
Broad-Spectrum Resistance and Monogenic Inheritance of Bacterial Blight Resistance in an Indigenous Upland Rice Germplasm ULR207


Abstract
:1. Introduction
2. Materials and Methods
2.1. Confirmation of Broad-Spectrum Resistance for Bacterial Blight Disease
2.1.1. Plant Materials
2.1.2. Bacterial Isolates and Inoculum Preparation
2.1.3. The Inoculation of Bacterial Blight
2.1.4. Data Analysis
2.1.5. The Identification of BB Resistance Gene in Indigenous Upland Rice ULR207
2.2. Genetic Analysis of Bacterial Blight Resistance in ULR207
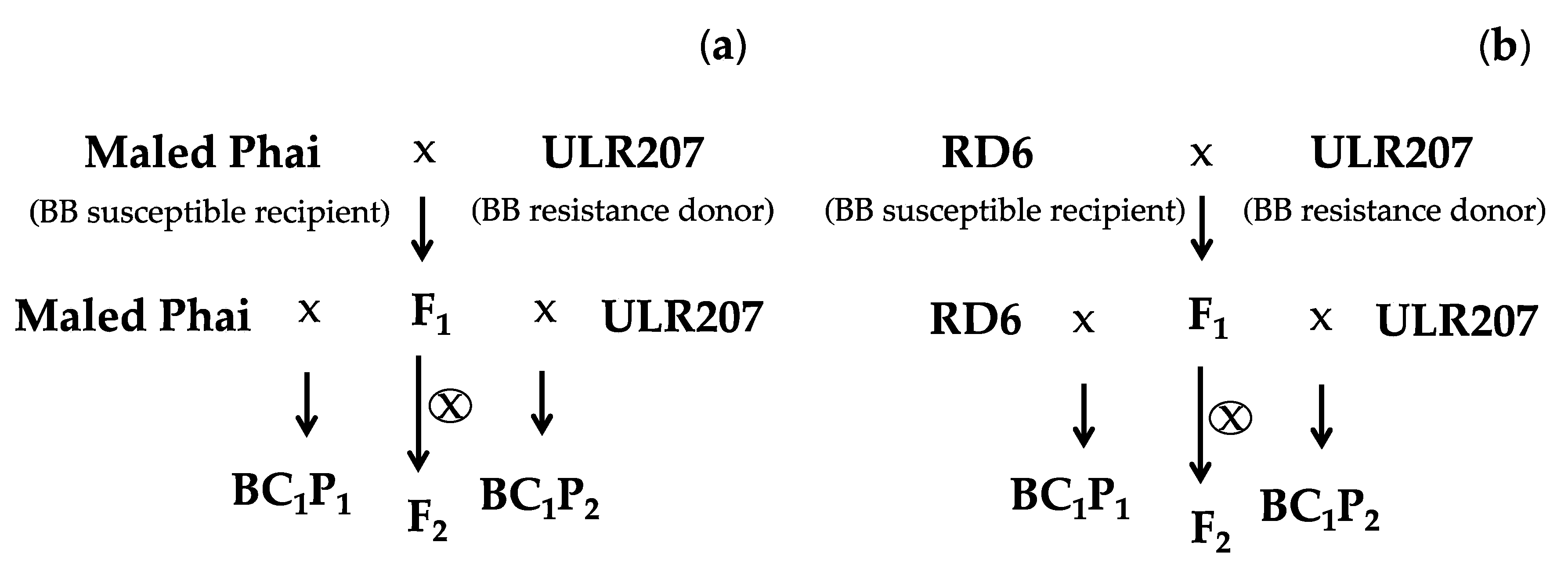
2.2.1. Population Construction
2.2.2. Pathogenic Assay of BB Resistance
2.2.3. Statistical Analysis
3. Results
3.1. Confirmation of Broad-Spectrum Resistance for Bacterial Blight Disease
Survey of R Gene
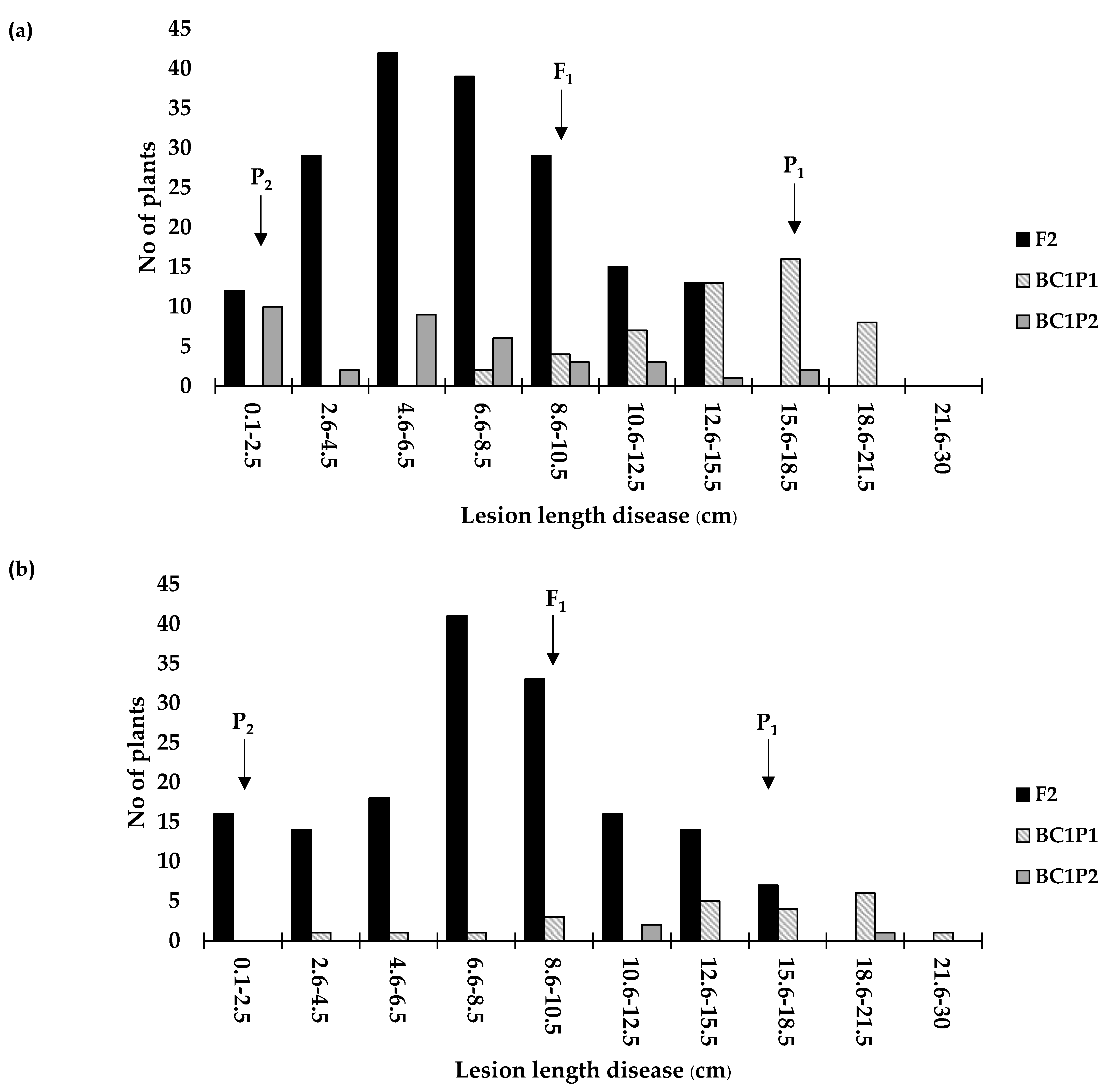
3.2. Genetic Analysis
4. Discussion
4.1. Local Rice Germplasm and Broad-Spectrum Resistance in Bacterial Blight Disease
4.2. Genetic Resistance of Bacterial Blight in ULR207
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.H.; Francis, J.A.; Miller, J.R. Surface temperature of the arctic: Comparison of TOVS satellite retrievals with surface observations. J. Clim. 2002, 15, 3698–3708. [Google Scholar] [CrossRef]
- Kosawang, C.; Smitamana, P.; Toojinda, T.; Nilpanit, N.; Sirithunya, P. Amplified Fragment Length Polymorphism Fingerprinting Differentiates Genetic Diversity of Xanthomonas oryzae pv. oryzae from Northern Thailand. J. Phytopathol. 2006, 154, 550–555. [Google Scholar] [CrossRef]
- Chen, L.N.; Yang, Y.; Yan, C.; Wang, X.M. Identification of Quantitative Trait Loci for Bacterial Blight Resistance Derived from Oryza meyeriana and Agronomic Traits in Recombinant Inbred Lines of Oryza sativa. Phytopathology 2012, 160, 19. [Google Scholar] [CrossRef]
- Korinsak, S.; Darwell, C.T.; Wanchana, S.; Praphaisal, L.; Korinsak, S.; Thunnom, B.; Patarapuwadol, S.; Toojinda, T. Identification of bacterial blight resistance loci in rice (Oryza sativa L.) against diverse Xoo Thai strains by genome-wide association study. Plants 2021, 10, 518. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T. Report of committee on gene symbolization nomenclature and linkage group. Rice Genet. Newsl. 1995, 12, 9–153. Available online: https://shigen.nig.ac.jp/rice/oryzabase/asset/rgn/vol3/v3C.html (accessed on 1 August 2023).
- Lin, X.H.; Zhang, D.P.; Xie, Y.F.; Gao, H.P.; Zhang, Q.F. Identifying and mapping a new gene forbacterial blight resistance in rice based on RFLP marker. Phytopathology 1996, 86, 1156–1159. [Google Scholar] [CrossRef]
- Zhang, H.; Jia, J.; Gale, M.D.; Devos, K.M. Relationship between the chromosomes of Aegilops umbellulata and wheat. Theor. Appl. Genet. 1998, 96, 69–75. [Google Scholar] [CrossRef]
- Furuya, N.; Taura, S.; Goto, T.; Thuy, B.T.; Ton, P.H.; Tsuchiya, K.; Yoshimura, A. Diversity in virulence of Xanthomonasoryzae pv. oryzae from Northern Vietnam. Jpn. Agric. Res. Q. 2012, 46, 329–338. [Google Scholar] [CrossRef]
- Mishra, D.; Vishnupriya, M.R.; Anil, M.G.; Konda, K.; Raj, Y.; Sonti, R.V. Pathotype and Genetic Diversity amongst Indian Isolates of Xanthomonas oryzae pv. oryzae. PLoS ONE 2013, 8, e81996. [Google Scholar] [CrossRef]
- Periyannam, S.; Milne, R.J.; Figueroa, M.; Lagudah, E.S.; Dodds, P.N. An overview of genetic rust resistance: From broad to specific mechanisms. PLOS Pathog. 2017, 13, e1006380. [Google Scholar] [CrossRef]
- Wongkhamchan, A.; Chankaew, S.; Monkham, T.; Saksirirat, W.; Sanitchon, J. Broad resistance of RD6 introgression lines with xa5 gene from IR62266 rice variety to bacterial leaf blight disease for rice production in Northeastern Thailand. Agric. Nat. Resour. 2018, 52, 21–245. [Google Scholar] [CrossRef]
- Korinsak, S.; Sirithunya, K.; Toojinda, T. Identifying a source of a bacterial blight resistance gene xa5 in rice variety ‘IR62266′ and development of a functional marker ‘PAxa5′, the easy agarose-based detection. Thai J. Genet. 2014, 7, 164–172. [Google Scholar] [CrossRef]
- Sombunjitt, S.; Tanee, S.; Chatuporn, K.; Vipa, H. Searching for and analysis of bacterial blight resistance genes from Thailand rice germplasm. Agric. Nat. Resour. 2017, 65, 365–375. [Google Scholar] [CrossRef]
- Sontornkarun, T.; Chankaew, S.; Sanitchon, J. Donor parental determination for breeding the RD41 rice cultivar to improve bacterial blight resistance. Khon Kaen Agric. J. 2020, 48, 1162–1171. [Google Scholar] [CrossRef]
- Kwanwah, M.R.; Wongsa, T.; Monkham, T.; Chankaew, S.; Falab, S.; Sanitchon, J. Thai Indigenous Lowland Rice Germplasms: Sources of Bacterial Blight Disease Resistance and Agronomic Attributes. AGRIVITA J. Agric. Sci. 2020, 42, 367–380. [Google Scholar] [CrossRef]
- Chumpol, A.; Monkham, T.; Saepaisan, S.; Sanitchon, J.; Falab, S.; Chankaew, S. Phenotypic broad spectrum of bacterial blight disease resistance from Thai indigenous upland rice germplasm implies novel genetic resource for breeding program. Agron. J. 2022, 12, 1930. [Google Scholar] [CrossRef]
- Fouad, H.M. Six Generations Mean Analysis Using Scaling and Joint Scaling Tests in Faba Bean (Vicia faba L.). J. Sustain. Agric. 2020, 46, 1–11. [Google Scholar] [CrossRef]
- Perumalsamy, S.; Bharani, M.; Sudah, M.; Nagarajan, P.; Arul, L.; Sarawathi, R.; Balasubramaninan, P.; Ramalingam, J. Functional marker-assisted selection for bacterial leaf blight resistance genes in rice (Oryza sativa L.). Int. J. Plant Breed. 2010, 129, 400–406. [Google Scholar] [CrossRef]
- Rajpurohit, D.; Kumar, R.; Kumar, M.; Paul, P.; Awasthi, A.; Basha, P.O.; Puri, A.; Jhang, T.; Singh, K.; Dhaliwal, H.S. Pyramiding of two bacterial blight resistance and a semi dwarfing gene in Type 3 Basmati using marker-assisted selection. Euphytica 2010, 178, 111–126. [Google Scholar] [CrossRef]
- Arif, M.; Jaffar, M.; Babar, M.; Sheikh, M.A.; Kousar, S.; Arif, A.; Zafar, Y. Identification of bacterial blight resistance genes Xa4 in Pakistani rice germplasm using PCR. Afr. J. Biotechnol. 2008, 7, 541–545. Available online: https://www.researchgate.net/publication/27798155_Identification_of_bacterial_blight_resistance_genes_Xa4_in_Pakistani_rice_germplasm_using_PCR (accessed on 1 August 2023).
- Muhammad, W.K.; Fida, M.A.; Mohammed, S.M.; Ashiq, R.; Muniba, F.A.; Muhammad, S.; Uzma, K.; Habib, A. Identification of bacterial blight resistance gene Xa7 in rice (Oryzae sativa L.) through STS marker. Int. J. Biosci. 2015, 6, 318–324. [Google Scholar]
- Ullah, I.; Jamil, S.; Iqbal, M.Z.; Shaheen, H.L.; Hasni, S.M.; Jabeen, S. Detection of bacterial blight resistance genes in basmati rice landraces. Genet. Mol. Res. 2012, 11, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Vikal, Y.; Bhatia, D. Genetics and genomics of bacterial blight resistance in rice. Adv. Int. Rice Res. 2017, 10, 175–213. [Google Scholar] [CrossRef]
- Viana, J.M.S. Generation mean analysis to polygenic systems with epistasis and fixed genes. Pesqui. Agropecu. Bras. 2000, 35, 1159–1167. [Google Scholar] [CrossRef]
- Kearsey, M.J.; Pooni, H.S. The Genetical Analysis of Quantitative Traits; Chapman and Hall: London, UK, 2004; pp. 18–52. [Google Scholar]
- Deb, A.C.; Khaleque, M.A. Nature of gene action of some quantitative traits in chickpea (Cicer arietinum L.). World J. Agric. Sci. 2009, 5, 361–368. Available online: https://www.researchgate.net/publication/308026816_Nature_of_gene_action_of_some_quantitative_traits_in_chickpea (accessed on 1 August 2023).
- Abd-El-Haleem, S.H.; Metwali, E.M.; Al-Felaly, A.M. Genetic analysis of yield and its components of some Egyptian cotton (Gossypium barbadense L.) varieties. World J. Agric. Sci. 2010, 6, 615–621. [Google Scholar] [CrossRef]
- Eshghi, R.; Akhundova, E. Genetic Diversity of the Monomeric Prolamins and Hordein in Hulless Barley Genotypes and Their Relation with Agronomical Traits. Afr. J. Biotechnol. 2009, 8, 1819–1826. [Google Scholar]
- Azizi, F.; Rezai, A.M.; Saeidi, G. Generation mean analysis to estimate genetic parameters for different traits in two crosses of corn inbred lines at three planting densities. J. Agric. Sci. Technol. 2006, 8, 153–169. [Google Scholar] [CrossRef]
- Nikita, K.; Rajesh, K.; Avinash, K. Genetic variability, and association of traits in mutant lines of rice (Oryza sativa L.) for submergence tolerance. Curr. Appl. Sci. Technol. 2019, 33, 1–7. [Google Scholar] [CrossRef]
- Thandar, K.; Laosatit, K.; Yamram, T.; Somta, P. Genetic analysis of seed resistance to Callosobruchus chinensis and Callosobruchus maculatus in cowpea. J. Stored Prod. Res. 2021, 92, 101783. [Google Scholar] [CrossRef]
- Ramli, A.B.; Rafii, M.Y.; Latif, M.A.; Saleh, G.B.; Omar, O.B.; Puteh, A.B. Generation mean analysis of grain quality traits in selected rice populations derived from different amylose characteristics. J. Sci. Food Agric. 2016, 96, 1593–1600. [Google Scholar] [CrossRef]
- Sribunrueang, A.; Chankaew, S.; Thummabenjapone, P.; Sanitchon, J. Stability of four new sources of bacterial leaf blight resistance in Thailand obtained from indigenous rice varieties. Agrivita 2017, 39, 128–136. [Google Scholar] [CrossRef]
- Kauffman, H.E.; Reddy, A.P.K.; Hsieh, S.P.Y.; Merca, S.D. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19731306925 (accessed on 10 August 2023).
- IRRI. Standard Evaluation System for Rice; International Rice Research Institute: Los Banos, Philippines, 2002; pp. 1–45. Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf (accessed on 10 August 2023).
- Ahn, S.W. International collaboration on breeding for resistance to rice blast. In Rice Blast Disease; Zeigler, R.S., Leong, S.A., Teng, P.S., Eds.; CABI: Wallingford, UK, 1994; pp. 137–153. Available online: https://agris.fao.org/search/en/providers/122430/records/6471f6302a40512c710f03de (accessed on 25 March 2024).
- Sirithunya, P.; Tragoonrung, S.; Vanavichit, A.; Pa-In, M.; Vongsaprom, C.; Toojinda, T. Quantitative trait loci associated with leaf and neck blast resistance in recombinant inbred line population of rice (Oryza sativa). DNA Res. 2002, 9, 79–88. [Google Scholar] [CrossRef]
- Madden, L.V.; Hughes, G.; Bosch, V.D. The Study of Plant Disease Epidemics; American Phytopathology Society: St. Paul, MN, USA, 2007; pp. 63–116. [Google Scholar] [CrossRef]
- International Rice Research Institute (IRRI). Standard Evaluation System for Rice (SES), 3rd ed.; International Rice Research Institute: Manila, Philippines, 1996. [Google Scholar]
- Mather, K. The Measurement of Linkage in Heredity; Methuen: London, UK, 1951. [Google Scholar]
- Pearson, K. On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Lond. Edinb. Dublin Philos. Mag. J. Sci. 1900, 50, 157–175. [Google Scholar] [CrossRef]
- Cavalli, L.L. An analysis of linkage in quantitative inheritance. In Quantitative Inheritance; Reeve, E.C.R., Waddington, C.H., Eds.; HMSO: London, UK, 1952; pp. 135–141. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19541603514 (accessed on 10 August 2023).
- Mather, K.; Jinks, J.L. Biometrical Genetics, 3rd ed.; Chapman and Hall Ltd.: London, UK, 1982. [Google Scholar]
- Warner, J.N. A method for estimating heritability. Agron. J. 1952, 44, 427–430. [Google Scholar] [CrossRef]
- Poehlman, J.M. Breeding Field Crops, 3rd ed.; Van Nostrand Reinhold Catalysis Series: New York, NY, USA, 1987; pp. 38–86. Available online: https://0-link-springer-com.brum.beds.ac.uk/book/10.1007/978-94-015-7271-2?page=1#toc (accessed on 10 August 2023).
- Prakash, S.; Singh, H.B.; Singh, O.N. Inheritance of bacterial leaf blight (Xanthomanas oryzae pv. Oryzae) resistance in indica rice cultivar HUR4-3. Int. J. Agric. Environ. Biotechnol. 2014, 7, 777–785. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Keller, B. Resistance: Double gain with one gene. Nat. Plants 2017, 3, 17019. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yin, F.; Chen, L.; Li, D.; Xiao, S.; Zhong, Q.; Wang, B.; Ke, X.; Fu, J.; Li, X. Identification of bacterial blight resistance genes in rice landraces from Yunnan Province, China. Australas. Plant Pathol. 2022, 51, 59–69. [Google Scholar] [CrossRef]
- Pinta, W.; Toojinda, T.; Thummabenjapone, P.; Sanitchon, J. Pyramiding of blast and bacterial leaf blight resistance genes into rice cultivar RD6 using marker assisted selection. Afr. J. Biotechnol. 2013, 12, 4432–4438. [Google Scholar] [CrossRef]
- Aung Nan, M.S.; Janto, J.; Sribunrueang, A.; Chankaew, S.; Monkham, T.; Sanitchon, J. Field Evaluation of RD6 Introgression Lines for Yield Performance, Blast, and Bacterial Blight Resistance; and their Cooking and Eating Qualities. Agronomy 2019, 9, 825. [Google Scholar] [CrossRef]
- Leonard, K.J. Selection pressure and plant pathogens. Ann. N. Y. Acad. Sci. 1977, 287, 207–222. [Google Scholar] [CrossRef]
- Sakaguchi, S. Linkage studies on the resistance to bacterial leaf blight, Xanthomonas oryzae (Uyeda et Ishiyama) Dowson, in rice. Bull. Natl. Inst. Agric. Sci. Ser. 1967, 16, 1–18. Available online: https://www.cabidigitallibrary.org/doi/full/0.5555/19671103426 (accessed on 10 August 2023).
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. USA 1998, 95, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Ezuka, A.; Horino, O.; Toriyama, K.; Shinoda, H.; Morinaka, T. Inheritance of resistance of rice variety Wase Aikoku 3 to Xanthomonas oryzae. Bull. Tokai-Kinki Natl. Agric. Exp. Stn. 1975, 28, 124–130. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/19751320328 (accessed on 14 August 2023).
- Yoshimura, S.; Yoshimura, A.; Saito, A.; Kishimoto, N.; Kawase, M.; Yano, M.; Nakagahra, M.; Ogawa, T.; Iwata, N. RFLP analysis of introgressed chromosomal segments in three near-isogenic lines of rice for bacterial blight resistance genes, Xa-1, Xa-3, and Xa-4. Jpn. J. Genet. 1992, 67, 29–37. [Google Scholar] [CrossRef]
- Gao, L.F.; Cao, Y.H.; Xia, Z.H.; Jiang, G.H.; Liu, G.Z.; Zhang, W.X.; Zhai, W.X. Do transgenesis and marker-assisted backcross breeding produce substantially equivalent plants. A comparative study of transgenic and backcross rice carrying bacterial blight resistant gene Xa21. BMC Genom. 2013, 14, 738. [Google Scholar] [CrossRef] [PubMed]
- Leach, J.E.; Leung, H.; Tisserat, N.A. Plant Disease and Resistance; Encyclopedia of Agriculture and Food Systems: Davis, CA, USA, 2014; pp. 360–371. [Google Scholar] [CrossRef]
- Banerjee, A.; Somnath, R.; Manas, K.B.; Someswar, B.; Meera, K.K.; Mandal, M.P.; Arup, K.M.; Dipankar, M. A survey of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice germplasm from eastern and northeastern India using molecular markers. Crop. Prod. 2018, 112, 168–176. [Google Scholar] [CrossRef]
- Hasan, N.A.; Mohd, Y.R.; Harun, A.R.; Ahmad, F.; Ismail, N. Identification of bacterial leaf blight resistance genes in Malaysian local rice varieties. Genet. Mol. Res. 2020, 19, 18545. [Google Scholar] [CrossRef]
- Said, A.A. Generation mean analysis in wheat (Triticum aestivum L.) under drought stress conditions. Ann. Agric. Sci. 2014, 59, 177–184. [Google Scholar] [CrossRef]
- Mir, G.N.; Khush, G.S. Genetics of resistance to bacterial blight in rice (Oryza sativa L.). Indian J. Genet. 1991, 51, 72–78. Available online: https://www.isgpb.org/journal/index.php/IJGPB/article/view/3076 (accessed on 14 August 2023).
- Lee, K.S.; Rasabandith, S.; Angeles, E.R.; Khush, G.S. Inheritance of resistance to bacterial blight in 21 cultivars of rice. Phytopathology 2003, 93, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Kameswara, K.R.; Lakshminarasub, M.; Jena, K.K. DNA markers and marker-assisted breeding for durable resistance to bacterial blight disease in rice. Biotechnol. Adv. 2002, 20, 33–47. [Google Scholar] [CrossRef]
- Iyer-Pascuzzi, A.S.; McCouch, S.R. Recessive resistance genes and the Oryza sativa-Xanthomonas oryzae pv. oryzae pathosystem. Mol. Plant Microbe Interact. 2007, 20, 731–739. [Google Scholar] [CrossRef]
- Shanti, M.L.; Shenoy, V.V.; Devi, G.L.; Kumar, V.M.; Premalatha, P.; Kumar, G.N.; Shashidhar, H.E.; Zehr, U.B.; Freeman, W.H. Marker-assisted breeding for resistance to bacterial leaf blight in popular cultivar and parental lines of hybrid rice. Plant Pathol. J. 2010, 1, 495–501. [Google Scholar] [CrossRef]
- Acharya, A.; Adhikari, N.R.; Amgain, R.B.; Poudel, A.; Yadav, R.; Poudyal, K. Identification of rice genotypes resistance to bacterial leaf blight disease using SSR markers. J. Inst. Agric. Anim. Sci. 2018, 35, 113–120. [Google Scholar] [CrossRef]
- Kumar, M.; Singh, R.P.; Singh, O.N.; Singh, P. Genetic analysis for bacterial blight resistance in indica rice (Oryza sativa L.) cultivars. Oryza Int. J. Rice 2019, 56, 247–255. [Google Scholar] [CrossRef]
- Divvy, B.; Biswas, A.; Robin, S.; Rabindran, R.; Joel John, A. Gene interactions and genetics of blast resistance and yield attributes in rice (Oryza sativa L.). J. Genet. 2014, 93, 415–424. [Google Scholar] [CrossRef]
- Khan, I.A.; Hussain, M.; Rauf, S.; Khan, T.M. Inheritance of resistance to cotton leaf curl virus in cotton (Gossypium hirsutum L.). Plant Prot. Sci. 2007, 43, 5–9. [Google Scholar] [CrossRef]
- Karami, E.; Talebi, R. Nature of gene action and genetic parameters for yield and its components in chickpea. Afr. J. Biotechnol. 2013, 12, 7038–7042. [Google Scholar] [CrossRef]
- Usman, I.; Smiullah, S.; Muhammad, K. Genetic study of quantitative traits in spring wheat through generation means analysis. Am.-Eurasian J. Agric. Environ. Sci. 2013, 13, 191–197. [Google Scholar] [CrossRef]
- Sari, W.K.; Nualsri, C.; Junsawang, N.; Soonsuwon, W. Combining ability and heritability for yield and its related traits in Thai upland rice (Oryza sativa L.). Agric. Nat. Resour. 2019, 54, 229–236. [Google Scholar] [CrossRef]
- Sleper, D.A.; Poehlman, J.M. Breeding Field Crops, 4th ed.; Blackwell Publishing: Columbia, SC, USA, 2006; pp. 345–366. [Google Scholar]
- Govintharaj, P.; Manonmani, S.; Robin, S. Variability and genetic diversity study in an advanced segregating population of rice with bacterial blight resistance genes introgressed. Agric. Sci. 2018, 42, 291–296. [Google Scholar] [CrossRef]
- Govintharaj, P.; Tannidi, S.; Manonmani, S.; Robin, S. Genetic parameters studies on bacterial blight resistance genes introgressed segregating population in Rice. World Sci. News. 2016, 59, 85–96. Available online: https://www.researchgate.net/publication/310043914_Genetic_parameters_studies_on_bacterial_blight_resistance_genes_introgressed_segregating_population_in_Rice (accessed on 14 August 2023).
- Fiyaz, R.A.; Ramya, K.T.; Chikkalingaiah, C.; Ajay, B.C.; Gireesh, C.; Kulkarni, R.S. Genetic variability, correlation, and path coefficient analysis studies in rice (Oryza sativa L.) under alkaline soil condition. Electron. J. Plant Breed. 2011, 2, 531–537. Available online: https://www.indianjournals.com/ijor.aspx?target=ijor:ejpb&volume=2&issue=4&article=011 (accessed on 14 August 2023).
- Keller, E.F. The Century of the Gene; Harvard University Press: Cambridge, MA, USA, 2002; p. 186. [Google Scholar]
- Subhas, C.R.; Shil, P. Assessment of genetic heritability in rice breeding lines based on morphological traits and caryopsis ultrastructure. Sci. Rep. 2020, 10, 7830. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Sources | BB Resistance Genes |
|---|---|---|
| ULR207 | Indigenous, Thailand | Unknown |
| IRBB1 | IRRI | Xa1 |
| IRBB3 | IRRI | Xa3 |
| IRBB4 | IRRI | Xa4 |
| IRBB5 | IRRI | xa5 |
| IRBB7 | IRRI | Xa7 |
| IRBB8 | IRRI | xa8 |
| IRBB10 | IRRI | Xa10 |
| IRBB11 | IRRI | Xa11 |
| IRBB13 | IRRI | xa13 |
| IRBB14 | IRRI | Xa14 |
| IRBB21 | IRRI | Xa21 |
| IR62266 | IRRI | Resistance check |
| IR21 | IRRI | Resistance check |
| RD6 | Department of rice, Thailand | Susceptible check |
| KDML105 | Department of rice, Thailand | Susceptible check |
| Maled Phai | Indigenous, Thailand | Susceptible check |
| Isolates | Sources | |
|---|---|---|
| Provinces | Part of Thailand | |
| UT2-1 | Uthai Thani | Central |
| CM4-1 | Chiang Mai | Northern |
| CM3-1 | Chiang Mai | Northern |
| NB7-7 | Nonthaburi | Central |
| PR5-1 | Prachinburi | Eastern |
| NB7-8 | Nonthaburi | Central |
| CN2-1 | Chainat | Central |
| NY1-1 | Nakhon Nayok | Central |
| SP1-1 | Suphan Buri | Central |
| MS1-2 | Maha Sarakham | Northeastern |
| Gene | Marker Name | Type of Marker | Forward (5′-3′) | Reverse (3′-5′) |
|---|---|---|---|---|
| Xa1 | Xa1 | Gene specific | ACTGCCCTCTTGCACACGCCTTTGG | CCGGTACATCAGTATTGTCCATCGG |
| Xa3 | RM113 | SSR | CACCATTGCCCATCAGCACAAC | TCGCCCTCTGCTGCTTGATGGC |
| Xa4 | RM224 | SSR | ATCGATCGATCTTCACGAGG | TGCTATAAAAGGCATTCGGG |
| xa5 | PAxa5 | Gene specific | CTGGAAGAAGCTCTTAATTT | GATTCCTTTAGCAAGGTGTG |
| Xa7 | Xa7 | Gene linked | CGATCTTACTGGCTCTGCAACTCTGT | GCATGTCTGTGTCGATTCGTCCGTACGA |
| xa13 | xa13 | Gene specific | AGCTCCAGCTCTCCAAATG | CATTGCTACTGGTGATGAAGG |
| xa8 | RM214 | SSR | CTGATGATAGAAACCTCTTCTC | AAGAACAGCTGACTTCACAA |
| Xa10 | RM206 | SSR | CCCATGCGTTTAACTATTCT | CGTTCCATCGATCCGTATGG |
| Xa11 | RM1350 | SSR | CGCCCTAGTAGATAGGTAATTG | AAATCAGCAAGAAAGCTCTG |
| Xa14 | RM303 | SSR | GCATGGCCAAATATTAAAGG | GGTTGGAAATAGAAGTTCGGT |
| Xa21 | RM21 | SSR | ACAGTATTCCGTAGGCACGG | GCTCCATGAGGGTGGTAGAG |
| Varieties | Isolates Code/Disease Lesion Length (cm) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes | UT2-1 | CM4-1 | CM3-1 | NB7-7 | PR5-1 | NB7-8 | CN2-1 | NY1-1 | SP1-1 | MS1-2 | BSR | |
| ULR207 | - | 6.58 | 11 | 3.96 | 8.66 | 5.53 | 4.81 | 5.85 | 12.73 | 6.09 | 8.18 | 0.8 |
| Maled Phai | - | 13.31 | 13.79 | 11.02 | 15.18 | 12.72 | 11.87 | 11.18 | 12.56 | 17.51 | 17.71 | 0 |
| RD6 | - | 17.28 | 19.92 | 16.88 | 21.34 | 16.27 | 19.64 | 12.01 | 20.69 | 21.34 | 22.33 | 0 |
| KDML105 | - | 21.84 | 21.8 | 18.55 | 26.81 | 19.14 | 24.49 | 16.54 | 17.7 | 25.96 | 22.32 | 0 |
| IRBB21 | Xa21 | 16.19 | 18.53 | 13.89 | 16.31 | 13.96 | 15.96 | 10.21 | 13.13 | 17.05 | 17.96 | 0 |
| IRBB1 | Xa1 | 3.03 | 8.94 | 0.76 | 7.18 | 8.64 | 4.89 | 5.38 | 7.4 | 7.5 | 7.87 | 1.0 |
| IRBB3 | Xa3 | 6.28 | 8.77 | 6.94 | 6.23 | 3.59 | 5.27 | 3.04 | 4.02 | 5.58 | 6.17 | 1.0 |
| IRBB14 | Xa14 | 15.25 | 16.26 | 12.65 | 15.22 | 13.98 | 19.14 | 10.88 | 15.97 | 14.92 | 16.99 | 0 |
| IRRBB13 | Xa13 | 18.36 | 15.78 | 12.74 | 16.64 | 16.01 | 12.98 | 10.82 | 15.13 | 15.98 | 16.37 | 0 |
| IR21 | - | 18.35 | 16.74 | 13.46 | 19.07 | 14.48 | 17.63 | 9.77 | 15.99 | 12.72 | 20.87 | 0.1 |
| IRBB11 | Xa11 | 13.41 | 9.88 | 11.09 | 15.73 | 12.94 | 16.03 | 9.72 | 11.11 | 12.32 | 13.43 | 0.2 |
| IRBB4 | Xa4 | 15.64 | 4.87 | 7.71 | 16.52 | 13.93 | 17.97 | 10.89 | 5.82 | 12.48 | 18.04 | 0.3 |
| IRBB5 | Xa5 | 4.08 | 1.61 | 1.79 | 2.19 | 1.89 | 2.46 | 2.14 | 3.31 | 1.11 | 1.63 | 1.0 |
| IR62266 | Xa21 | 13.86 | 2.38 | 2.03 | 9.32 | 12.88 | 10.18 | 8.32 | 1.77 | 11.64 | 12.02 | 0.7 |
| IRBB8 | Xa8 | 8.98 | 6.15 | 7.39 | 7.88 | 6.15 | 5.09 | 4.53 | 10.45 | 7.12 | 7.26 | 1.0 |
| IRBB10 | Xa10 | 16.17 | 14.07 | 13.19 | 12.98 | 13.3 | 18.75 | 9.67 | 15.21 | 16.84 | 18.75 | 0 |
| IRBB7 | Xa7 | 1.85 | 0.9 | 0.28 | 2.33 | 2.31 | 0.61 | 0.47 | 0.26 | 0.14 | 2.07 | 1.0 |
| F-test | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| C.V.% | 18.52 | 17.61 | 12.41 | 16.75 | 23.41 | 18.38 | 19.68 | 39.73 | 23.75 | 20.72 | ||
| Variety | Disease Reaction | Amplicon Length Polymorphism (bp) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Xa1 | Xa3 | Xa4 | xa5 | Xa7 | xa8 | Xa10 | Xa11 | xa13 | Xa14 | Xa21 | ||
| Gene Specific | RM114 | RM224 | PAxa5 | Gene Linked | RM214 | RM206 | RM1350 | RM224 | RM303 | RM21 | ||
| ULR207 | R | 600 [-] | 180 [-] | 150 [-] | 221 [-] | 1100 [-] | 180 [-] | 180 [-] | 220 [-] | 500 [-] | 210 [-] | 200 [-] |
| KDML105 | S | 600 [-] | 220 [-] | 250 [-] | 221 [-] | 1100 [-] | 120 [+] | 190 [-] | 220 [-] | 500 [-] | 240 [-] | 180 [+] |
| Positive control | R | IRBB1 (600) [-] | IRBB3 (190) [+] | IRBB4 (200) [+] | IRBB5 (134) [+] | IRBB7 (300) [+] | IRBB8 (120) [+] | IRBB10 (200) [+] | IRBB11 (210) [+] | IRBB13 (500) [-] | IRBB14 (220) [+] | IRBB21 (180) [+] |
| Negative control | S | IR24 (600) [-] | IR24 (200) [-] | IR24 (180) [-] | IR24 (221) [-] | IR24 (1000) [-] | IR24 (120) [+] | IR24 (180) [-] | IR24 (220) [-] | IR24 (500) [-] | IR24 (240) [-] | IR24 (200) [-] |
| Cross | Population | No. of Plants | No. of Plants * | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Resistant | Susceptible | Ratio | Chi-Square | p-Value | |||||
| O | E | O | E | ||||||
| Maled Fai x ULR207 | F2 | 179 | 47 | 44 | 132 | 135 | 1:3 | 0.24 | 0.96 |
| BC1P2 | 33 | 13 | 17 | 22 | 17 | 1:1 | 2.41 | 0.49 | |
| RD6 x ULR207 | F2 | 159 | 37 | 39 | 122 | 120 | 1:3 | 0.14 | 0.98 |
| BC1P2 | 3 | 0 | 1.5 | 3 | 1.5 | 1:1 | 3.00 | 0.39 | |
| Gene Action/Scaling Test | Crosses | |
|---|---|---|
| Maled Phai x ULR207 | RD6 x ULR207 | |
| A | 36.56 ns | 0.68 ns |
| B | 1.40 ns | 22.05 ns |
| C | 35.74 ns | 43.11 ns |
| m (mean) | 65.63 ** | 49.57 ** |
| d (additive) | 7.30 ** | 7.59 ** |
| h (dominant) | 17.24 ns | 46.18 ns |
| i (additive × additive) | 4.65 ns | 13.77 ns |
| l (dominant × dominant) | 3.51 ns | −17.58 ns |
| j (additive × dominant) | −10.97 ns | −32.72 ns |
| Crosses | Heritability | No. of Gene | |
|---|---|---|---|
| Broad Sense | Narrow Sense | ||
| Maled Fai x ULR207 | 0.817 | 0.709 | 1.4 |
| RD6 x ULR207 | 0.716 | 0.621 | 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wongsa, T.; Chankaew, S.; Monkham, T.; Sanitchon, J. Broad-Spectrum Resistance and Monogenic Inheritance of Bacterial Blight Resistance in an Indigenous Upland Rice Germplasm ULR207. Agronomy 2024, 14, 898. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050898
Wongsa T, Chankaew S, Monkham T, Sanitchon J. Broad-Spectrum Resistance and Monogenic Inheritance of Bacterial Blight Resistance in an Indigenous Upland Rice Germplasm ULR207. Agronomy. 2024; 14(5):898. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050898
Chicago/Turabian StyleWongsa, Tanawat, Sompong Chankaew, Tidarat Monkham, and Jirawat Sanitchon. 2024. "Broad-Spectrum Resistance and Monogenic Inheritance of Bacterial Blight Resistance in an Indigenous Upland Rice Germplasm ULR207" Agronomy 14, no. 5: 898. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050898