

Multi-Omics of Campylobacter jejuni Growth in Chicken Exudate Reveals Molecular Remodelling Associated with Altered Virulence and Survival Phenotypes
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Experimental Design
2.3. Quantitative Proteomics by LC-MS/MS
2.4. Processing of Proteomics Mass Spectrometry Files
2.5. Targeted Metabolomics by LC-MS/MS
2.6. Biofilm Assays
2.7. Analysis of Lipid A by MALDI-TOF MS
2.8. Phenotypic Assays
3. Results
3.1. Proteomics of C. jejuni Response to Growth in 5% and 100% CE
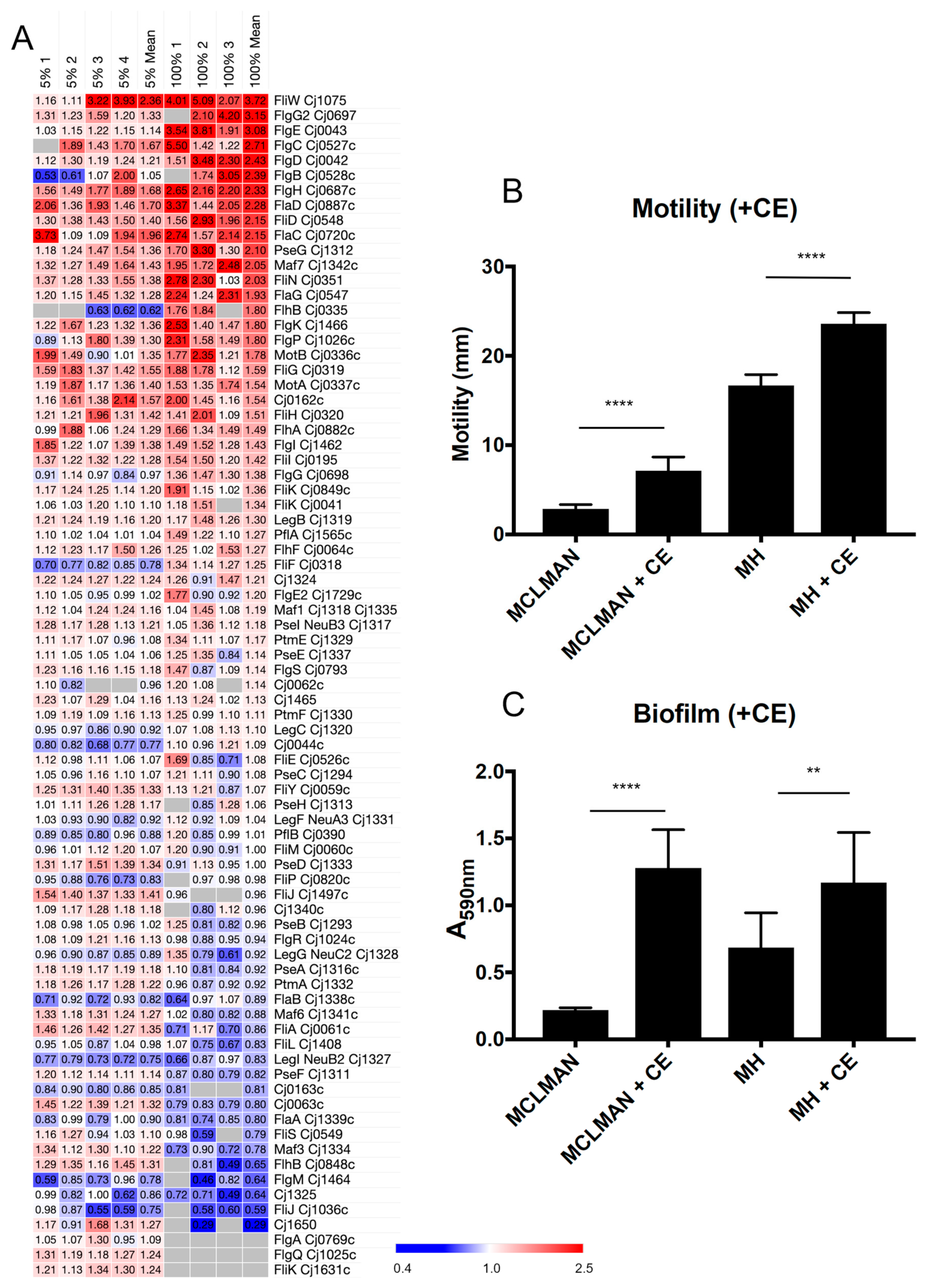
3.2. CE Increases C. jejuni Flagellar Motility and Induces Biofilm Formation
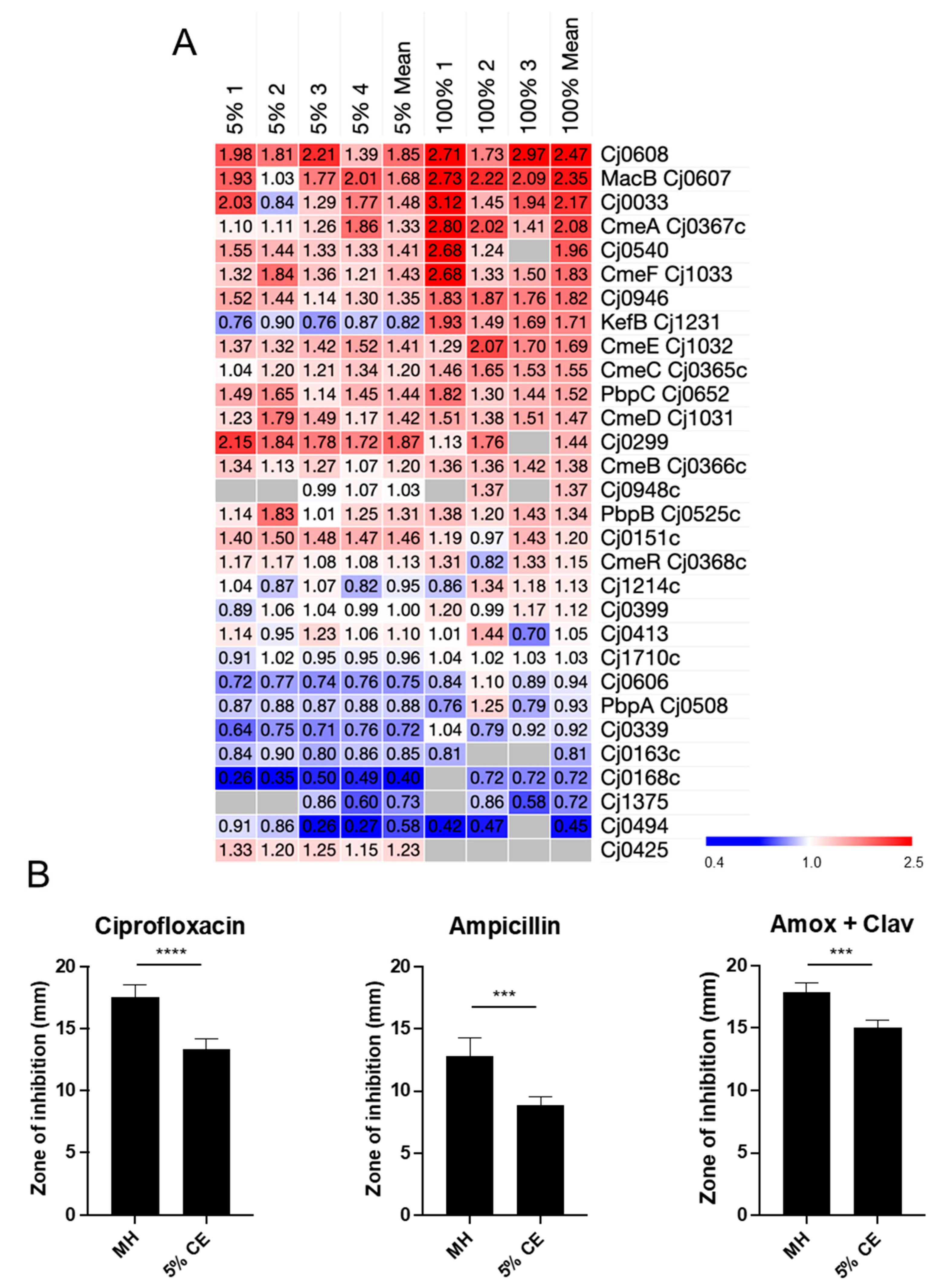
3.3. Growth in CE Leads to Increased C. jejuni Antibiotic Resistance
3.4. CE Induces Iron Uptake and Leads to Increased Oxidative Stress Resistance in C. jejuni
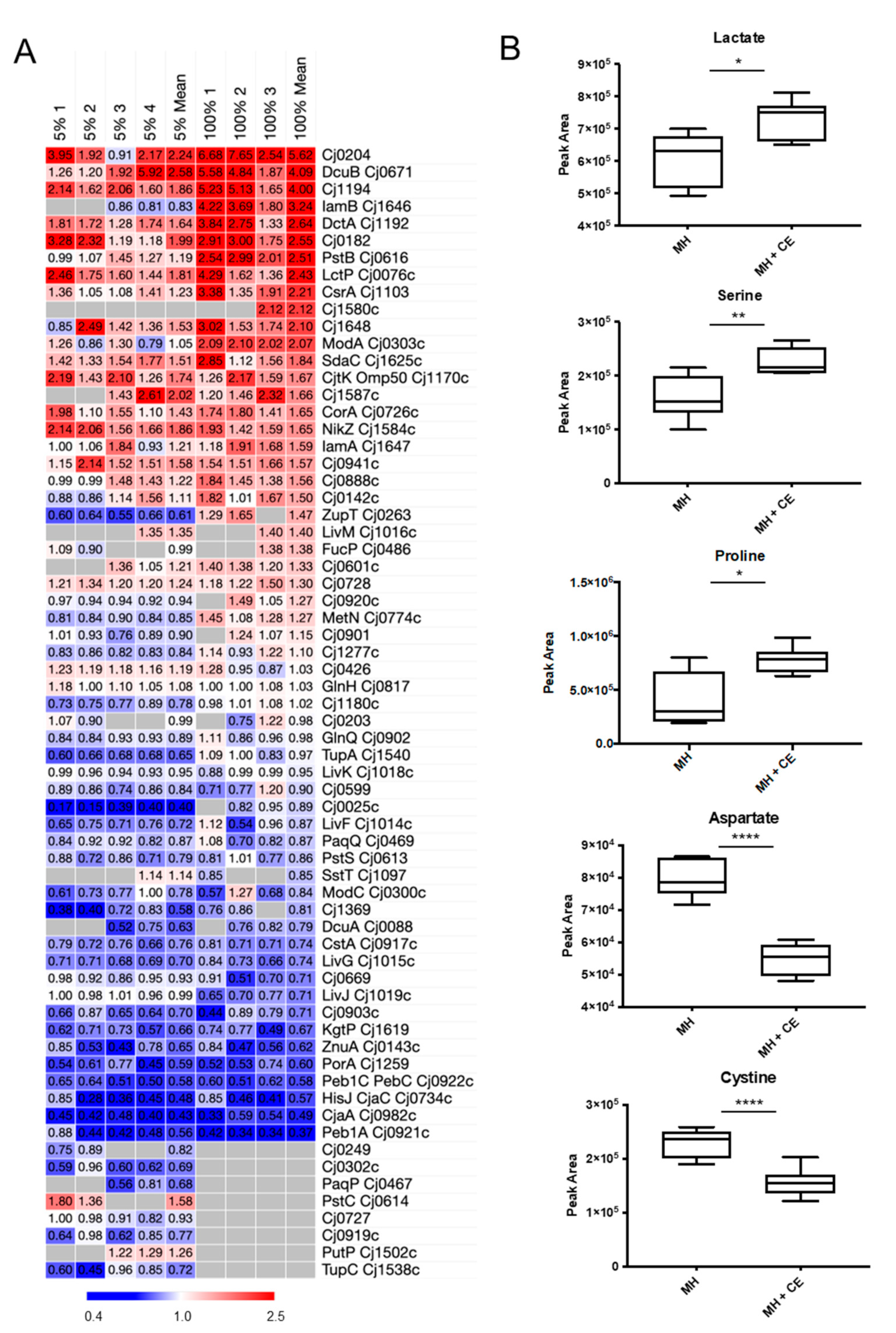
3.5. CE Results in Changes to C. jejuni Nutrient Transport
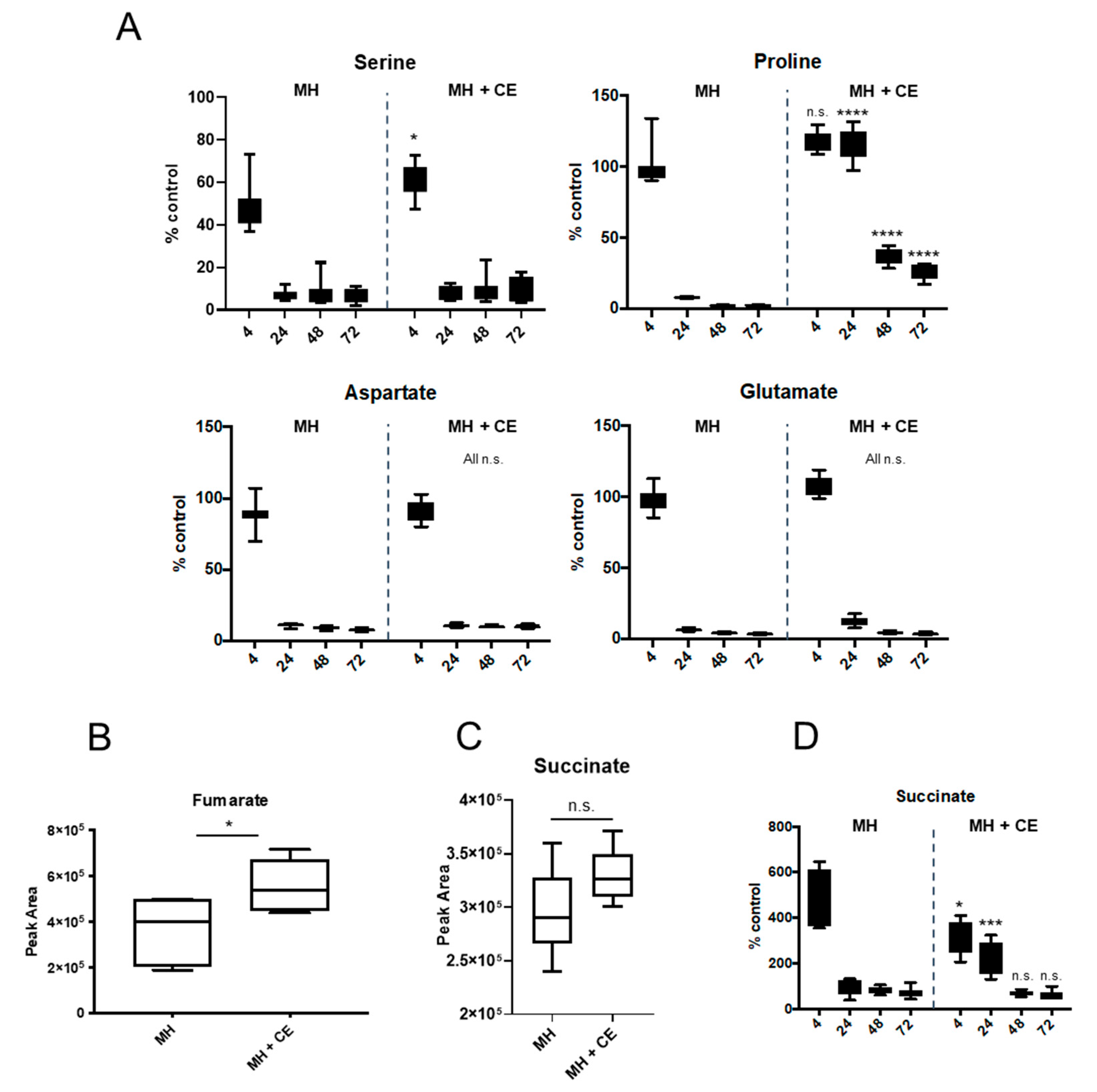
3.6. CE Alters C. jejuni Depletion of Specific Substrates from Culture Supernatants
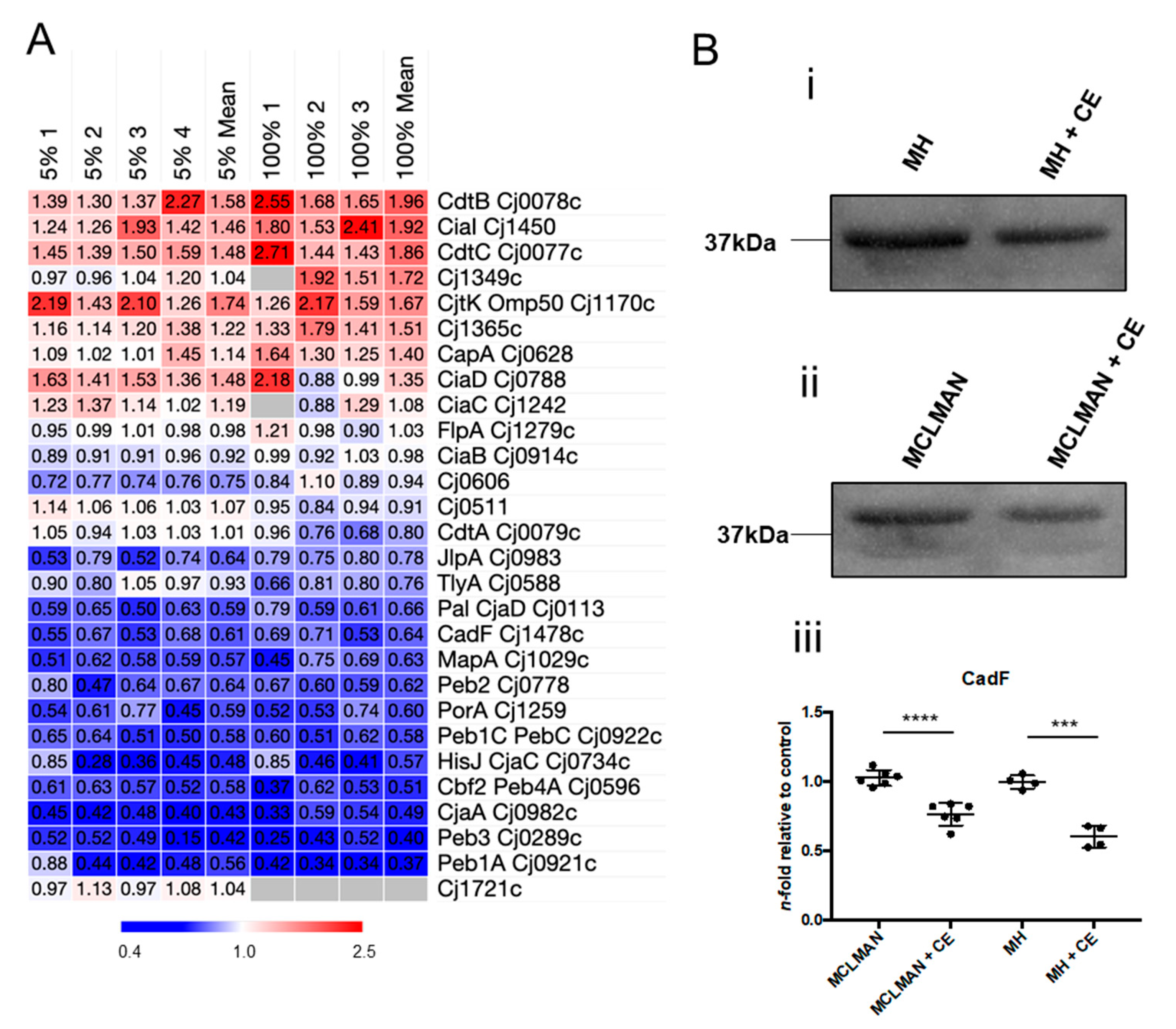
3.7. CE Alters Abundance of C. jejuni Virulence-Associated Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bryan, F.L.; Doyle, M.P. Health risks and consequences of Salmonella and Campylobacter jejuni in raw poultry. J. Food Prot. 1995, 58, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Al Hakeem, W.G.; Fathima, S.; Shanmugasundaram, R.; Selvaraj, R.K. Campylobacter jejuni in poultry: Pathogenesis and control strategies. Microorganisms 2022, 10, 2134. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.J.; Ferreira, V.; Truninger, M.; Maia, R.; Teixeira, P. Cross-contamination events of Campylobacter spp. in domestic kitchens associated with consumer handling of practices of raw poultry. Int. J. Food Microbiol. 2011, 338, 108984. [Google Scholar] [CrossRef]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter jejuni infections in humans. J. Infect. Dis. 1988, 157, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, D.; Thames, H.T.; Zhang, L.; Dinh, T.T.N.; Schilling, W.; White, S.B.; Ramachandran, R.; Theradiyil Sukumaran, A. Roles of aerotolerance, biofilm formation, and viable but non-culturable state in the survival of Campylobacter jejuni in poultry processing environments. Microorganisms 2022, 10, 2165. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. As a foodborne pathogen: A review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, N.; Sharafutdinov, I.; Harrer, A.; Soltan Esmaeili, D.; Linz, B.; Backert, S. Campylobacter virulence factors and molecular host-pathogen interactions. Curr. Top. Microbiol. Immunol. 2021, 431, 169–202. [Google Scholar]
- Lopes, G.V.; Ramires, T.; Kleinubing, N.R.; Scheik, L.K.; Fiorentini, Â.M.; Padilha da Silva, W. Virulence factors of foodborne pathogen Campylobacter jejuni. Microb. Pathog. 2021, 161 Pt A, 105265. [Google Scholar] [CrossRef]
- Korolik, V. The role of chemotaxis during Campylobacter jejuni colonization and pathogenesis. Curr. Opin. Microbiol. 2019, 47, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.A.; Sichel, S.R.; Salama, N.R. Bent bacteria: A comparison of cell shape mechanisms in proteobacteria. Annu. Rev. Microbiol. 2019, 73, 457–480. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Frirdich, E.; Vermeulen, J.; Badayeva, Y.; Li, X.; Vallance, B.A.; Gaynor, E.C. The helical cell shape of Campylobacter jejuni promotes in vivo pathogenesis by aiding transit through intestinal mucus and colonization of crypts. Infect. Immun. 2016, 84, 3399–3407. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Talukdar, P.K.; Negretti, N.M.; Klappenbach, C.M. Taking control: Campylobacter jejuni binding to fibronectin sets the stage for cellular adherence and invasion. Front. Microbiol. 2020, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Joe, A.; Lynett, J.; Hani, E.K.; Sherman, P.; Chan, V.L. JlpA, a novel surface-exposed lipoprotein specific to Campylobacter jejuni, mediates adherence to host epithelial cells. Mol. Microbiol. 2001, 39, 1225–1236. [Google Scholar] [PubMed]
- Konkel, M.E.; Klena, J.D.; Rivera-Amill, V.; Monteville, M.R.; Biswas, D.; Raphael, B.; Mickelson, J. Secretion of virulence proteins from Campylobacter jejuni is dependent on a functional flagellar export apparatus. J. Bacteriol. 2004, 186, 3296–3303. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Tobon, A.M.; Hendrixson, D.R. Identification and analysis of flagellar coexpressed determinants (Feds) of Campylobacter jejuni involved in colonization. Mol. Microbiol. 2012, 84, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Gabbert, A.D.; Mydosh, J.L.; Talukdar, P.K.; Gloss, L.M.; McDermott, J.E.; Cooper, K.K.; Clair, G.C.; Konkel, M.E. The missing pieces: The role of secretion systems in Campylobacter jejuni virulence. Biomolecules 2023, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, C.M. Bacterial glycosylation, it’s complicated. Front. Mol. Biosci. 2023, 9, 1015771. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, C.M.; Logan, S.M.; Linton, D.; Wren, B.W. Campylobacter—A tale of two protein glycosylation systems. Trends Microbiol. 2003, 11, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Guerry, P. Campylobacter flagella: Not just for motility. Trends Microbiol. 2007, 15, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.A.; Dale, A.L.; Sumer-Bayraktar, Z.; Solis, N.; Cordwell, S.J. Identifying the targets and functions of N-linked glycosylation in Campylobacter jejuni. Mol. Omics 2020, 16, 287–304. [Google Scholar] [CrossRef]
- Cain, J.A.; Dale, A.L.; Niewold, P.; Klare, W.P.; Man, L.; White, M.Y.; Scott, N.E.; Cordwell, S.J. Proteomics reveals multiple phenotypes associated with N-linked glycosylation in Campylobacter jejuni. Mol. Cell Proteom. 2019, 18, 715–734. [Google Scholar] [CrossRef] [PubMed]
- Abouelhadid, S.; Raynes, J.; Bui, T.; Cuccui, J.; Wren, B.W. Characterization of posttranslationally modified multidrug efflux pumps reveals an unexpected link between glycosylation and antimicrobial resistance. mBio 2020, 11, e02604-20. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.L.; Man, L.; Cordwell, S.J. Global acetylomics of Campylobacter jejuni shows lysine acetylation regulates CadF adhesin processing and human fibronectin binding. J. Proteome Res. 2023, 22, 3519–3533. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, L.C.; Craven, S.E. Campylobacter jejuni survival in chicken meat as a function of temperature. Appl. Environ. Microbiol. 1982, 44, 88–92. [Google Scholar] [CrossRef]
- Lee, A.; Smith, S.C.; Coloe, P.J. Survival and growth of Campylobacter jejuni after artificial inoculation onto chicken skin as a function of temperature and packaging conditions. J. Food Prot. 1998, 61, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Birk, T.; Ingmer, H.; Andersen, M.T.; Jørgensen, K.; Brøndsted, L. Chicken juice, a food-based model system suitable to study survival of Campylobacter jejuni. Lett. Appl. Microbiol. 2004, 38, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.T.; Mendonça, E.P.; Monteiro, G.P.; Siqueira, M.C.; Pereira, C.B.; Peres, P.A.B.M.; Fernandez, H.; Rossi, D.A. Intrinsic and extrinsic aspects on Campylobacter jejuni biofilms. Front. Microbiol. 2017, 8, 1332. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Friis, L.M.; Nothaft, H.; Liu, X.; Li, J.; Szymanski, C.M.; Stintzi, A. L-fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc. Natl. Acad. Sci. USA 2011, 108, 7194–7199. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, W.T.; Zhang, Q. Phenotypic and genotypic evidence for L-fucose utilization by Campylobacter jejuni. J. Bacteriol. 2011, 193, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Hofreuter, D. Defining the metabolic requirements for the growth and colonization capacity of Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2014, 4, 137. [Google Scholar] [CrossRef] [PubMed]
- Stahl, M.; Butcher, J.; Stintzi, A. Nutrient acquisition and metabolism by Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2012, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Velayudhan, J.; Jones, M.A.; Barrow, P.A.; Kelly, D.J. L-serine catabolism via an oxygen-labile L-serine dehydratase is essential for colonization of the avian gut by Campylobacter jejuni. Infect. Immun. 2004, 72, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hofreuter, D.; Mohr, J.; Wensel, O.; Rademacher, S.; Schreiber, K.; Schomburg, D.; Gao, B.; Galán, J.E. Contribution of amino acid catabolism to the tissue specific persistence of Campylobacter jejuni in a murine colonization model. PLoS ONE 2012, 7, e50699. [Google Scholar] [CrossRef] [PubMed]
- Del Rocio Leon-Kempis, M.; Guccione, E.; Mulholland, F.; Williamson, M.P.; Kelly, D.J. The Campylobacter jejuni PEB1a adhesin is an aspartate/glutamate-binding protein of an ABC transporter essential for microaerobic growth on dicarboxylic amino acids. Mol. Microbiol. 2006, 60, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Wösten, M.M.; van de Lest, C.H.; van Dijk, L.; van Putten, J.P. Function and regulation of the C4-dicarboxylate transporters in Campylobacter jejuni. Front. Microbiol. 2017, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.T.; Shepherd, M.; Poole, R.K.; van Vliet, A.H.M.; Kelly, D.J.; Pearson, B.M. Two respiratory enzyme systems in Campylobacter jejuni NCTC 11168 contribute to growth on L-lactate. Environ. Microbiol. 2011, 13, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; LeVeque, R.M.; Callahan, S.M.; Chatterjee, S.; Stopnisek, N.; Kuipel, M.; Johnson, J.G.; DiRita, V.J. Gut metabolite L-lactate supports Campylobacter jejuni population expansion during acute infection. Proc. Natl. Acad. Sci. USA 2024, 121, e2316540120. [Google Scholar] [CrossRef]
- Taylor, A.J.; Kelly, D.J. The function, biogenesis and regulation of the electron transport chains in Campylobacter jejuni: New insights into the bioenergetics of a major food-borne pathogen. Adv. Microb. Physiol. 2019, 74, 239–329. [Google Scholar] [PubMed]
- Sellars, M.J.; Hall, S.J.; Kelly, D.J. Growth of Campylobacter jejuni supported by respiration of fumarate, nitrate, nitrite, trimethylamine-N-oxide, or dimethyl sulfoxide requires oxygen. J. Bacteriol. 2002, 184, 4187–4196. [Google Scholar] [CrossRef]
- Guccione, E.; Hitchcock, A.; Hall, S.J.; Mulholland, F.; Shearer, N.; van Vliet, A.H.; Kelly, D.J. Reduction of fumarate, mesaconate and crotonate by Mfr, a novel oxygen-regulated periplasmic reductase in Campylobacter jejuni. Environ. Microbiol. 2010, 12, 576–591. [Google Scholar] [CrossRef]
- Weingarten, R.A.; Taveirne, M.E.; Olson, J.W. The dual-functioning fumarate reductase is the sole succinate:quinone reductase in Campylobacter jejuni and is required for full host colonization. J. Bacteriol. 2009, 191, 5293–5300. [Google Scholar] [CrossRef]
- Weerakoon, D.R.; Borden, N.J.; Goodson, C.M.; Grimes, J.; Olson, J.W. The role of respiratory donor enzymes in Campylobacter jejuni host colonization and physiology. Microb. Pathog. 2009, 47, 8–15. [Google Scholar] [CrossRef]
- Kim, J.C.; Oh, E.; Kim, J.; Jeon, B. Regulation of oxidative stress resistance in Campylobacter jejuni, a microaerophilic foodborne pathogen. Front. Microbiol. 2015, 6, 751. [Google Scholar] [CrossRef]
- Ligowska, M.; Thorup Cohn, M.; Stabler, R.A.; Wren, B.W.; Brøndsted, L. Effect of chicken meat environment on gene expression of Campylobacter jejuni and its relevance to survival in food. Int. J. Food Microbiol. 2011, 145 (Suppl. 1), S111–S115. [Google Scholar] [CrossRef]
- Brown, H.L.; Reuter, M.; Salt, L.J.; Cross, K.L.; Betts, R.P.; van Vliet, A.H. Chicken juice enhances surface attachment and biofilm formation of Campylobacter jejuni. Appl. Environ. Microbiol. 2014, 80, 7053–7060. [Google Scholar] [CrossRef]
- Man, L.; Klare, W.P.; Dale, A.L.; Cain, J.A.; Cordwell, S.J. Integrated mass spectrometry-based multi-omics for elucidating mechanisms of bacterial virulence. Biochem. Soc. Trans. 2021, 49, 1905–1926. [Google Scholar] [CrossRef]
- Man, L.; Dale, A.L.; Klare, W.P.; Cain, J.A.; Sumer-Bayraktar, Z.; Niewold, P.; Solis, N.; Cordwell, S.J. Proteomics of Campylobacter jejuni growth in deoxycholate reveals Cj0025c as a cystine transport protein required for wild-type human infection phenotypes. Mol. Cell. Proteom. 2020, 19, 1263–1280. [Google Scholar] [CrossRef]
- Alazzam, B.; Bonnassie-Rouxin, S.; Dufour, V.; Ermel, G. MCLMAN, a new minimal medium for Campylobacter jejuni NCTC 11168. Res. Microbiol. 2011, 162, 173–179. [Google Scholar] [CrossRef]
- Leung, L.M.; Fondrie, W.E.; Doi, Y.; Johnson, J.K.; Strickland, D.K.; Ernst, R.K.; Goodlett, D.R. Identification of the ESKAPE pathogens by mass spectrometric analysis of microbial membrane glycolipids. Sci. Rep. 2017, 7, 6403. [Google Scholar] [CrossRef]
- Gibb, S.; Strimmer, K. MALDIquant: A versatile R package for the analysis of mass spectrometry data. Bioinformatics 2012, 28, 2270–2271. [Google Scholar] [CrossRef]
- Scott, N.E.; Marzook, N.B.; Deutscher, A.; Falconer, L.; Crossett, B.; Djordjevic, S.P.; Cordwell, S.J. Mass spectrometric characterization of the Campylobacter jejuni adherence factor CadF reveals post-translational processing that removes immunogenicity while retaining fibronectin binding. Proteomics 2010, 10, 277–288. [Google Scholar] [CrossRef]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E. Polymyxin: Alternative mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef]
- Rasmussen, J.J.; Vegge, C.S.; Frøkiær, H.; Howlett, R.M.; Krogfelt, K.A.; Kelly, D.J.; Ingmer, H. Campylobacter jejuni carbon starvation protein A (CstA) is involved in peptide utilization, motility and agglutination, and has a role in stimulation of dendritic cells. J. Med. Microbiol. 2013, 62, 1135–1143. [Google Scholar] [CrossRef]
- Vorwerk, H.; Mohr, J.; Huber, C.; Wensel, O.; Schmidt-Hohagen, K.; Gripp, E.; Josenhans, C.; Schomburg, D.; Eisenreich, W.; Hofreuter, D. Utilization of host-derived cysteine-containing peptides overcomes the restricted sulphur metabolism of Campylobacter jejuni. Mol. Microbiol. 2014, 93, 1224–1245. [Google Scholar] [CrossRef]
- Hakeem, M.J.; Lu, X. Survival and control of Campylobacter in poultry production environment. Front. Cell. Infect. Microbiol. 2021, 10, 615049. [Google Scholar] [CrossRef]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and potential treatments for reducing Campylobacter colonization in animal hosts and disease in humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- Riedel, C.T.; Cohn, M.T.; Stabler, R.A.; Wren, B.; Brøndsted, L. Cellular response of Campylobacter jejuni to trisodium phosphate. Appl. Environ. Microbiol. 2012, 78, 1411–1415. [Google Scholar] [CrossRef]
- Oyarzabal, O.A. Reduction of Campylobacter spp. by commercial antimicrobials applied during the processing of broiler chickens: A review from the United States perspective. J. Food Prot. 2005, 68, 1752–1760. [Google Scholar] [CrossRef]
- Kalmokoff, M.; Lanthier, P.; Tremblay, T.L.; Foss, M.; Lau, P.C.; Sanders, G.; Austin, J.; Kelly, J.; Szymanski, C.M. Proteomic analysis of Campylobacter jejuni 11168 biofilms reveals a role for the motility complex in biofilm formation. J. Bacteriol. 2006, 188, 4312–4320. [Google Scholar] [CrossRef]
- Püning, C.; Su, Y.; Lu, X.; Gölz, G. Molecular mechanisms of Campylobacter jejuni biofilm formation and quorum sensing. Curr. Top. Microbiol. Immunol. 2021, 431, 293–319. [Google Scholar]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef] [PubMed]
- Hendrixson, D.R.; DiRita, V.J. Identification of Campylobacter jejuni genes involved in commensal colonization of the chick gastrointestinal tract. Mol. Microbiol. 2004, 52, 471–484. [Google Scholar] [CrossRef]
- Li, J.; Gulbronson, C.J.; Bogacz, M.; Hendrixson, D.R.; Thompson, S.A. FliW controls growth-phase expression of Campylobacter jejuni flagella and non-flagellar proteins via the post-transcriptional regulator CsrA. Microbiology 2018, 164, 1308–1319. [Google Scholar] [CrossRef]
- Dugar, G.; Svensson, S.L.; Bischler, T.; Wäldchen, S.; Reinhardt, R.; Sauer, M.; Sharma, C.M. The CsrA-FliW network controls polar localization of the dual-function flagellin mRNA in Campylobacter jejuni. Nat. Commun. 2016, 7, 11667. [Google Scholar] [CrossRef]
- Fields, J.A.; Thompson, S.A. Campylobacter jejuni CsrA complements an Escherichia coli csrA mutation for the regulation of biofilm formation, motility and cellular morphology but not glycogen accumulation. BMC Microbiol. 2012, 12, 233. [Google Scholar] [CrossRef]
- Cox, C.A.; Bogacz, M.; El Abbar, F.M.; Browning, D.D.; Hsueh, B.Y.; Waters, C.M.; Lee, V.T.; Thompson, S.A. The Campylobacter jejuni response regulator and cyclic-di-GMP binding CbrR is a novel regulator of flagellar motility. Microorganisms 2021, 10, 86. [Google Scholar] [CrossRef]
- Elgamoudi, B.A.; Starr, K.S.; Korolik, V. Extracellular c-di-GMP plays a role in biofilm formation and dispersion of Campylobacter jejuni. Microorganisms 2022, 10, 2030. [Google Scholar] [CrossRef]
- Reuter, M.; Mallett, A.; Pearson, B.M.; van Vliet, A.H. Biofilm formation by Campylobacter jejuni is increased under aerobic conditions. Appl. Environ. Microbiol. 2010, 76, 2122–2128. [Google Scholar] [CrossRef]
- Kassem, I.I.; Candelero-Rueda, R.A.; Esseili, K.A.; Rajashekara, G. Formate simultaneously reduces oxidase activity and enhances respiration in Campylobacter jejuni. Sci. Rep. 2017, 7, 40117. [Google Scholar] [CrossRef]
- Feng, J.; Ma, L.; Nie, J.; Konkel, M.E.; Lu, X. Environmental stress-induced bacterial lysis and extracellular DNA release contribute to Campylobacter jejuni biofilm formation. Appl. Environ. Microbiol. 2018, 84, e02068-17. [Google Scholar] [CrossRef]
- Shen, Z.; Wang, Y.; Zhang, Q.; Shen, J. Antimicrobial resistance in Campylobacter spp. Microbiol. Spectr. 2018, 6, 2. [Google Scholar] [CrossRef]
- Alfredson, D.A.; Korolik, V. Isolation and expression of a novel molecular class D β-lactamase, OXA-61, from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2515–2518. [Google Scholar] [CrossRef]
- Zeng, X.; Brown, S.; Gillespie, B.; Lin, J. A single nucleotide in the promoter region modulates the expression of the β-lactamase OXA-61 in Campylobacter jejuni. J. Antimicrob. Chemother. 2014, 69, 1215–1223. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in Gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef]
- Van Mourik, A.; Steeghs, L.; van Laar, J.; Meiring, H.D.; Hamstra, H.J.; van Putten, J.P.; Wösten, M.M. Altered linkage of hydroxyacyl chains in lipid A of Campylobacter jejuni reduces TLR4 activation and antimicrobial resistance. J. Biol. Chem. 2010, 285, 15828–15836. [Google Scholar] [CrossRef]
- Mettert, E.L.; Kiley, P.J. How is Fe-S cluster formation regulated? Annu. Rev. Microbiol. 2015, 69, 505–526. [Google Scholar] [CrossRef]
- Van Vliet, A.H.M.; Ketley, J.M.; Park, S.F.; Penn, C.W. The role of iron in Campylobacter gene regulation, metabolism and oxidative stress defense. FEMS Microbiol. Rev. 2002, 26, 173–186. [Google Scholar] [CrossRef]
- Van Vliet, A.H.; Baillon, M.L.; Penn, C.W.; Ketley, J.M. Campylobacter jejuni contains two fur homologs: Characterization of iron-responsive regulation of peroxide stress defense genes by the PerR repressor. J. Bacteriol. 1999, 181, 6371–6376. [Google Scholar] [CrossRef]
- Van der Stel, A.X.; van de Lest, C.H.A.; Huynh, S.; Parker, C.T.; van Putten, J.P.M.; Wösten, M.M.S.M. Catabolite repression in Campylobacter jejuni correlates with intracellular succinate levels. Environ. Microbiol. 2018, 20, 1374–1388. [Google Scholar] [CrossRef] [PubMed]
- Kassem, I.I.; Khatri, M.; Sanad, Y.M.; Wolboldt, M.; Saif, Y.M.; Olson, J.W.; Rajashekara, G. The impairment of methylmenaquinol:fumarate reductase affects hydrogen peroxide susceptibility and accumulation in Campylobacter jejuni. Microbiologyopen 2014, 3, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.C.; Baek, K.H.; Lee, Y.E.; Kang, T.; Kim, H.J.; Lee, D.; Jo, C. Using 2D qNMR analysis to distinguish between frozen and frozen/thawed chicken meat and evaluate freshness. NPJ Sci. Food. 2022, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Dragsted, L.O. Biomarkers of meat intake and the application of nutrigenomics. Meat Sci. 2010, 84, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Composition of amino acids and related nitrogenous nutrients in feedstuffs for animal diets. Amino Acids 2020, 52, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Banerjee, S.; Vogel, H.J. Ligand binding specificity of the Escherichia coli periplasmic histidine binding protein, HisJ. Protein Sci. 2017, 26, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Kemper, L.; Hensel, A. Campylobacter jejuni: Targeting host cells, adhesion, invasion, and survival. Appl. Microbiol. Biotechnol. 2023, 107, 2725–2754. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Tobon, A.M.; Hendrixson, D.R. Flagellar biosynthesis exerts temporal regulation of secretion of specific Campylobacter jejuni colonization and virulence determinants. Mol. Microbiol. 2014, 93, 957–974. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gharaibeh, R.Z.; Newsome, R.C.; Pope, J.L.; Dougherty, M.W.; Tomkovich, S.; Pons, B.; Mirey, G.; Vignard, J.; Hendrixson, D.R.; et al. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut 2019, 68, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Morrison, M.J.; Imperiali, B. The renaissance of bacillosamine and its derivatives: Pathway characterization and implications in pathogenicity. Biochemistry 2014, 53, 624–638. [Google Scholar] [CrossRef]
- Cain, J.A.; Dale, A.L.; Cordwell, S.J. Exploiting pglB oligosaccharyltransferase-positive and -negative Campylobacter jejuni and a multiprotease digestion strategy to identify novel sites modified by N-linked protein glycosylation. J. Proteome Res. 2021, 20, 4995–5009. [Google Scholar] [CrossRef] [PubMed]
- Nothaft, H.; Liu, X.; Li, J.; Szymanski, C.M. Campylobacter jejuni free oligosaccharides: Function and fate. Virulence 2010, 1, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Frirdich, E.; Vermeulen, J.; Biboy, J.; Vollmer, W.; Gaynor, E.C. Multiple Campylobacter jejuni proteins affecting the peptidoglycan structure and the degree of helical cell curvature. Front. Microbiol. 2023, 14, 1162806. [Google Scholar] [CrossRef] [PubMed]
- Ha, R.; Frirdich, E.; Sychantha, D.; Biboy, J.; Taveirne, M.E.; Johnson, J.G.; DiRita, V.J.; Vollmer, W.; Clarke, A.J.; Gaynor, E.C. Accumulation of peptidoglycan O-acetylation leads to altered cell wall biochemistry and negatively impacts pathogenesis factors of Campylobacter jejuni. J. Biol. Chem. 2016, 291, 22686–22702. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Mathure, S.A.; Lee, M.; Boorman, J.; Zeng, X.; Lin, J.; Hesek, D.; Lastochkin, E.; Mobashery, S.; van den Akker, F. Turnover chemistry and structural characterization of the Cj0843c lytic transglycosylase of Campylobacter jejuni. Biochemistry 2021, 60, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Dik, D.A.; Marous, D.R.; Fisher, J.F.; Mobashery, S. Lytic transglycosylases: Concinnity in concision of the bacterial cell wall. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 503–542. [Google Scholar] [CrossRef]
- Zhang, X.L.; Báti, G.; Li, C.; Guo, A.; Yeo, C.; Ding, H.; Pal, K.B.; Xu, Y.; Qiao, Y.; Liu, X.W. GlcNAc-1,6-anhydro-MurNAc moiety affords unusual glycosyl acceptor that terminates peptidoglycan elongation. J. Am. Chem. Soc. 2024, 146, 7400–7407. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Man, L.; Soh, P.X.Y.; McEnearney, T.E.; Cain, J.A.; Dale, A.L.; Cordwell, S.J. Multi-Omics of Campylobacter jejuni Growth in Chicken Exudate Reveals Molecular Remodelling Associated with Altered Virulence and Survival Phenotypes. Microorganisms 2024, 12, 860. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050860
Man L, Soh PXY, McEnearney TE, Cain JA, Dale AL, Cordwell SJ. Multi-Omics of Campylobacter jejuni Growth in Chicken Exudate Reveals Molecular Remodelling Associated with Altered Virulence and Survival Phenotypes. Microorganisms. 2024; 12(5):860. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050860
Chicago/Turabian StyleMan, Lok, Pamela X. Y. Soh, Tess E. McEnearney, Joel A. Cain, Ashleigh L. Dale, and Stuart J. Cordwell. 2024. "Multi-Omics of Campylobacter jejuni Growth in Chicken Exudate Reveals Molecular Remodelling Associated with Altered Virulence and Survival Phenotypes" Microorganisms 12, no. 5: 860. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050860