholar] [CrossRef]
- Bucher, M.; Ofiti, N.O.E.; Malhotra, A. Plant functional types and microtopography mediate climate change responses of fine roots in forested boreal peatlands. Front. For. Glob. Chang. 2023, 6, 1170252. [Google Scholar] [CrossRef]
- Chroňáková, A.; Bárta, J.; Kaštovská, E.; Urbanová, Z.; Picek, T. Spatial heterogeneity of belowground microbial communities linked to peatland microhabitats with different plant dominants. Fems Microbiol. Ecol. 2019, 95, fiz130. [Google Scholar] [CrossRef]
- Myers, B.; Webster, K.L.; Mclaughlin, J.W.; Basiliko, N. Microbial activity across a boreal peatland nutrient gradient: The role of fungi and bacteria. Wetl. Ecol. Manag. 2012, 20, 77–88. [Google Scholar] [CrossRef]
- McGuire, K.L.; Treseder, K.K. Microbial communities and their relevance for ecosystem models: Decomposition as a case study. Soil Biol. Biochem. 2010, 42, 529–535. [Google Scholar] [CrossRef]
- Yu, S.P.; Xu, N.H.; Ding, J.N.; Shi, C.Q. Studies on microbial mechanisms of permafrost carbon conversion to climate warming: Retrospect and prospect. Appl. Ecol. Environ. Res. 2022, 20, 265–283. [Google Scholar] [CrossRef]
- Belyea, L.R.; Malmer, N. Carbon sequestration in peatland: Patterns and mechanisms of response to climate change. Glob. Chang. Biol. 2004, 10, 1043–1052. [Google Scholar] [CrossRef]
- Asemaninejad, A.; Thorn, R.G.; Branfireun, B.A.; Lindo, Z. Vertical stratification of peatland microbial communities follows a gradient of functional types across hummock–hollow microtopographies. Écoscience 2019, 26, 249–258. [Google Scholar] [CrossRef]
- Jiang, L.; Ma, X.Y.; Song, Y.Y.; Gao, S.Q.; Ren, J.S.; Zhang, H.; Wang, X.W. Warming-induced labile carbon change soil organic carbon mineralization and microbial abundance in a northern peatland. Microorganisms 2022, 10, 1329. [Google Scholar] [CrossRef]
- Graham, J.D.; Ricciuto, D.M.; Glenn, N.F.; Hanson, P.J. Incorporating microtopography in a land surface model and quantifying the effect on the carbon cycle. J. Adv. Model. Earth Syst. 2022, 14, e2021MS002721. [Google Scholar] [CrossRef]
- Li, X.W.; Li, X.L.; Shi, Y.; Zhao, S.J.; Liu, J.L.; Lin, Y.Y.; Li, C.L.; Zhang, C.H. Effects of microtopography on soil microbial communities in alpine meadows on the Qinghai-Tibetan Plateau. Catena 2024, 239, 107945. [Google Scholar] [CrossRef]
- Ahn, C.; Gillevet, P.M.; Sikaroodi, M.; Wolf, K.L. An assessment of soil bacterial community structure and physicochemistry in two microtopographic locations of a palustrine forested wetland. Wetl. Ecol. Manag. 2009, 17, 397–407. [Google Scholar] [CrossRef]
- Gao, T.G.; Zhang, T.J.; Wan, X.D.; Kang, S.C.; Sillanpää, M.; Zheng, Y.; Cao, L. Influence of microtopography on active layer thaw depths in Qilian Mountain, northeastern Tibetan Plateau. Environ. Earth Sci. 2016, 75, 382. [Google Scholar] [CrossRef]
- Dong, X.F.; Liu, C.; Wu, X.D.; Man, H.R.; Wu, X.W.; Ma, D.L.; Li, M.; Zang, S.Y. Linking soil organic carbon mineralization with soil variables and bacterial communities in a permafrost-affected tussock wetland during laboratory incubation. Catena 2023, 221, 106783. [Google Scholar] [CrossRef]
- Lu, B.Q.; Song, L.Q.; Zang, S.Y.; Wang, H.X. Warming promotes soil CO2 and CH4 emissions but decreasing moisture inhibits CH4 emissions in the permafrost peatland of the Great Xing’an Mountains. Sci. Total Environ. 2022, 829, 154725. [Google Scholar] [CrossRef]
- Swindles, G.T.; Morris, P.J.; Mullan, D.; Watson, E.J.; Turner, T.E.; Roland, T.P.; Amesbury, M.J.; Kokfelt, U.; Schoning, K.; Pratte, S.; et al. The long-term fate of permafrost peatlands under rapid climate warming. Sci. Rep. 2015, 5, 17951. [Google Scholar] [CrossRef]
- Minick, K.J.; Kelley, A.M.; Miao, G.; Li, X.; Noormets, A.; Mitra, B.; King, J.S. Microtopography alters hydrology, phenol oxidase activity and nutrient availability in organic soils of a coastal freshwater forested wetland. Wetlands 2019, 39, 263–273. [Google Scholar] [CrossRef]
- Parvin, S.; Blagodatskaya, E.; Becker, J.N.; Kuzyakov, Y.; Uddin, S.; Dorodnikov, M. Depth rather than microrelief controls microbial biomass and kinetics of C-, N-, P- and S-cycle enzymes in peatland. Geoderma 2018, 324, 67–76. [Google Scholar] [CrossRef]
- Strack, M.; Waddington, J.M.; Tuittila, E.-S. Effect of water table drawdown on northern peatland methane dynamics: Implications for climate change. Glob. Biogeochem. Cycles 2004, 18, GB400. [Google Scholar] [CrossRef]
- Li, X.; Cui, Y.Q.; Ma, D.L.; Song, D.D.; Liu, L. Vertical distribution of bacterial community diversity in the Greater Khingan Mountain permafrost region. Ecol. Evol. 2022, 12, e9106. [Google Scholar] [CrossRef]
- Wieczorek, D.; Żyszka-Haberecht, B.; Kafka, A.; Lipok, J. Determination of phosphorus compounds in plant tissues: From colourimetry to advanced instrumental analytical chemistry. Plant Methods 2022, 18, 22. [Google Scholar] [CrossRef]
- Li, X.H.; Ai, X.B.; Wang, X.Y.; Li, L.; Xin, Z.J.; Sun, X.Y. Composite phosphorylated-modified rice straw prepared for highly effective immobilisation of cadmium in agriculture contaminated soil. Environ. Chem. 2023, 19, 495–505. [Google Scholar] [CrossRef]
- Bu, N.-S.; Qu, J.-F.; Li, G.; Zhao, B.; Zhang, R.-J.; Fang, C.-M. Reclamation of coastal salt marshes promoted carbon loss from previously-sequestered soil carbon pool. Ecol. Eng. 2015, 81, 335–339. [Google Scholar] [CrossRef]
- McGeough, K.L.; Watson, C.J.; Müller, C.; Laughlin, R.J.; Chadwick, D.R. Evidence that the efficacy of the nitrification inhibitor dicyandiamide (DCD) is affected by soil properties in UK soils. Soil Biol. Biochem. 2016, 94, 222–232. [Google Scholar] [CrossRef]
- Duan, B.X.; Xiao, R.H.; Cai, T.J.; Man, X.L.; Ge, Z.X.; Gao, M.L.; Mencuccini, M. Understory species composition mediates soil greenhouse gas fluxes by affecting bacterial community diversity in boreal forests. Front. Microbiol. 2023, 13, 1090169. [Google Scholar] [CrossRef]
- Song, Y.S.; Liu, R.L.; Yang, L.R.; Xiao, X.Y.; He, G.H. Effects of moss-dominated biocrusts on soil microbial community structure in an ionic rare earth tailings area of southern China. Toxics 2022, 10, 782. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Wu, F.Z.; Yang, W.Q.; Tan, B.; He, W. Variations in bacterial communities during foliar litter decomposition in the winter and growing seasons in an alpine forest of the eastern Tibetan Plateau. Can. J. Microbiol. 2016, 62, 35–48. [Google Scholar] [CrossRef]
- Sandeu, M.M.; Maffo, C.G.T.; Dada, N.; Njiokou, F.; Hughes, G.L.; Wondji, C.S. Seasonal variation of microbiota composition in anopheles gambiae and anopheles coluzzii in two different eco-geographical localities in Cameroon. Med. Vet. Entomol. 2022, 36, 269–282. [Google Scholar] [CrossRef]
- Hu, J.Y.; Chen, Q.; Zhong, S.N.; Liu, Y.P.; Gao, Q.; Graham, E.B.; Chen, H.; Sun, W.L. Insight into co-hosts of nitrate reduction genes and antibiotic resistance genes in an urban river of the Qinghai-Tibet Plateau. Water Res. 2022, 225, 119189. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and extensible approach for metagenome inference. bioRxiv 2019. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Jaatinen, K.; Fritze, H.; Laine, J.; Laiho, R. Effects of short- and long-term water-level drawdown on the populations and activity of aerobic decomposers in a boreal peatland. Glob. Chang. Biol. 2007, 13, 491–510. [Google Scholar] [CrossRef]
- Zhao, H.; Wei, D.; Yan, Y.; Wu, J.B.; Wang, X.D. Alpine hummocks drive plant diversity and soil fertile islands on the Tibetan Plateau. Wetlands 2020, 40, 1217–1227. [Google Scholar] [CrossRef]
- Barreto, C.; Lindo, Z. Drivers of decomposition and the detrital invertebrate community differ across a hummock-hollow microtopology in boreal peatlands. Écoscience 2018, 25, 39–48. [Google Scholar] [CrossRef]
- Lamit, L.J.; Romanowicz, K.J.; Potvin, L.R.; Lennon, J.T.; Tringe, S.G.; Chimner, R.A.; Kolka, R.K.; Kane, E.S.; Lilleskov, E.A. Peatland microbial community responses to plant functional group and drought are depth-dependent. Mol. Ecol. 2021, 30, 5119–5136. [Google Scholar] [CrossRef]
- Seward, J.; Carson, M.A.; Lamit, L.J.; Basiliko, N.; Yavitt, J.B.; Lilleskov, E.; Schadt, C.W.; Smith, D.S.; Mclaughlin, J.; Mykytczuk, N.; et al. Peatland microbial community composition is driven by a natural climate gradient. Microb. Ecol. 2020, 80, 593–602. [Google Scholar] [CrossRef]
- Ding, Z.L.; Gong, L.; Zhu, H.Q.; Tang, J.H.; Li, X.C.; Zhang, H. Changes in soil microbial communities under mixed organic and inorganic nitrogen addition in temperate forests. Forests 2022, 14, 21. [Google Scholar] [CrossRef]
- Xiao, Y.; Yu, Y.N.; Wang, Y.; Wang, X.Q.; Wang, Y.Y.; Dai, W.; Luan, Y.N. Effect of two exogenous organic acids on the excitation effect of soil organic carbon in Beijing, China. Forests 2022, 13, 487. [Google Scholar] [CrossRef]
- Speirs, L.B.M.; Rice, D.T.F.; Petrovski, S.; Seviour, R.J. The phylogeny, biodiversity, and ecology of the Chloroflexi in activated sludge. Front. Microbiol. 2019, 10, 2015. [Google Scholar] [CrossRef]
- Serkebaeva, Y.M.; Kim, Y.; Liesack, W.; Dedysh, S.N. Pyrosequencing-based assessment of the bacteria diversity in surface and subsurface peat layers of a northern wetland, with focus on poorly studied phyla and candidate divisions. PLoS ONE 2013, 8, e63994. [Google Scholar] [CrossRef]
- Juottonen, H.; Eiler, A.; Biasi, C.; Tuittila, E.-S.; Yrjälä, K.; Fritze, H. Distinct anaerobic bacterial consumers of cellobiose-derived carbon in boreal fens with different CO2/CH4 production ratios. Appl. Environ. Microb. 2017, 83, e02533-16. [Google Scholar] [CrossRef]
- Huang, Q.Y.; Yang, F.; Cao, H.J.; Cheng, J.H.; Jiang, M.Y.; Li, M.H.; Ni, H.W.; Xie, L.H. Comparison of microbial diversity of two typical volcanic soils in Wudalianchi, China. Microorganisms 2024, 12, 656. [Google Scholar] [CrossRef]
- Asemaninejad, A.; Thorn, R.G.; Lindo, Z. Vertical distribution of fungi in hollows and hummocks of boreal peatlands. Fungal Ecol. 2017, 27, 59–68. [Google Scholar] [CrossRef]
- Peltoniemi, K.; Adamczyk, S.; Fritze, H.; Minkkinen, K.; Pennanen, T.; Penttilä, T.; Sarjala, T.; Laiho, R. Site fertility and soil water-table level affect fungal biomass production and community composition in boreal peatland forests. Environ. Microbiol. 2021, 23, 5733–5749. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef]
- Wang, M.; Tian, J.Q.; Bu, Z.J.; Lamit, L.J.; Chen, H.; Zhu, Q.; Peng, C.H. Structural and functional differentiation of the microbial community in the surface and subsurface peat of two minerotrophic fens in China. Plant Soil 2019, 437, 21–40. [Google Scholar] [CrossRef]
- Nungesser, M.K. Modelling microtopography in boreal peatlands: Hummocks and hollows. Ecol. Model. 2003, 165, 175–207. [Google Scholar] [CrossRef]
- Andersen, R.; Chapman, S.J.; Artz, R.R.E. Microbial communities in natural and disturbed peatlands: A review. Soil Biol. Biochem. 2013, 57, 979–994. [Google Scholar] [CrossRef]
- Rosset, T.; Binet, S.; Rigal, F.; Gandois, L. Peatland dissolved organic carbon export to surface waters: Global significance and effects of anthropogenic disturbance. Geophys. Res. Lett. 2022, 49, e2021GL096616. [Google Scholar] [CrossRef]
- Ward, C.P.; Nalven, S.G.; Crump, B.C.; Kling, G.W.; Cory, R.M. Photochemical alteration of organic carbon draining permafrost soils shifts microbial metabolic pathways and stimulates respiration. Nat. Commun. 2017, 8, 772. [Google Scholar] [CrossRef]
- Breeuwer, A.; Robroek, B.J.M.; Limpens, J.; Heijmans, M.M.P.D.; Schouten, M.G.C.; Berendse, F. Decreased summer water table depth affects peatland vegetation. Basic Appl. Ecol. 2009, 10, 330–339. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Heděnec, P.; Rui, J.P.; Lin, Q.; Yao, M.J.; Li, J.B.; Li, H.; Frouz, J.; Li, X.Z. Functional and phylogenetic response of soil prokaryotic community under an artificial moisture gradient. Appl. Soil Ecol. 2018, 124, 372–378. [Google Scholar] [CrossRef]
- Liu, Y.; Du, J.Q.; Xu, X.L.; Kardol, P.; Hu, D. Microtopography-induced ecohydrological effects alter plant community structure. Geoderma 2020, 362, 114119. [Google Scholar] [CrossRef]
- Ma, X.Y.; Song, Y.Y.; Song, C.C.; Wang, X.W.; Wang, N.N.; Gao, S.Q.; Cheng, X.F.; Liu, Z.D.; Gao, J.L.; Du, Y. Effect of nitrogen addition on soil microbial functional gene abundance and community diversity in permafrost peatland. Microorganisms 2021, 9, 2498. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, L.X.; Duan, W.B. Responses of bacterial and fungal communities to short-term nitrogen and phosphorus additions in temperate forest soil aggregates in northeastern China. Appl. Soil Ecol. 2024, 197, 105341. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Soil Depth | pH | TP (g·kg−1) | TN (g·kg−1) | SWC (%) | TOC (g·kg−1) | DOC (g·kg−1) | NO3–-N (mg·kg−1) | NH4+-N (mg·kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| Hummock | 0–20 | 4.95 ± 0.01 Aa | 2.14 ± 0.07 Bc | 14.51 ± 0.72 Cb | 58.15 ± 3.28 Bb | 215.32 ± 22.15 Ba | 0.41 ± 0.06 Aa | 7.16 ± 1.09 Ab | 72.57 ± 7.92 Aa |
| 20–40 | 4.76 ± 0.01 Ca | 2.38 ± 0.13 Ac | 20.27 ± 1.4 Ab | 66.67 ± 4.91 Bb | 319.63 ± 36.19 Ab | 0.33 ± 0.05 Aa | 5.32 ± 0.82 Ba | 46.81 ± 5.21 Ba | |
| 40–60 | 4.89 ± 0.01 Ba | 2.34 ± 0.12 ABb | 17.12 ± 1.19 Bb | 69.22 ± 5.41 Ab | 247.51 ± 26.31 Bb | 0.12 ± 0.01 Ba | 4.17 ± 0.72 Ba | 32.15 ± 3.41 Cb | |
| Flat | 0–20 | 4.88 ± 0.01 Ab | 2.53 ± 0.19 Bb | 18.68 ± 1.24 Ba | 71.14 ± 4.32 Ba | 221.36 ± 24.24 Ca | 0.15 ± 0.05 Ab | 8.13 ± 1.21 Bab | 65.88 ± 6.84 Aa |
| 20–40 | 4.63 ± 0.05 Bb | 2.95 ± 0.20 Ab | 24.99 ± 0.74 Aa | 82.36 ± 5.12 Aa | 376.88 ± 37.21 Aab | 0.10 ± 0.03 Ab | 6.62 ± 1.01 ABa | 50.19 ± 5.17 Ba | |
| 40–60 | 4.69 ± 0.02 Bb | 2.79 ± 0.15 ABa | 20.65 ± 1.39 Ba | 80.81 ± 5.24 ABa | 285.07 ± 26.47 Bab | 0.07 ± 0.04 Ab | 5.01 ± 0.62 Aa | 36.29 ± 3.02 Cb | |
| Hollow | 0–20 | 4.71 ± 0.02 Ac | 2.92 ± 0.21 Ba | 20.05 ± 1.37 Ba | 79.72 ± 5.53 Ba | 258.05 ± 33.19 Ca | 0.11 ± 0.02 Ab | 9.81 ± 1.32 Aa | 58.26 ± 5.91 Aa |
| 20–40 | 4.57 ± 0.07 Bb | 3.41 ± 0.23 Aa | 25.01 ± 1.53 Aa | 93.09 ± 6.34 Aa | 423.36 ± 39.75 Aa | 0.09 ± 0.03 Bb | 7.25 ± 1.07 Ba | 57.10 ± 5.72 Aa | |
| 40–60 | 4.63 ± 0.05 ABb | 2.96 ± 0.20 ABa | 22.33 ± 1.41 ABa | 86.35 ± 5.74 ABa | 336.13 ± 34.32 Ba | 0.06 ± 0.01 Bb | 5.32 ± 0.87 Ba | 41.35 ± 4.28 Ba |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Fu, L.; Ma, D.; Wang, X.; Liu, A. Effects of Microtopography on Soil Microbial Community Structure and Abundance in Permafrost Peatlands. Microorganisms 2024, 12, 867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050867
Zhang M, Fu L, Ma D, Wang X, Liu A. Effects of Microtopography on Soil Microbial Community Structure and Abundance in Permafrost Peatlands. Microorganisms. 2024; 12(5):867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050867
Chicago/Turabian StyleZhang, Man, Lingyu Fu, Dalong Ma, Xu Wang, and Anwen Liu. 2024. "Effects of Microtopography on Soil Microbial Community Structure and Abundance in Permafrost Peatlands" Microorganisms 12, no. 5: 867. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050867