
Immune-Related Gene Profiles and Differential Expression in the Grey Garden Slug Deroceras reticulatum Infected with the Parasitic Nematode Phasmarhabditis hermaphrodita
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
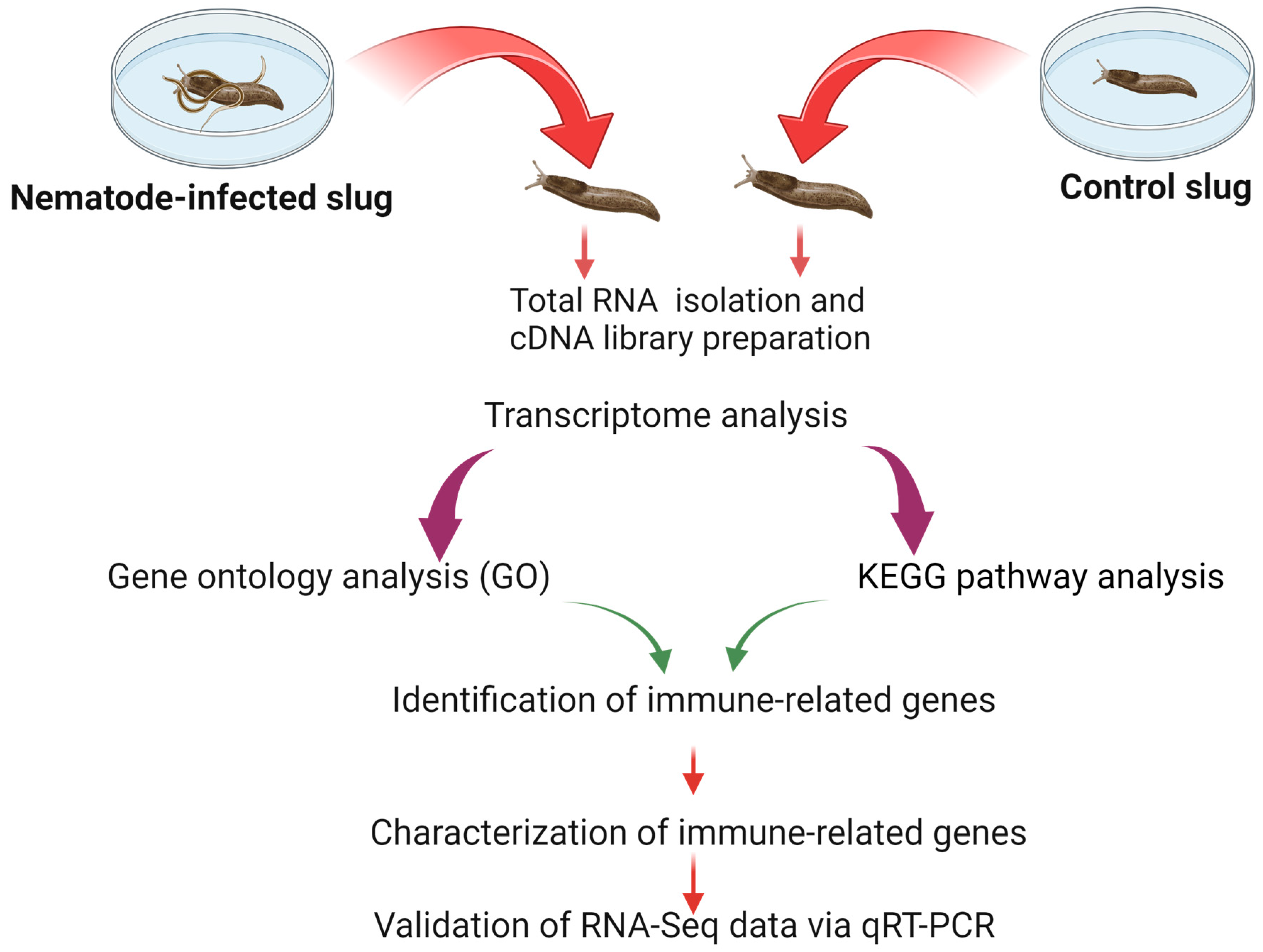
2.1. Slugs and Infection of the Nematodes
2.2. Total RNA Preparation
2.3. cDNA Library Preparation and RNA Illumina Sequencing
2.4. De Novo Transcriptome Assembly
2.5. Functional Annotation
2.6. Identification of Immunity-Related Genes
2.7. Validation of DEGs Results by qRT-PCR
2.8. Statistical Analysis
3. Results
3.1. Illumina Sequencing and De Novo Assembly
3.2. Functional Annotation of the Control Slug and Nematode-Infected Slug Unigenes
3.3. Differentially Expressed Genes in the Nematode-Infected Slug
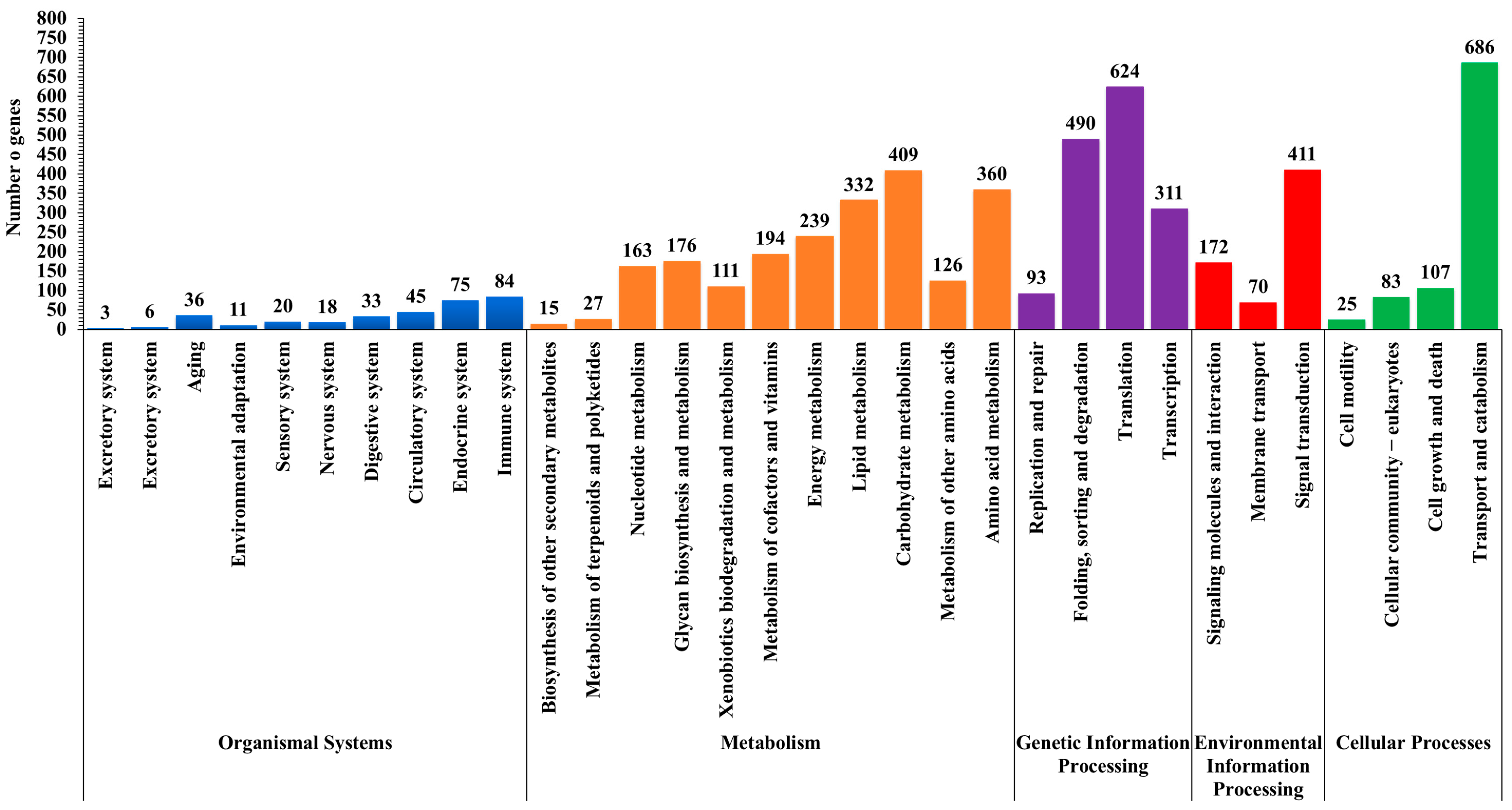
3.4. GO and KEGG Functional Classification of Nematode-Infected Slug Unigenes
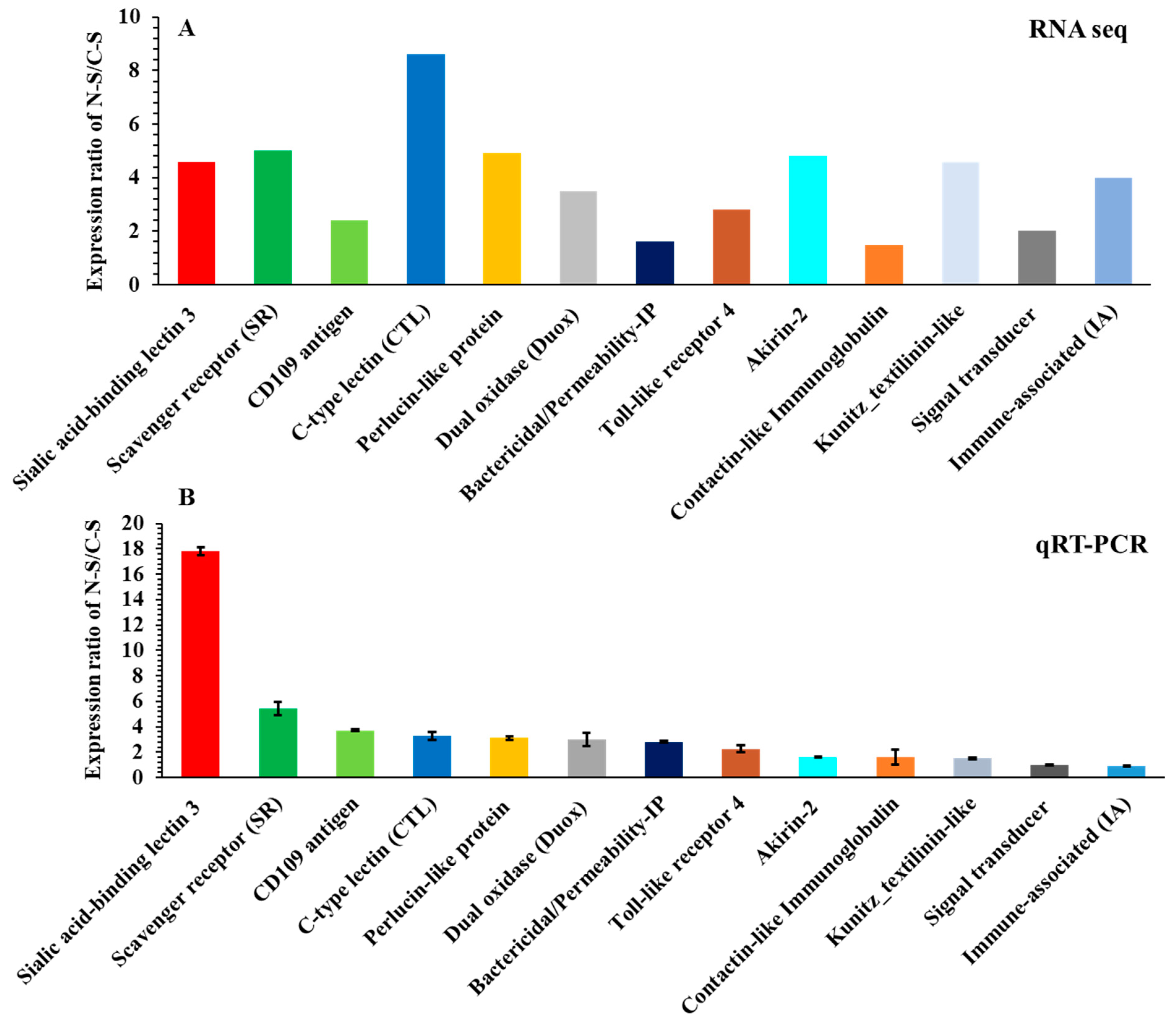
3.5. Identification and Expression Pattern of Immunity-Related Genes in Response to Pathogenic Nematode Infection
3.6. Validation of Differentially Expressed Immunity-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- South, A. Terrestrial Slugs: Biology, Ecology and Control, 1st ed.; Springer: Singapore, 1992. [Google Scholar] [CrossRef]
- Godan, D. Pest Slugs and Snails: Biology and Control; Springer: Berlin, Germany, 1983. [Google Scholar]
- Maheshini, P.W.D.B.; Thilakarathne, K.G.D.D.; Hirimuthugoda, G.N.; Ranawana, K.B.; Kumburegama, S. The Distribution of Terrestrial Pest Gastropods and Their Damage to Agricultural Crops in Kandy and Nuwara Eliya Districts in Sri Lanka. Ceylon J. Sci. 2019, 48, 177–184. [Google Scholar] [CrossRef]
- Wilson, M.J.; Glen, D.M.; George, S.K. The Rhabditid Nematode Phasmarhabditis Hermaphrodita as a Potential Biological Control Agent for Slugs. Biocontrol Sci. Technol. 1993, 3, 503–511. [Google Scholar] [CrossRef]
- Willis, J.C.; Bohan, D.A.; Choi, Y.H.; Conrad, K.F.; Semenov, M.A. Use of an Individual-Based Model to Forecast the Effect of Climate Change on the Dynamics, Abundance and Geographical Range of the Pest Slug Deroceras Reticulatum in the UK. Glob. Chang. Biol. 2006, 12, 643–1657. [Google Scholar] [CrossRef]
- Port, G.; Ester, A. Gastropods as Pests in Vegetable and Ornamental Crops in Western Europe. In Molluscs as Crop Pests; CABI Publishing: Wallingford, UK, 2002; pp. 337–351. [Google Scholar]
- Hommay, G. The Slugs Injurious to Crops. Rev. Suisse d’Agric. 1995, 27, 267–286. [Google Scholar]
- Barua, A.; Williams, C.D.; Ross, J.L. A Literature Review of Biological and Bio-Rational Control Strategies for Slugs: Current Research and Future Prospects. Insects 2021, 12, 541. [Google Scholar] [CrossRef] [PubMed]
- Rae, R.; Sheehy, L.; McDonald-Howard, K. Thirty Years of Slug Control Using the Parasitic Nematode Phasmarhabditis Hermaphrodita and Beyond. Pest Manag. Sci. 2023, 79, 3408–3424. [Google Scholar] [CrossRef] [PubMed]
- Hladik, M.L.; Main, A.R.; Goulson, D. Environmental Risks and Challenges Associated with Neonicotinoid Insecticides. Environ. Sci. Technol. 2018, 52, 3329–3335. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.M.; Verma, V.K.; Rawat, B.S.; Kaur, B.; Babu, N.; Sharma, A.; Dewali, S.; Yadav, M.; Kumari, R.; Singh, S.; et al. Current Status of Pesticide Effects on Environment, Human Health and It’s Eco-Friendly Management as Bioremediation: A Comprehensive Review. Front. Microbiol. 2022, 13, 962619. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, A.; Hafeez, F.; Hafeez, M.; Farooq, M.; Aziz, M.A.; Sohaib, M.; Naeem, A.; Lu, Y. Correction to: Sublethal Effects of a Juvenile Hormone Analog, Pyriproxyfen Demographic Parameters of Non-Target Predator, Hippodamia convergens Guerin-Meneville (Coleoptera: Coccinellidae). Ecotoxicology 2020, 29, 1017–1028. [Google Scholar] [PubMed]
- Al-Khalaifah, H. Cellular and Humoral Immune Response between Snail Hosts and Their Parasites. Front. Immunol. 2022, 13, 981314. [Google Scholar] [CrossRef] [PubMed]
- Viljakainen, L. Evolutionary Genetics of Insect Innate Immunity. Brief. Funct. Genom. 2015, 14, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Farrell, G.; Kavanagh, K. Immune Priming: The Secret Weapon of the Insect World. Virulence 2020, 11, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, S.; De Mandal, S.; Gao, Y.; Yu, J.; Zeng, L.; Huang, J.; Zafar, J.; Jin, F.; Xu, X. Combined Transcriptomic and Proteomic Analysis of Developmental Features in the Immune System of Plutella Xylostella during Larva-to-Adult Metamorphosis. Genomics 2022, 114, 110381. [Google Scholar] [CrossRef]
- Wu, G.; Yi, Y. Transcriptome Analysis of Differentially Expressed Genes Involved in Innate Immunity following Bacillus Thuringiensis Challenge in Bombyx Mori Larvae. Mol. Immunol. 2018, 103, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Altincicek, B.; Glöckner, G.; Vilcinskas, A. A Comprehensive Transcriptome and Immune-Gene Repertoire of the Lepidopteran Model Host Galleria Mellonella. BMC Genom. 2011, 12, 308. [Google Scholar] [CrossRef] [PubMed]
- Patnaik, B.B.; Chung, J.M.; Hwang, H.J.; Sang, M.K.; Park, J.E.; Min, H.R.; Cho, H.C.; Dewangan, N.; Baliarsingh, S.; Kang, S.W.; et al. Transcriptome Analysis of Air-Breathing Land Slug, Incilaria Fruhstorferi Reveals Functional Insights into Growth, Immunity, and Reproduction. BMC Genom. 2019, 154, 20. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, N.; Gao, X.; Guo, D.; Chang, Z.; Fu, Y.; Akinyemi, I.A.; Wu, Q. Understanding the Immune System Architecture and Transcriptome Responses to Southern Rice Black-Streaked Dwarf Virus in Sogatella Furcifera. Sci. Rep. 2016, 6, 36254. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.J.; Martin, R.; Rao, S.; Choi, M.Y. Neuropeptides Predicted from the Transcriptome Analysis of the Gray Garden Slug Deroceras Reticulatum. Peptides 2017, 93, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Xu, X.; Xu, J.; Zhu, X.; Li, S.; Zhou, X.; Yu, J.; Xu, X.; Hu, Q.; Yu, X.; et al. Identification of Immunity-Related Genes in Plutella Xylostella in Response to Fungal Peptide Destruxin A: RNA-Seq and DGE Analysis. Sci. Rep. 2017, 7, 10966. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Garg, S.; Gupta, L.; Gupta, K.; Diagne, C.T.; Missé, D.; Pompon, J.; Kumar, S.; Saxena, V. Delineating the Role of Aedes Aegypti Abc Transporter Gene Family during Mosquito Development and Arboviral Infection via Transcriptome Analyses. Pathogens 2021, 10, 1127. [Google Scholar] [CrossRef] [PubMed]
- Grewal, P.S.; Grewal, S.K.; Tan, L.; Adams, B.J. Parasitism of Molluscs by Nematodes: Types of Associations and Evolutionary Trends. J. Nematol. 2003, 35, 146–156. [Google Scholar] [PubMed]
- Pieterse, A.; Malan, A.P.; Ross, J.L. Nematodes That Associate with Terrestrial Molluscs as Definitive Hosts, Including Phasmarhabditis hermaphrodita (Rhabditida: Rhabditidae) and Its Development as a Biological Molluscicide. J. Helminthol. 2017, 91, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Thiengo, S.C.; Ramos-de-Souza, J.; Silva, G.M.; Fernandez, M.A.; Silva, E.F.; Sousa, A.K.P.; Rodrigues, P.S.; Mattos, A.C.; Costa, R.A.F.; Gomes, S.R. Parasitism of Terrestrial Gastropods by Medically-Important Nematodes in Brazil. Front. Vet. Sci. 2022, 9, 1023426. [Google Scholar] [CrossRef] [PubMed]
- Mc Donnell, R.J.; Colton, A.J.; Howe, D.K.; Denver, D.R. Lethality of Four Species of Phasmarhabditis (Nematoda: Rhabditidae) to the Invasive Slug, Deroceras reticulatum (Gastropoda: Agriolimacidae) in Laboratory Infectivity Trials. Biol. Control 2020, 150, 104349. [Google Scholar] [CrossRef]
- McDonnell, R.J.; Howe, D.K.; Denver, D.R. First Report of the Gastropod-Killing Nematode, Phasmarhabditis Californica, in Washington State, U.S.A. J. Nematol. 2023, 55, e2023-1. [Google Scholar] [CrossRef] [PubMed]
- Brophy, T.; Howe, D.K.; Denver, D.R.; Luong, L.T. First Report of a Gastropod Parasitic Nematode Phasmarhabditis californica (Nematoda: Rhabditidae) in Alberta, Canada. J. Nematol. 2020, 52, 1–3. [Google Scholar] [CrossRef]
- McDonnell, R.J.; Lutz, M.S.; Howe, D.K.; Denver, D.R. First Report of the Gastropod-Killing Nematode, Phasmarhabditis Hermaphrodita, in Oregon, U.S.A. J. Nematol. 2018, 50, 77–78. [Google Scholar] [CrossRef] [PubMed]
- De Ley, I.T.; Holovachov, O.; McDonnell, R.J.; Bert, W.; Paine, T.D.; De Ley, P. Description of Phasmarhabditis californica n. sp. and First Report of P. papillosa (Nematoda: Rhabditidae) from Invasive Slugs in the USA. Nematology 2016, 18, 175–193. [Google Scholar] [CrossRef]
- Lopez, S.; Paine, T.D.; De Ley, P.; De Ley, I.T.; McDonnell, R.D. Phasmarhabditis hermaphrodita (Nematoda: Rhabditidae), a Potential Biocontrol Agent Isolated for the First Time from Invasive Slugs in North America. Nematology 2014, 16, 1129–1138. [Google Scholar] [CrossRef]
- Tan, L.; Grewal, P.S. Infection Behavior of the Rhabditid Nematode Phasmarhabditis Hermaphrodita to the Grey Garden Slug Deroceras Reticulatum. J. Parasitol. 2001, 87, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Mc Donnell, R.J.; Paine, T.D.; Gormally, M.J. Slugs: A Guide to the Invasive and Native Fauna of California; UCANR Publications: Davis, CA, USA, 2009. [Google Scholar]
- Vlach, J. Slugs and Snails in Oregon; Oregon Department of Agriculture Guides: Salem, OR, USA, 2016. [Google Scholar]
- Donnell, R.M.; Colton, A.; Howe, D.; Denver, D. Susceptibility of Testacella haliotidea (Testacellidae: Mollusca) to a U.S. Strain of Phasmarhabditis hermaphrodita (Rhabditidae: Nematoda). Biocontrol Sci. Technol. 2022, 32, 262–266. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.A.; Christie, K.R.; Dolan, M.E.; Drabkin, H.J.; Hill, D.P.; Ni, L.; Sitnikov, D.; Burgess, S.; Buza, T.; Gresham, C.; et al. Gene Ontology Consortium: Going Forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Eleftherianos, I. Parasitic Nematode Immunomodulatory Strategies: Recent Advances and Perspectives. Pathogens 2016, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Bobardt, S.D.; Dillman, A.R.; Nair, M.G. The Two Faces of Nematode Infection: Virulence and Immunomodulatory Molecules From Nematode Parasites of Mammals, Insects and Plants. Front. Microbiol. 2020, 11, 577846. [Google Scholar] [CrossRef] [PubMed]
- Kallali, N.S.; Ouijja, A.; Goura, K.; Laasli, S.-E.; Kenfaoui, J.; Benseddik, Y.; Blenzar, A.; Joutei, A.B.; El Jarroudi, M.; Mokrini, F.; et al. From Soil to Host: Discovering the Tripartite Interactions between Entomopathogenic Nematodes, Symbiotic Bacteria and Insect Pests and Related Challenges. J. Nat. Pestic. Res. 2023, 7, 100065. [Google Scholar] [CrossRef]
- Dziarski, R.; Gupta, D. How Innate Immunity Proteins Kill Bacteria and Why They Are Not Prone to Resistance. Curr. Genet. 2018, 64, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Medzhitov, R. Innate Immune Recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Kajla, M.; Kakani, P.; Choudhury, T.P.; Gupta, K.; Gupta, L.; Kumar, S. Characterization and Expression Analysis of Gene Encoding Heme Peroxidase HPX15 in Major Indian Malaria Vector Anopheles stephensi (Diptera: Culicidae). Acta Trop. 2016, 158, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Kajla, M.; Kakani, P.; Choudhury, T.P.; Kumar, V.; Gupta, K.; Dhawan, R.; Gupta, L.; Kumar, S. Anopheles Stephensi Heme Peroxidase HPX15 Suppresses Midgut Immunity to Support Plasmodium Development. Front. Immunol. 2017, 14, 249. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.; Choi, J.; Jang, A.; Choi, S.; Yeon, J.; Jang, M.; Lee, Y.; Son, K.; Shin, S.Y.; Jeong, M.S.; et al. Molecular Mechanism Underlying the TLR4 Antagonistic and Antiseptic Activities of Papiliocin, an Insect Innate Immune Response Molecule. Proc. Natl. Acad. Sci. USA 2022, 119, e2115669119. [Google Scholar] [CrossRef]
- Li, E.; Qin, J.; Feng, H.; Li, J.; Li, X.; Nyamwasa, I.; Cao, Y.; Ruan, W.; Li, K.; Yin, J. Immune-Related Genes of the Larval Holotrichia Parallela in Response to Entomopathogenic Nematodes Heterorhabditis Beicherriana LF. BMC Genom. 2021, 22, 192. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.; Arnot, C.J.; Beeston, H.; McCoy, A.; Ashcroft, A.E.; Gay, N.J.; Gangloff, M. Cytokine Spätzle Binds to the Drosophila Immunoreceptor Toll with a Neurotrophin-like Specificity and Couples Receptor Activation. Proc. Natl. Acad. Sci. USA 2013, 110, 20461–20466. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Heryanto, C. Transcriptomic Insights into the Insect Immune Response to Nematode Infection. Genes 2021, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, R.; Gupta, K.; Kajla, M.; Kumar, S.; Gakhar, S.K.; Kakani, P.; Choudhury, T.P.; Gupta, L. Molecular Characterization of SOCS Gene and Its Expression Analysis on Plasmodium Berghei Infection in Anopheles Culicifacies. Acta Trop. 2015, 152, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune Pathways and Defence Mechanisms in Honey Bees Apis Mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Qin, S.; Song, X.; Li, Y.; Jin, P.; Chen, L.; Ma, F. Evolutionary Rate Patterns of Genes Involved in the Drosophila Toll and Imd Signaling Pathway. BMC Evol. Biol. 2013, 13, 245. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; St. Leger, R.J.; Wu, L.P. Fungal Peptide Destruxin a Plays a Specific Role in Suppressing the Innate Immune Response in Drosophila Melanogaster. J. Biol. Chem. 2007, 282, 8969–8977. [Google Scholar] [CrossRef] [PubMed]
- Huot, L.; George, S.; Girard, P.A.; Severac, D.; Nègre, N.; Duvic, B. Spodoptera Frugiperda Transcriptional Response to Infestation by Steinernema Carpocapsae. Sci. Rep. 2019, 9, 12879. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Q.; Kanost, M.R. Manduca Sexta Lipopolysaccharide-Specific Immulectin-2 Protects Larvae from Bacterial Infection. Dev. Comp. Immunol. 2003, 27, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Agaisse, H.; Burrack, L.S.; Philips, J.A.; Rubin, E.J.; Perrimon, N.; Higgins, D.E. Genome-Wide RNAi Screen for Host Factors Required for Intracellular Bacterial Infection. Science 2005, 309, 248–1251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Wang, Y.; Wang, X.W.; Wang, L.; Mu, Y.; Wang, J.X. A C-Type Lectin with an Immunoglobulin-like Domain Promotes Phagocytosis of Hemocytes in Crayfish Procambarus Clarkii. Sci. Rep. 2016, 6, 29924. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Hussain, M.; Asgari, S. Suppression of Scavenger Receptors Transcription by Parasitoid Factors. Dev. Comp. Immunol. 2012, 38, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Gökçen, F.; Felföldi, G.; Millichap, P.J.; Trenczek, T.E.; Ffrench-constant, R.H.; Reynolds, S.E. The Immunoglobulin Family Protein Hemolin Mediates Cellular Immune Responses to Bacteria in the Insect Manduca Sexta. Cell. Microbiol. 2007, 9, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-Type Lectins in Innate Immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Arefin, B.; Kucerova, L.; Dobes, P.; Markus, R.; Strnad, H.; Wang, Z.; Hyrsl, P.; Zurovec, M.; Theopold, U. Genome-Wide Transcriptional Analysis of Drosophila Larvae Infected by Entomopathogenic Nematodes Shows Involvement of Complement, Recognition and Extracellular Matrix Proteins. J. Innate Immun. 2014, 6, 92–204. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.C.; Creasy, T.; Kumari, P.; Shetty, A.; Shokal, U.; Tallon, L.J.; Eleftherianos, I. Drosophila Anti-Nematode and Antibacterial Immune Regulators Revealed by RNA-Seq. BMC Genom. 2015, 16, 519. [Google Scholar] [CrossRef] [PubMed]
- Sackton, T.B.; Lazzaro, B.P.; Schlenke, T.A.; Evans, J.D.; Hultmark, D.; Clark, A.G. Dynamic Evolution of the Innate Immune System in Drosophila. Nat. Genet. 2007, 164, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Nei, M. Pattern of Nucleotide Substitution at Major Histocompatibility Complex Class I Loci Reveals Overdominant Selection. Nature 1988, 335, 167–170. [Google Scholar] [CrossRef]
- Wang, C.; St. Leger, R.J. A Collagenous Protective Coat Enables Metarhizium Anisopliae to Evade Insect Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 6647–6652. [Google Scholar] [CrossRef] [PubMed]
- Oduol, F.; Xu, J.; Niaré, O.; Natarajan, R.; Vernick, K.D. Genes Identified by an Expression Screen of the Vector Mosquito Anopheles Gambiae Display Differential Molecular Immune Response to Malaria Parasites and Bacteria. Proc. Natl. Acad. Sci. USA 2000, 97, 11397–11402. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Raw_Bases | Valid Reads | Valid_Bases | Valid% | Q20% | Q30% | GC% |
|---|---|---|---|---|---|---|---|---|
| Control slug1 | 41,649,952 | 6.29 G | 38,996,996 | 5.39G | 93.63 | 97.81 | 93.85 | 37.50 |
| Control slug2 | 45,331,020 | 6.84 G | 42,417,080 | 5.87G | 93.57 | 98.04 | 94.38 | 40.81 |
| Control slug3 | 52,415,534 | 7.91 G | 49,437,474 | 6.86G | 94.32 | 98.05 | 94.33 | 40.31 |
| Nematode-infected slug1 | 47,876,956 | 7.23 G | 45,098,082 | 6.27G | 94.20 | 97.86 | 94.17 | 39.40 |
| Nematode-infected slug2 | 47,830,174 | 7.22 G | 45,316,882 | 6.33G | 94.75 | 98.05 | 94.15 | 37.55 |
| Nematode-infected slug3 | 44,114,910 | 6.66 G | 41,826,018 | 5.82G | 94.81 | 98.08 | 94.28 | 37.89 |
| Databases (DB) | Number of Unigenes | Ratio (%) |
|---|---|---|
| All | 64,679 | 100.00 |
| GO | 7786 | 12.04 |
| KEGG | 7767 | 12.01 |
| Pfam | 7787 | 12.04 |
| swissprot | 7049 | 10.90 |
| eggNOG | 9618 | 14.87 |
| NR | 13,285 | 20.54 |
| All | 64,679 | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafeez, M.; Mc Donnell, R.; Colton, A.; Howe, D.; Denver, D.; Martin, R.C.; Choi, M.-Y. Immune-Related Gene Profiles and Differential Expression in the Grey Garden Slug Deroceras reticulatum Infected with the Parasitic Nematode Phasmarhabditis hermaphrodita. Insects 2024, 15, 311. https://0-doi-org.brum.beds.ac.uk/10.3390/insects15050311
Hafeez M, Mc Donnell R, Colton A, Howe D, Denver D, Martin RC, Choi M-Y. Immune-Related Gene Profiles and Differential Expression in the Grey Garden Slug Deroceras reticulatum Infected with the Parasitic Nematode Phasmarhabditis hermaphrodita. Insects. 2024; 15(5):311. https://0-doi-org.brum.beds.ac.uk/10.3390/insects15050311
Chicago/Turabian StyleHafeez, Muhammad, Rory Mc Donnell, Andrew Colton, Dana Howe, Dee Denver, Ruth C. Martin, and Man-Yeon Choi. 2024. "Immune-Related Gene Profiles and Differential Expression in the Grey Garden Slug Deroceras reticulatum Infected with the Parasitic Nematode Phasmarhabditis hermaphrodita" Insects 15, no. 5: 311. https://0-doi-org.brum.beds.ac.uk/10.3390/insects15050311