
A Review on the Methodology and Use of the Pregnant Mouse Model in the Study of Brucella Reproductive Pathogenesis and Its Abortifacient Effect
 , and
, and
Abstract
:1. Brucellosis and Brucella Reproductive Pathogenesis
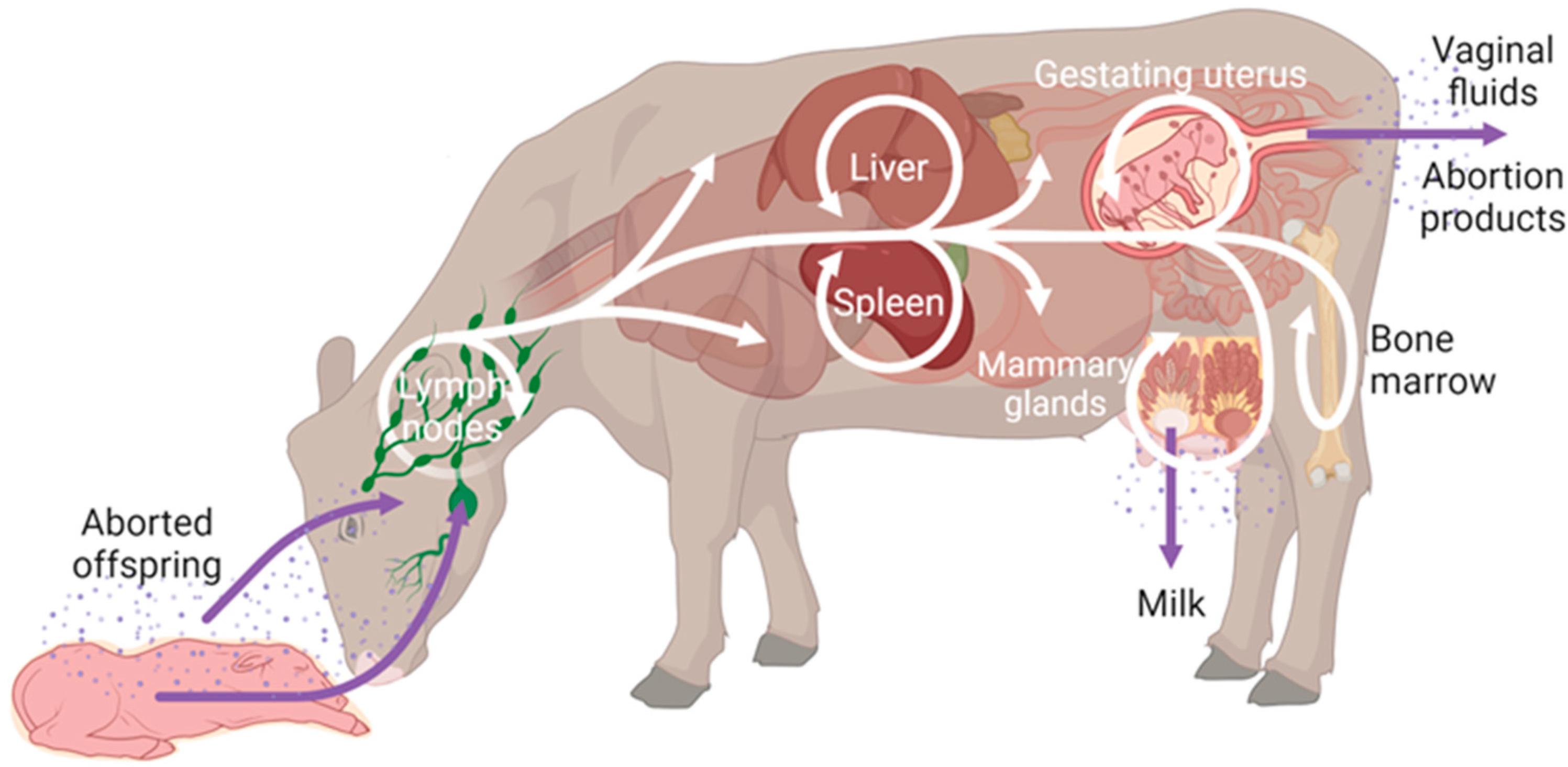
2. Placentation in Brucella Natural Hosts and Mouse
3. The Pregnant Mouse Model in Brucella Research
3.1. Methodological Aspects of the Model
3.1.1. Mouse Pregnancy Outcome and Abortion Readout


{kind=link}
{kind=link}
{kind=link}
| Mouse Strain/ Mating Conditions | Time of Infection/ Time of Euthanasia | Brucella Strain, Administration Route and Dose (CFU/Mouse) | Main Findings |
|---|---|---|---|
| Bosseray, N. Colonization of Mouse Placentas by Brucella abortus Inoculated during Pregnancy. Br. J. Exp. Pathol. 1980, 61, 361–368. [61] | |||
| CD1 Mating: 1–2 in-house-born females were kept for 3 nights with 1 in-house-born male. Day 1 = Vaginal plug | Infection at 3, 7, 11, or 15 dpc Euthanasia at 18 dpc (15, 11, 7, or 3 dpi) | B. abortus 544 IP, 1.5 × 101–5 IV, 2.5 × 101–4 SC, 1.0 × 102–8 |
|
| Bosseray, N. Mother to Young Transmission of Brucella abortus Infection in Mouse Model. Ann. De Rech. Vet. 1982, 13, 341–349. [63] | |||
| CD-1 Mating: as in Bosseray, 1980 [61] | Infection at 6 dpc Euthanasia at different times post-birth | B. abortus 544 Wb B. abortus 544 CO2-independent B. abortus 544 Strp-resistant IP, 1.5–1.8 × 105 |
|
| Bosseray, N. Kinetics of Placental Colonization of Mice Inoculated Intravenously with Brucella abortus at Day 15 of Pregnancy. Br. J. Exp. Pathol. 1983, 64, 612–616. [62] | |||
| CD-1, OF1 Mating: as in Bosseray, 1980 [61] | Infection at 15 dpc Euthanasia at 5, 10, 20, or 40-min or 1, 1.5, 2, 4, 6, 24, 48, or 72 h p.i. | B. abortus 544 IV, 1–2 × 104 |
|
| Bosseray, N. Vaccine and Serum-Mediated Protection against Brucella Infection of Mouse Placenta. Br. J. Exp. Pathol. 1983, 64, 617–625. [64] | |||
| CD-1, OF1 Mating: as in Bosseray, 1980 [61] | Infection at 7 or 15 dpc Euthanasia at 16 or 18 dpc | B. abortus 544 IP, 1.8–2.1 × 105 IV, 1.8–4.5 × 104/1.8–2.1 × 105 |
|
| Bosseray, N.; Plommet, M. Serum- and Cell-Mediated Immune Protection of Mouse Placenta and Fetus against a Brucella abortus Challenge: Expression of Barrier Effect of Placenta. Placenta 1988, 9, 65–79. doi:10.1016/0143-4004(88)90074-4. [65] | |||
| OF1, DBA/2 Mating: as in Bosseray, 1980 [61] | Infection at 12 or 14 dpc Euthanasia at 3 or 5 dpi | B. abortus 544 IV, 1.8–4.5 × 104/1.8–2.1 × 105 |
|
| Tobias, L.; Schurig, G.G.; Cordes, D.O. Comparative Behaviour of Brucella abortus Strains 19 and RB51 in the Pregnant Mouse. Res. Vet. Sci. 1992, 53, 179–183. doi:10.1016/0034-5288(92)90107-D. [37] | |||
| BALB/C | Infection at 9 dpc Euthanasia at 18 dpc (9 dpi) | B. abortus 2308 (IP, 105.7) B. abortus S19 (IP, 107.5) B. abortus RB51 (IP, 109.5) |
|
| Tobias, L.; Cordes, D.O.; Schurig, G.G. Placental Pathology of the Pregnant Mouse Inoculated with Brucella abortus Strain 2308. Vet. Pathol. 1993, 30, 119–129. doi:10.1177/030098589303000204. [38] | |||
| BALB/C Mating: in-house-born mice were individually mated. Day 1 = Vaginal plug | Infection at 9 dpc Euthanasia at 12, 14, 16, or 18 dpc (2, 5, 7, or 9 dpi) | B. abortus 2308 IP, 104.7/5.7/6/6.7 |
|
| Kim, S.; Dong, S.L.; Watanabe, K.; Furuoka, H.; Suzuki, H.; Watarai, M. Interferon-γ Promotes Abortion Due to Brucella Infection in Pregnant Mice. BMC Microbiol. 2005, 5, 22. doi:10.1186/1471-2180-5-22. [39] | |||
| ICR Mating: 6–10-week-old females individually mated to 6–10-week-old males. Day 0.5 = Vaginal plug | Infection at 3.5, 4.5, 6.5, 9.5, or 14.5 dpc Euthanasia at 18.5 dpc (15, 14, 12, 9, or 4 dpi) | B. abortus 544 B. abortus ΔvirB4 B. abortus S19 IP, 104 |
|
| Watanabe, K.; Iwai, N.; Tachibana, M.; Furuoka, H.; Suzuki, H.; Watarai, M. Regulated upon Activation Normal T-Cell Expressed and Secreted (RANTES) Contributes to Abortion Caused by Brucella abortus Infection in Pregnant Mice. J. Vet. Med. Sci. 2008, 70, 681–686. doi:10.1292/jvms.70.681. [66] | |||
| BALB/c, IFN-γ KO BALB/c Mating: as in Kim et al., 2005 [39] | Infection at 4.5 dpc Euthanasia at 18.5 dpc | B. abortus 544 B. abortus ΔvirB4 IP, 104 |
|
| Watanabe, K.; Tachibana, M.; Tanaka, S.; Furuoka, H.; Horiuchi, M.; Suzuki, H.; Watarai, M. Heat Shock Cognate Protein 70 Contributes to Brucella Invasion into Trophoblast Giant Cells That Cause Infectious Abortion. BMC Microbiol. 2008, 8, 212. doi:10.1186/1471-2180-8-212. [67] | |||
| ICR Mating: as in Kim et al., 2005 [39] | Infection at 4.5 dpc Euthanasia at 18.5 dpc | B. abortus 544 IP, 104 |
|
| Hashino, M.; Kim, S.; Tachibana, M.; Shimizu, T.; Watarai, M. Vertical Transmission of Brucella abortus Causes Sterility in Pregnant Mice. J. Vet. Med. Sci. 2012, 74, 1075–1077. doi:10.1292/jvms.11-0566. [68] | |||
| ICR Mating: as in Kim et al., 2005 [39] | Infection at 6.5 dpc Euthanasia at different times post-birth | B. abortus (strain not specified; presumably 544) IP, 104 |
|
| Wang, Z.; Wang, S.S.; Wang, G.L.; Wu, T.L.; Lv, Y.L.; Wu, Q.M. A Pregnant Mouse Model for the Vertical Transmission of Brucella melitensis. Vet. J. 2014, 200, 116–121. doi:10.1016/j.tvjl.2013.12.021. [40] | |||
| ICR Mating: as in Kim et al., 2005 [39] | Infection at 4.5 dpc Euthanasia at 18.5 dpc (14 dpi) | B. melitensis NI IP, 103–6 |
|
| Keestra-Gounder, A.M.; Byndloss, M.X.; Seyffert, N.; Young, B.M.; Chávez-Arroyo, A.; Tsai, A.Y.; Cevallos, S.A.; Winter, M.G.; Pham, O.H.; Tiffany, C.R.; et al. NOD1 and NOD2 Signalling Links ER Stress with Inflammation. Nature 2016, 532, 394–397. doi:10.1038/nature17631. [71] | |||
| C57BL/6 Mating: in-house mating. No further specifications | Infection at 5 dpc Euthanasia at 8, 12, and 18 dpc (3, 7, and 13 dpi) | B. abortus 2308 B. abortus 2308 ∆vceC IP, 105 |
|
| Barbier, T.; Machelart, A.; Zúñiga-Ripa, A.; Plovier, H.; Hougardy, C.; Lobet, E.; Willemart, K.; Muraille, E.; De Bolle, X.; Van Schaftingen, E.; et al. Erythritol Availability in Bovine, Murine and Human Models Highlights a Potential Role for the Host Aldose Reductase during Brucella Infection. Front. Microbiol. 2017, 8, 1088. doi:10.3389/fmicb.2017.01088. [69] | |||
| C57BL/6 Mating: oestrus was synchronised 3 days before mating. Each female was mated with one male and then isolated. Adapted from Bosseray, 1982 and Kim et al., 2005 [39,63] | Infection at 6 and 14 dpc Euthanasia at 15 dpc (9 and 1 dpi) | B. abortus 2308 B. abortus 2308 ∆eryA B. abortus 2308 ∆eryH IP, 105 |
|
| Byndloss, M.X.; Tsai, A.Y.; Walker, G.T.; Miller, C.N.; Young, B.M.; English, B.C.; Seyffert, N.; Kerrinnes, T.; de Jong, M.F.; Atluri, V.L.; et al. Brucella abortus Infection of Placental Trophoblasts Triggers Endoplasmic Reticulum Stress-Mediated Cell Death and Fetal Loss via Type IV Secretion System-Dependent Activation of CHOP. mBio 2019, 10, e01538–19. doi:10.1128/mBio.01538-19. [41] | |||
| C57BL/6J Mating: as in Keestra-Gounder et al., 2016 [71] | Infection at 5 dpc Euthanasia at 18 dpc (13 dpi) | B. abortus 2308 B. abortus 2308 ∆virB2 B. abortus 2308 ∆vceC IP, 105 |
|
| Adetunji, S.A.; Faustman, D.L.; Adams, L.G.; Garcia-Gonzalez, D.G.; Hensel, M.E.; Khalaf, O.H.; Arenas-Gamboa, A.M. Brucella abortus and Pregnancy in Mice: Impact of Chronic Infection on Fertility and the Role of Regulatory T Cells in Tissue Colonization. Infect. Immun. 2020, 88, e00257-20. doi:10.1128/IAI.00257-20. [82] | |||
| ICR Mating: oestrus was synchronised by caging of mice with male house bedding for 3–5 days | Infection 8 weeks prior to mating Euthanasia at 18 dpc | B. abortus S2308 B. abortus S19 B. abortus S2308 ∆virB2 IP, 106 |
|
| Ren, J.; Hou, H.; Zhao, W.; Wang, J.; Peng, Q. Administration of Exogenous Progesterone Protects Against Brucella abortus Infection–Induced Inflammation in Pregnant Mice. J. Infect. Dis. 2021, 224, 532–543. doi:10.1093/infdis/jiaa722. [83] | |||
| C57BL/6J Mating: as in Keestra-Gounder et al., 2016 [71] | Infection at 5 dpc Euthanasia at 18 dpc (13 dpi) | B. abortus 2308 IP, 105 |
|
| Tsai, A.Y.; Byndloss, M.X.; Seyffert, N.; Winter, M.G.; Young, B.M.; Tsolis, R.M. Tumor Necrosis Factor Alpha Contributes to Inflammatory Pathology in the Placenta during Brucella abortus Infection. Infect. Immun. 2022, 90, e0001322. doi:10.1128/iai.00013-22. [72] | |||
| C57BL/6J Mating: as in Kim et al., 2005 [39] | Infection at 5 dpc Euthanasia at 18 dpc (13 dpi) | B. abortus 2308 B. abortus 2308 ∆virB2 B. abortus 2308 ∆vceC IP, 105 |
|
| Poveda-Urkixo, I.; Ramírez, G.A.; Grilló, M.-J. Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens 2022, 11, 279. doi:10.3390/pathogens11030279. [73] | |||
| CD1 Mating: 5 females were mated with 1 male for 2 days | Infection at 4.5 ± 1 dpc Euthanasia at 5.5, 7.5, 9.5, 11.5, 14.5, and 18.5 ± 1 dpc (1, 3, 5, 7, 10, and 14 dpi) | B. melitensis 16M B. melitensis Rev1 B. suis biovar 2 CITA198 IP, 6–7 × 105 |
|
| Braz, H.M.B.; Silva, M.F.; Carvalho, T.P. de; Silva, L.A. da; Soares, J.B.; Costa, F.B.; Sossai, B.G.; Paixão, T.A. da; Santos, R.L. Pathogenesis of Brucella ovis in Pregnant Mice and Protection Induced by the Candidate Vaccine Strain B. ovis ΔAbcBA. Vaccine 2022, 40, 4617–4624. doi:10.1016/j.vaccine.2022.06.044. [76] | |||
| Balb/c | Infection at 5 dpc Euthanasia at 17 dpc (12 dpi) | B. ovis ATCC 25840 B. ovis ΔabcBA IP, 106 |
|
| Mena-Bueno, S.; Poveda-Urkixo, I.; Irazoki, O.; Palacios, L.; Cava, F.; Zabalza-Baranguá, A.; Grilló, M.J. Brucella melitensis Wzm/Wzt System: Changes in the Bacterial Envelope Lead to Improved Rev1Δwzm Vaccine Properties. Front. Microbiol. 2022, 13, 908495. doi:10.3389/fmicb.2022.908495. [75] | |||
| CD1 Mating: as in Poveda-Urkixo et al., 2022 [73] | Infection at 4.5 ± 1 dpc Euthanasia at 18.5 dpc (14 dpi) | B. melitensis Rev1 (IP, 106) Rev1Δwzm (IP, 107) |
|
| Zabalza-Baranguá, A.; Poveda-Urkixo, I.; Mena-Bueno, S.; Ramírez, G.A.; De Bolle, X.; Grilló, M.J. Vaccine Properties of Brucella melitensis 16MΔwzm and Reactivation of Placental Infection in Pregnant Sheep. Vaccine 2023, 41, 1554–1566. doi:10.1016/j.vaccine.2023.01.017. [74] | |||
| CD1 Mating: as in Poveda-Urkixo et al., 2022 [73] | Infection at 4.5 ± 1 dpc Euthanasia at 18.5 dpc (14 dpi) | B. melitensis Rev1 (IP, 106) B. melitensis 16M (IP, 106) B. melitensis 16MΔwzm (IP, 107) |
|
3.1.2. Inoculation Day
3.1.3. Route of Administration
3.1.4. Dose
3.2. Reproductive Pathogenesis in the Pregnant Mouse
3.3. Vertical Transmission
3.4. Vaccine Safety Screening
4. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
| 1 | When not provided, data were extracted from graphs using the informatic tool PlotDigitizer, 3.1.5, 2024, https://plotdigitizer.com, accessed on 27 February 2024. |
| 2 | See note 1 above. |
| 3 | See note 1 above. |
| 4 | See note 1 above. |
| 5 | See note 1 above. |
| 6 | See note 1 above. |
| 7 | See note 1 above. |
References
- Moreno, E.; Cloeckaert, A.; Moriyón, I. Brucella Evolution and Taxonomy. Vet. Microbiol. 2002, 90, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Byndloss, M.X.; Tsolis, R.M. Brucella spp. Virulence Factors and Immunity. Annu. Rev. Anim. Biosci. 2016, 4, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Zinsstag, J.; Schelling, E.; Solera, J.; Blasco, J.M.; Moriyón, I. Brucellosis BT—Handbook of Zoonoses. In Handbook of Zoonoses; Palmer, S.R., Soulsby, L., Torgeson, P.R., Brown, D.G., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA, 2011; pp. 54–62. [Google Scholar]
- Meinershagen, W.A.; Frank, F.W.; Waldhalm, D.G. Brucella ovis as a Cause of Abortion in Ewes. Am. J. Vet. Res. 1974, 35, 723–724. [Google Scholar] [PubMed]
- Xavier, M.N.; Paixão, T.A.; Poester, F.P.; Lage, A.P.; Santos, R.L. Pathological, Immunohistochemical and Bacteriological Study of Tissues and Milk of Cows and Fetuses Experimentally Infected with Brucella abortus. J. Comp. Pathol. 2009, 140, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.D.; Cheville, N.F.; Meador, V.P. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. II. Ultrastructural Studies. Vet. Pathol. 1986, 23, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Alton, G.; Forsyth, J. Brucella. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Xavier, M.N.; Paixao, T.A.; den Hartigh, A.B.; Tsolis, R.M.; Santos, R.L. Pathogenesis of Brucella spp. Open Vet. Sci. J. 2010, 4, 109–118. [Google Scholar] [CrossRef]
- Corbel, M.J.; Alton, G.; Banai, M.; Díaz, R. Brucellosis in Humans and Animals; World Health Organization: Geneva, Switzerland, 2006; ISBN 9241547138. [Google Scholar]
- Renukaradhya, G.J.; Isloor, S.; Rajasekhar, M. Epidemiology, Zoonotic Aspects, Vaccination and Control/Eradication of Brucellosis in India. Vet. Microbiol. 2002, 90, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis Emergence Linked to Agricultural Intensification and Environmental Change. Proc. Natl. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef] [PubMed]
- Ducrotoy, M.J.; Bertu, W.J.; Ocholi, R.A.; Gusi, A.M.; Bryssinckx, W.; Welburn, S.; Moriyón, I. Brucellosis as an Emerging Threat in Developing Economies: Lessons from Nigeria. PLoS Negl. Trop. Dis. 2014, 8, e3008. [Google Scholar] [CrossRef]
- Ducrotoy, M.; Bertu, W.J.; Matope, G.; Cadmus, S.; Conde-Álvarez, R.; Gusi, A.M.; Welburn, S.; Ocholi, R.; Blasco, J.M.; Moriyón, I. Brucellosis in Sub-Saharan Africa: Current Challenges for Management, Diagnosis and Control. Acta Trop. 2017, 165, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Xu, W.; Lu, S.; Jiang, J.; Zhou, J.; Shao, Z.; Liu, X.; Xu, L.; Xiong, Y.; Zheng, H.; et al. Landscape of Emerging and Re-Emerging Infectious Diseases in China: Impact of Ecology, Climate, and Behavior. Front. Med. 2018, 12, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Mcdermott, J.J.; Grace, D.; Zinsstag, J. Economics of Brucellosis Impact and Control in Low-Income Countries. Rev. Sci. Tech. l’OIE 2013, 32, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Zhou, H.; Xiong, W.; Gilbert, M.; Huang, Z.; Yu, J.; Yin, W.; Wang, L.; Chen, Q.; Li, Y.; et al. Changing Epidemiology of Human Brucellosis, China, 1955–2014. Emerg. Infect. Dis. 2017, 23, 184–194. [Google Scholar] [CrossRef]
- Ran, X.; Cheng, J.; Wang, M.; Chen, X.; Wang, H.; Ge, Y.; Ni, H.; Zhang, X.-X.; Wen, X. Brucellosis Seroprevalence in Dairy Cattle in China during 2008–2018: A Systematic Review and Meta-Analysis. Acta Trop. 2019, 189, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Laine, C.G.; Johnson, V.E.; Scott, H.M.; Arenas-Gamboa, A.M. Global Estimate of Human Brucellosis Incidence. Emerg. Infect Dis. 2023, 29, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Mangen, M.J.; Pfeiffer, D.; Chilonda, P. Bovine Brucellosis in Sub-Saharan Africa: Estimation of Meat and Milk off Take Potential; FAO: Rome, Italy, 2005. [Google Scholar]
- Singh, B.B.; Kostoulas, P.; Gill, J.P.S.; Dhand, N.K. Cost-Benefit Analysis of Intervention Policies for Prevention and Control of Brucellosis in India. PLoS Negl. Trop. Dis. 2018, 12, e0006488. [Google Scholar] [CrossRef] [PubMed]
- Blasco, J.M.; Moreno, E.; Moriyón, I. Brucellosis Vaccines and Vaccine Candidates. In Veterinary Vaccines. Principles and Applications; Metwally, S., Vilioen, G.J., El Idrissi, A., Eds.; FAO: Rome, Italy; Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 295–316. [Google Scholar]
- Meador, V.P.; Deyoe, B.L. Intracellular Localization of Brucella abortus in Bovine Placenta. Vet. Pathol. 1989, 26, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, P. The Epidemiology of Bovine Brucellosis. Adv. Vet. Sci. Comp. Med. 1980, 24, 69–98. [Google Scholar] [PubMed]
- von Bargen, K.; Gagnaire, A.; Arce-Gorvel, V.; de Bovis, B.; Baudimont, F.; Chasson, L.; Bosilkovski, M.; Papadopoulos, A.; Martirosyan, A.; Henri, S.; et al. Cervical Lymph Nodes as a Selective Niche for Brucella during Oral Infections. PLoS ONE 2015, 10, e0121790. [Google Scholar] [CrossRef] [PubMed]
- Celli, J.; de Chastellier, C.; Franchini, D.-M.; Pizarro-Cerda, J.; Moreno, E.; Gorvel, J.-P. Brucella Evades Macrophage Killing via VirB-Dependent Sustained Interactions with the Endoplasmic Reticulum. J. Exp. Med. 2003, 198, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Comerci, D.J.; Martinez-Lorenzo, M.J.; Sieira, R.; Gorvel, J.-P.; Ugalde, R.A. Essential Role of the VirB Machinery in the Maturation of the Brucella abortus-Containing Vacuole. Cell Microbiol. 2001, 3, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Cerdá, J.; Méresse, S.; Parton, R.G.; van der Goot, G.; Sola-Landa, A.; Lopez-Goñi, I.; Moreno, E.; Gorvel, J. Brucella abortus Transits through the Autophagic Pathway and Replicates in the Endoplasmic Reticulum of Nonprofessional Phagocytes. Infect. Immun. 1998, 66, 5711–5724. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.; Child, R.; Wehrly, T.D.; Hansen, B.; Hwang, S.; López-Otin, C.; Virgin, H.W.; Celli, J. Selective Subversion of Autophagy Complexes Facilitates Completion of the Brucella Intracellular Cycle. Cell Host. Microbe 2012, 11, 33–45. [Google Scholar] [CrossRef]
- Sutherland, S.S.; Searson, J. The Immune Response of Brucella abortus: The Humoral Response. In Animal Brucellosis; Nielsen, K., Duncan, R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 65–81. [Google Scholar]
- Poester, F.P.; Samartino, L.E.; Santos, R.L. Pathogenesis and Pathobiology of Brucellosis in Livestock. Rev. Sci. Tech. l’OIE 2013, 32, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Enright, F.M. The Pathogenesis and Pathobiology of Brucella Infection in Domestic Animals. In Animal Brucellosis; Nielsen, K., Duncan, R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 301–320. [Google Scholar]
- Tohen, C.; Enright, F.M. Brucellosis. In Pathogenesis of Bacterial Lnfection in Animals; Gyles, C., Tohen, C., Eds.; Iowa State University Press: Ames, IA, USA, 1986. [Google Scholar]
- Bang, B. The Etiology of Epizootic Abortion. J. Comp. Pathol. Ther. 1897, 10, 125–149. [Google Scholar] [CrossRef]
- Anderson, T.D.; Meador, V.P.; Cheville, N.F. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. I. Gross and Histologic Lesions. Vet. Pathol. 1986, 23, 219–226. [Google Scholar] [CrossRef]
- Alexander, B.; Schnurrenberger, P.; Brown, R. Numbers of Brucella abortus in the Placenta, Umbilicus and Fetal Fluid of Two Naturally Infected Cows. Vet. Rec. 1981, 108, 500. [Google Scholar] [CrossRef]
- Smith, T. A Characteristic Localization of Bacillus abortus in the Bovine Fetal Membranes. J. Exp. Med. 1919, 29, 451–456. [Google Scholar] [CrossRef]
- Tobias, L.; Schurig, G.G.; Cordes, D.O. Comparative Behaviour of Brucella abortus Strains 19 and RB51 in the Pregnant Mouse. Res. Vet. Sci. 1992, 53, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Tobias, L.; Cordes, D.O.; Schurig, G.G. Placental Pathology of the Pregnant Mouse Inoculated with Brucella abortus Strain 2308. Vet. Pathol. 1993, 30, 119–129. [Google Scholar] [CrossRef]
- Kim, S.; Dong, S.L.; Watanabe, K.; Furuoka, H.; Suzuki, H.; Watarai, M. Interferon-γ Promotes Abortion Due to Brucella Infection in Pregnant Mice. BMC Microbiol. 2005, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, S.S.; Wang, G.L.; Wu, T.L.; Lv, Y.L.; Wu, Q.M. A Pregnant Mouse Model for the Vertical Transmission of Brucella melitensis. Vet. J. 2014, 200, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Byndloss, M.X.; Tsai, A.Y.; Walker, G.T.; Miller, C.N.; Young, B.M.; English, B.C.; Seyffert, N.; Kerrinnes, T.; de Jong, M.F.; Atluri, V.L.; et al. Brucella abortus Infection of Placental Trophoblasts Triggers Endoplasmic Reticulum Stress-Mediated Cell Death and Fetal Loss via Type IV Secretion System-Dependent Activation of CHOP. mBio 2019, 10, e01538-19. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M. Parasitization in Vitro of Bovine Cells by Brucella abortus. J. Bacteriol. 1959, 78, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; Terraza, A.; Ouahrani-Bettache, S.; Liautard, J.-P.; Dornand, J. In Vitro Brucella suis Infection Prevents the Programmed Cell Death of Human Monocytic Cells. Infect. Immun. 2000, 68, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Olarte, E.; Guzmán-Verri, C.; Méresse, S.; Desjardins, M.; Pizarro-Cerdá, J.; Badilla, J.; Gorvel, J.-P.; Moreno, E. Activation of Rho and Rab GTPases Dissociates Brucella abortus Internalization from Intracellular Trafficking. Cell Microbiol. 2002, 4, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Barquero-Calvo, E.; Moreno, E.; Guzmán-Verri, C.; Moriyón, I.; Rucavado, A.; Chacón-Díaz, C.; Weiss, D.S.; Chaves-Olarte, E. Brucella abortus Uses a Stealthy Strategy to Avoid Activation of the Innate Immune System during the Onset of Infection. PLoS ONE 2007, 2, e631. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Prada, C.M.; Zelazowska, E.B.; Nikolich, M.; Hadfield, T.L.; Roop, R.M.; Robertson, G.L.; Hoover, D.L. Interactions between Brucella melitensis and Human Phagocytes: Bacterial Surface O-Polysaccharide Inhibits Phagocytosis, Bacterial Killing, and Subsequent Host Cell Apoptosis. Infect. Immun. 2003, 71, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.M. The Pathogenesis of Experimental Brucellosis in the Pregnant Cow. J. Pathol. Bacteriol. 1959, 78, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Meador, V.P.; Deyoe, B.L. Experimentally Induced Brucella abortus Infection in Pregnant Goats. Am. J. Vet. Res. 1986, 47, 2337–2342. [Google Scholar]
- Caspe, S.G.; Ewing, D.A.; Livingstone, M.; Underwood, C.; Milne, E.; Sargison, N.D.; Wattegedera, S.R.; Longbottom, D. The Immune Response in the Uteri and Placentae of Chlamydia abortus-Infected Ewes and Its Association with Pregnancy Outcomes. Pathogens 2023, 12, 846. [Google Scholar] [CrossRef] [PubMed]
- Zavattieri, L.; Muñoz González, F.; Ferrero, M.C.; Baldi, P.C. Immune Responses Potentially Involved in the Gestational Complications of Brucella Infection. Pathogens 2023, 12, 1450. [Google Scholar] [CrossRef] [PubMed]
- Molello, J.A.; Jensen, R.; Flint, J.C.; Collier, J.R. Placental Pathology. I. Placental Lesions of Sheep Experimentally Infected with Brucella ovis. Am. J. Vet. Res. 1963, 24, 897–904. [Google Scholar]
- Molello, J.A.; Flint, J.C.; Collier, J.R.; Jensen, R. Placental Pathology. II. Placental Lesions of Sheep Experimentally Infected with Brucella melitensis. Am. J. Vet. Res. 1963, 24, 905–912. [Google Scholar] [PubMed]
- Molello, J.A.; Jensen, R.; Collier, J.R.; Flint, J.C. Placental Pathology. III. Placental Lesions of Sheep Experimentally Infected with Brucella abortus. Am. J. Vet. Res. 1963, 24, 915–922. [Google Scholar] [PubMed]
- Grilló, M.J.; Blasco, J.M.; Gorvel, J.P.; Moriyón, I.; Moreno, E. What Have We Learned from Brucellosis in the Mouse Model? Vet. Res. 2012, 43, 29. [Google Scholar] [CrossRef] [PubMed]
- Maltepe, E.; Fisher, S.J. Placenta: The Forgotten Organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef] [PubMed]
- Wooding, P.; Burton, G. Comparative Placentation: Structures, Functions and Evolution; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-78796-9. [Google Scholar]
- Geisert, R.D.; Spencer, T. Placentation in Mammals; Advances in Anatomy, Embryology and Cell Biology; Geisert, R.D., Spencer, T., Eds.; Springer International Publishing: Cham, Switzerland, 2021; Volume 234, ISBN 978-3-030-77359-5. [Google Scholar]
- Wooding, F.B.P. Ultrastructural Evidence for Placental Lactogen Transport and Secretion in Ruminants. J. Physiol. 1987, 386, 26. [Google Scholar]
- Elmore, S.A.; Cochran, R.Z.; Bolon, B.; Lubeck, B.; Mahler, B.; Sabio, D.; Ward, J.M. Histology Atlas of the Developing Mouse Placenta. Toxicol. Pathol. 2022, 50, 60–117. [Google Scholar] [CrossRef] [PubMed]
- Adamson, S.L.; Lu, Y.; Whiteley, K.J.; Holmyard, D.; Hemberger, M.; Pfarrer, C.; Cross, J.C. Interactions between Trophoblast Cells and the Maternal and Fetal Circulation in the Mouse Placenta. Dev. Biol. 2002, 250, 358–373. [Google Scholar] [CrossRef]
- Bosseray, N. Colonization of Mouse Placentas by Brucella abortus Inoculated during Pregnancy. Br. J. Exp. Pathol. 1980, 61, 361–368. [Google Scholar]
- Bosseray, N. Kinetics of Placental Colonization of Mice Inoculated Intravenously with Brucella abortus at Day 15 of Pregnancy. Br. J. Exp. Pathol. 1983, 64, 612–616. [Google Scholar] [PubMed]
- Bosseray, N. Mother to Young Transmission of Brucella abortus Infection in Mouse Model. Ann. Rech. Vet. 1982, 13, 341–349. [Google Scholar] [PubMed]
- Bosseray, N. Vaccine and Serum-Mediated Protection against Brucella Infection of Mouse Placenta. Br. J. Exp. Pathol. 1983, 64, 617–625. [Google Scholar] [PubMed]
- Bosseray, N.; Plommet, M. Serum- and Cell-Mediated Immune Protection of Mouse Placenta and Fetus against a Brucella abortus Challenge: Expression of Barrier Effect of Placenta. Placenta 1988, 9, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Iwai, N.; Tachibana, M.; Furuoka, H.; Suzuki, H.; Watarai, M. Regulated upon Activation Normal T-Cell Expressed and Secreted (RANTES) Contributes to Abortion Caused by Brucella abortus Infection in Pregnant Mice. J. Vet. Med. Sci. 2008, 70, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Tachibana, M.; Tanaka, S.; Furuoka, H.; Horiuchi, M.; Suzuki, H.; Watarai, M. Heat Shock Cognate Protein 70 Contributes to Brucella Invasion into Trophoblast Giant Cells That Cause Infectious Abortion. BMC Microbiol. 2008, 8, 212. [Google Scholar] [CrossRef] [PubMed]
- Hashino, M.; Kim, S.; Tachibana, M.; Shimizu, T.; Watarai, M. Vertical Transmission of Brucella abortus Causes Sterility in Pregnant Mice. J. Vet. Med. Sci. 2012, 74, 1075–1077. [Google Scholar] [CrossRef]
- Barbier, T.; Machelart, A.; Zúñiga-Ripa, A.; Plovier, H.; Hougardy, C.; Lobet, E.; Willemart, K.; Muraille, E.; De Bolle, X.; Van Schaftingen, E.; et al. Erythritol Availability in Bovine, Murine and Human Models Highlights a Potential Role for the Host Aldose Reductase during Brucella Infection. Front. Microbiol. 2017, 8, 1088. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, M.; Wu, J.; Wang, J.; Peng, Q. HMGB1 Release from Trophoblasts Contributes to Inflammation during Brucella melitensis Infection. Cell Microbiol. 2019, 21, e13080. [Google Scholar] [CrossRef] [PubMed]
- Keestra-Gounder, A.M.; Byndloss, M.X.; Seyffert, N.; Young, B.M.; Chávez-Arroyo, A.; Tsai, A.Y.; Cevallos, S.A.; Winter, M.G.; Pham, O.H.; Tiffany, C.R.; et al. NOD1 and NOD2 Signalling Links ER Stress with Inflammation. Nature 2016, 532, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.Y.; Byndloss, M.X.; Seyffert, N.; Winter, M.G.; Young, B.M.; Tsolis, R.M. Tumor Necrosis Factor Alpha Contributes to Inflammatory Pathology in the Placenta during Brucella abortus Infection. Infect. Immun. 2022, 90, e0001322. [Google Scholar] [CrossRef] [PubMed]
- Poveda-Urkixo, I.; Ramírez, G.A.; Grilló, M.-J. Kinetics of Placental Infection by Different Smooth Brucella Strains in Mice. Pathogens 2022, 11, 279. [Google Scholar] [CrossRef] [PubMed]
- Zabalza-Baranguá, A.; Poveda-Urkixo, I.; Mena-Bueno, S.; Ramírez, G.A.; De Bolle, X.; Grilló, M.J. Vaccine Properties of Brucella melitensis 16MΔwzm and Reactivation of Placental Infection in Pregnant Sheep. Vaccine 2023, 41, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Mena-Bueno, S.; Poveda-Urkixo, I.; Irazoki, O.; Palacios, L.; Cava, F.; Zabalza-Baranguá, A.; Grilló, M.J. Brucella melitensis Wzm/Wzt System: Changes in the Bacterial Envelope Lead to Improved Rev1Δwzm Vaccine Properties. Front. Microbiol. 2022, 13, 908495. [Google Scholar] [CrossRef] [PubMed]
- Braz, H.M.B.; Silva, M.F.; de Carvalho, T.P.; da Silva, L.A.; Soares, J.B.; Costa, F.B.; Sossai, B.G.; da Paixão, T.A.; Santos, R.L. Pathogenesis of Brucella ovis in Pregnant Mice and Protection Induced by the Candidate Vaccine Strain B. ovis ΔabcBA. Vaccine 2022, 40, 4617–4624. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, C.A.; Maurizio, E.; Rossi, U.A. Comparative Review of Brucellosis in Small Domestic Ruminants. Front. Vet. Sci. 2022, 9, 887671. [Google Scholar] [CrossRef] [PubMed]
- Letesson, J.-J.; Barbier, T.; Zúñiga-Ripa, A.; Godfroid, J.; De Bolle, X.; Moriyón, I. Brucella Genital Tropism: What’s on the Menu. Front. Microbiol. 2017, 8, 506. [Google Scholar] [CrossRef] [PubMed]
- Hecker, Y.P.; González-Ortega, S.; Cano, S.; Ortega-Mora, L.M.; Horcajo, P. Bovine Infectious Abortion: A Systematic Review and Meta-Analysis. Front. Vet. Sci. 2023, 10, 1249410. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasmosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, Twelfth Edition 2023; World Organisation for Animal Health: Paris, France, 2023; pp. 1–13. [Google Scholar]
- Schnee, D.; Borel, N.; Laroucau, K. Enzootic Abortion of Ewes (Ovine Chlamydiosis) (Infection with Chlamydia abortus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2023; pp. 1–10. [Google Scholar]
- Adetunji, S.A.; Faustman, D.L.; Adams, L.G.; Garcia-Gonzalez, D.G.; Hensel, M.E.; Khalaf, O.H.; Arenas-Gamboa, A.M. Brucella abortus and Pregnancy in Mice: Impact of Chronic Infection on Fertility and the Role of Regulatory T Cells in Tissue Colonization. Infect. Immun. 2020, 88, e00257-20. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Hou, H.; Zhao, W.; Wang, J.; Peng, Q. Administration of Exogenous Progesterone Protects Against Brucella abortus Infection–Induced Inflammation in Pregnant Mice. J. Infect. Dis. 2021, 224, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Bolon, B.; Ward, J. 4—Anatomy and Physiology of the Developing Mouse and Placenta. In Pathology of the Developing Mouse: A Systematic Approach; Bolon, B., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 39–98. [Google Scholar]
- Pang, S.; Janzen-Pang, J.; Tse, M.; Croy, B.; Lima, P. 1—The Cycling and Pregnant Mouse: Gross Anatomy. In The Guide to Investigation of Mouse Pregnancy; Croy, B., Yamada, A., DeMayo, F., Adamson, S., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 3–19. [Google Scholar]
- Robbins, J.R.; Bakardjiev, A.I. Pathogens and the Placental Fortress. Curr. Opin. Microbiol. 2012, 15, 36–43. [Google Scholar] [CrossRef]
- de Jong, M.F.; Tsolis, R.M. Brucellosis and Type IV Secretion. Future Microbiol. 2012, 7, 47–58. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, D.; Cazevieille, C.; Allardet-Servent, A.; Boschiroli, M.L.; Bourg, G.; Foulongne, V.; Frutos, P.; Kulakov, Y.; Ramuz, M. A Homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl Type IV Secretion Systems Is Essential for Intracellular Survival of Brucella suis. Mol. Microbiol. 1999, 33, 1210–1220. [Google Scholar] [CrossRef]
- Roux, C.M.; Rolán, H.G.; Santos, R.L.; Beremand, P.D.; Thomas, T.L.; Adams, L.G.; Tsolis, R.M. Brucella Requires a Functional Type IV Secretion System to Elicit Innate Immune Responses in Mice. Cell. Microbiol. 2007, 9, 1851–1869. [Google Scholar] [CrossRef] [PubMed]
- Sieira, R.; Comerci, D.J.; Sánchez, D.O.; Ugalde, R.A. A Homologue of an Operon Required for DNA Transfer in Agrobacterium Is Required in Brucella abortus for Virulence and Intracellular Multiplication. J. Bacteriol. 2000, 182, 4849–4855. [Google Scholar] [CrossRef] [PubMed]
- Celli, J. The Intracellular Life Cycle of Brucella spp. Microbiol. Spectr. 2019, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Renoux, G. Brucellosis in Goats and Sheep. In Advances in Veterinary Science; Brandly, C., Jungheer, E., Eds.; Academic Press: New York, NY, USA, 1957; Volume 3, pp. 241–273. [Google Scholar]
- Plommet, M.; Fensterbank, R.; Renoux, G.; Gestin, J.; Philippon, A. Brucellose Bovine Expérimentale. XII.—Persistance a l’âge Adulte de l’infection Congénitale de La Génisse. Ann. Rech. Vétérinaires 1973, 4, 419–435. [Google Scholar]
- Bosseray, N.; Plommet, M.; De Rycke, J. Development of Brucella abortus, Brucella melitensis and Brucella suis Infection Infection in Mice to a Chronic Stage and Cure. Ann. Rech. Vet. 1982, 13, 153–161. [Google Scholar] [PubMed]
- Lapraik, R.D.; Moffat, R. Latent Bovine Brucellosis. Vet. Rec. 1982, 111, 578–579. [Google Scholar]
- Renoux, G. Goat Brucellosis. II. Its Influence on Gestation and the Offspring. Ann. Zootech. 1962, 11, 61–76. [Google Scholar] [CrossRef]
- Grilló, M.J.; Barberán, M.; Blasco, J.M. Transmission of Brucella melitensis from Sheep to Lambs. Vet. Rec. 1997, 140, 602–605. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elizalde-Bielsa, A.; Muñoz, P.M.; Zúñiga-Ripa, A.; Conde-Álvarez, R. A Review on the Methodology and Use of the Pregnant Mouse Model in the Study of Brucella Reproductive Pathogenesis and Its Abortifacient Effect. Microorganisms 2024, 12, 866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050866
Elizalde-Bielsa A, Muñoz PM, Zúñiga-Ripa A, Conde-Álvarez R. A Review on the Methodology and Use of the Pregnant Mouse Model in the Study of Brucella Reproductive Pathogenesis and Its Abortifacient Effect. Microorganisms. 2024; 12(5):866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050866
Chicago/Turabian StyleElizalde-Bielsa, Aitor, Pilar M. Muñoz, Amaia Zúñiga-Ripa, and Raquel Conde-Álvarez. 2024. "A Review on the Methodology and Use of the Pregnant Mouse Model in the Study of Brucella Reproductive Pathogenesis and Its Abortifacient Effect" Microorganisms 12, no. 5: 866. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050866