
Clinical and Biologic Correlates of ADORA2A Transcriptomic Expression in Cancer
 , ,
, ,
Abstract
:1. Introduction
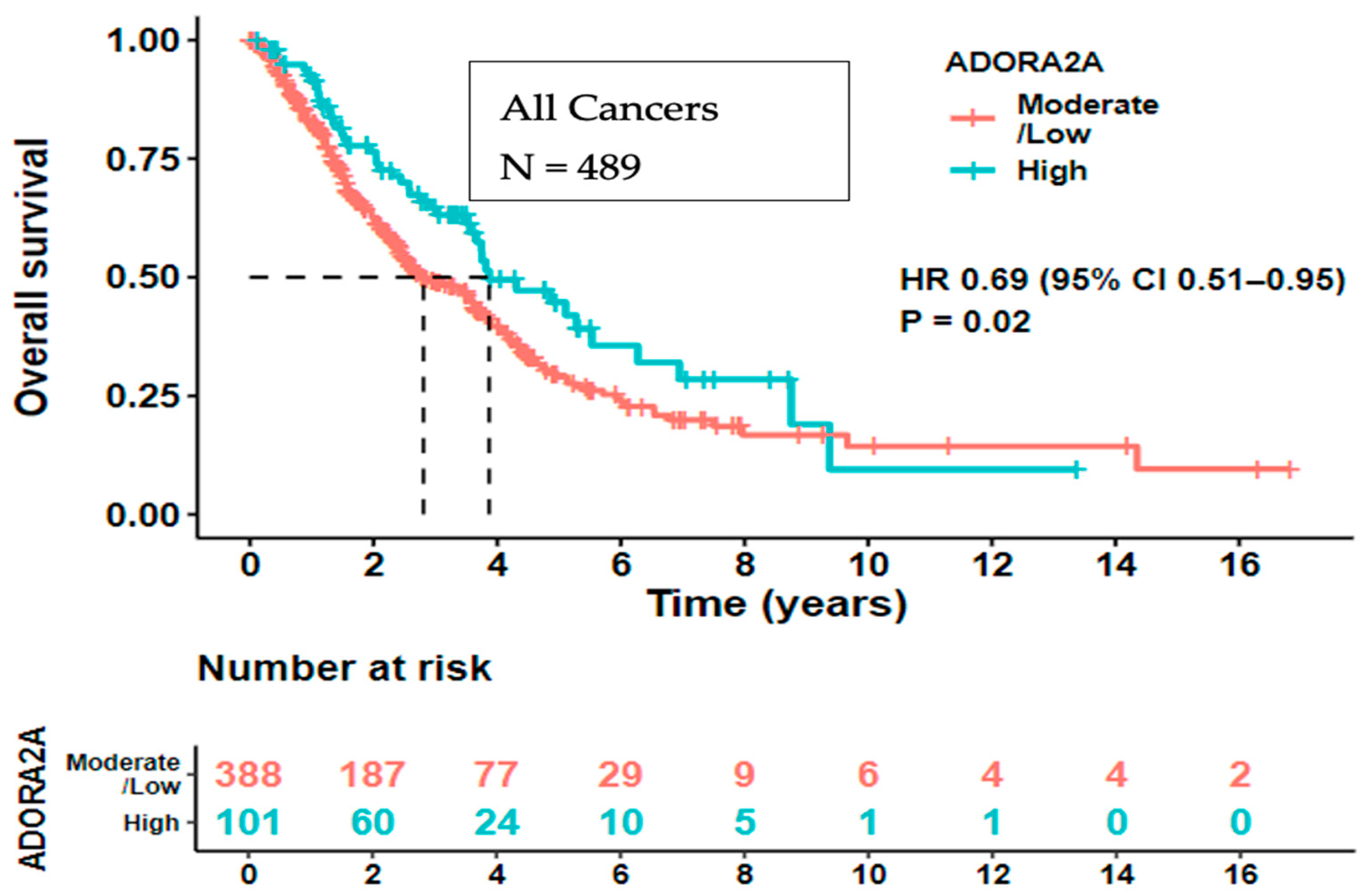
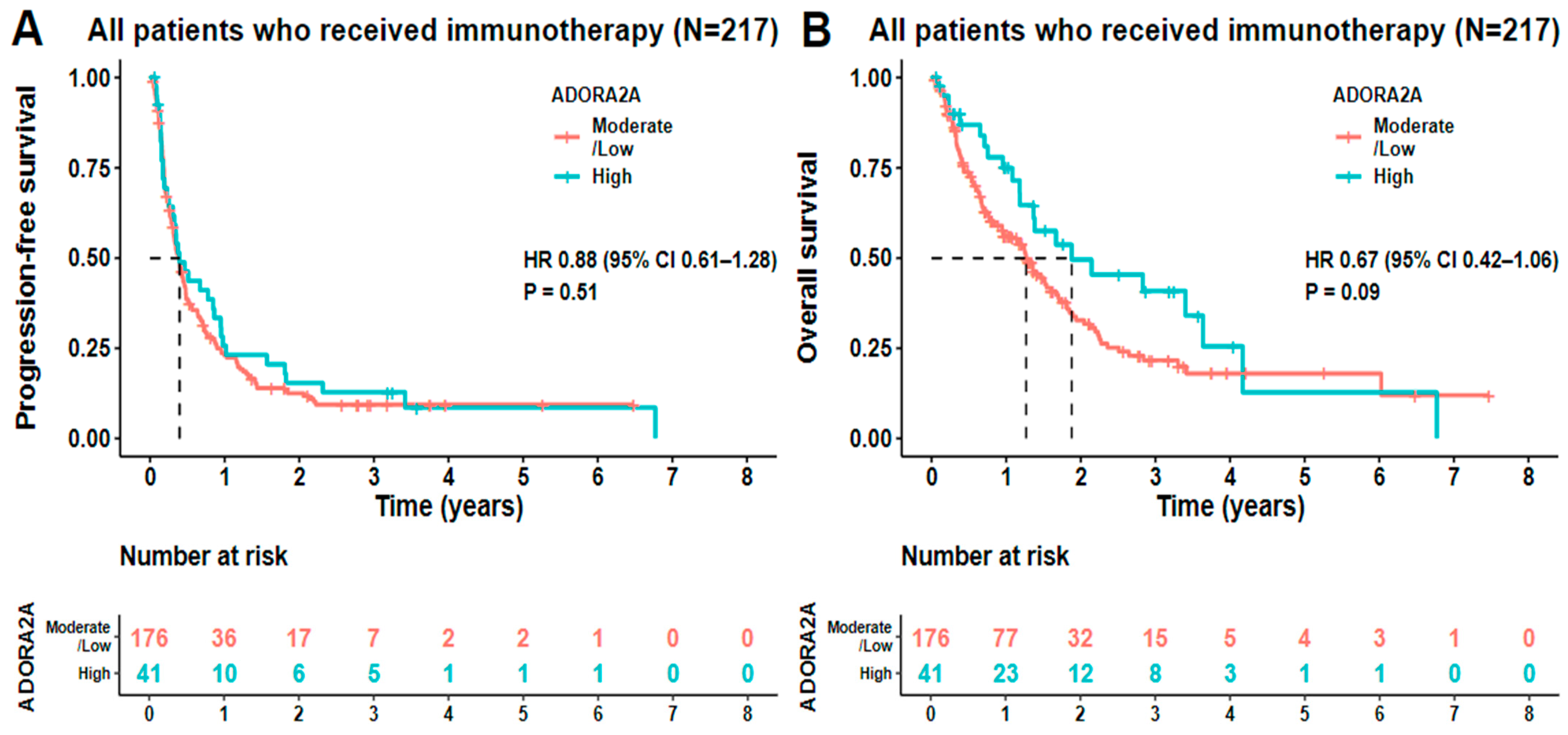
2. Results
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Transcriptomics
4.3. Tumor Mutational Burden (TMB)
4.4. Data Collection and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Klinger, M.; Freissmuth, M.; Nanoff, C. Adenosine receptors: G protein-mediated signalling and the role of accessory proteins. Cell Signal. 2002, 14, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Libert, F.; Passage, E.; Parmentier, M.; Simons, M.J.; Vassart, G.; Mattei, M.G. Chromosomal mapping of A1 and A2 adenosine receptors, VIP receptor, and a new subtype of serotonin receptor. Genomics 1991, 11, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, D.; Young, A.; Teng, M.W.L.; Smyth, M.J. Targeting immunosuppressive adenosine in cancer. Nat. Rev. Cancer 2017, 17, 709–724. [Google Scholar] [CrossRef]
- Blay, J.; White, T.D.; Hoskin, D.W. The extracellular fluid of solid carcinomas contains immunosuppressive concentrations of adenosine. Cancer Res. 1997, 57, 2602–2605. [Google Scholar] [PubMed]
- Leone, R.D.; Lo, Y.C.; Powell, J.D. A2aR antagonists: Next generation checkpoint blockade for cancer immunotherapy. Comput. Struct. Biotechnol. J. 2015, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Linden, J.; Muller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Sinha, D.; Barkauskas, D.; Young, A.; Kalimutho, M.; Stannard, K.; Caramia, F.; Haibe-Kains, B.; Stagg, J.; Khanna, K.K.; et al. Adenosine 2B Receptor Expression on Cancer Cells Promotes Metastasis. Cancer Res. 2016, 76, 4372–4382. [Google Scholar] [CrossRef]
- Ohta, A.; Gorelik, E.; Prasad, S.J.; Ronchese, F.; Lukashev, D.; Wong, M.K.; Huang, X.; Caldwell, S.; Liu, K.; Smith, P.; et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13132–13137. [Google Scholar] [CrossRef] [PubMed]
- Beavis, P.A.; Milenkovski, N.; Henderson, M.A.; John, L.B.; Allard, B.; Loi, S.; Kershaw, M.H.; Stagg, J.; Darcy, P.K. Adenosine Receptor 2A Blockade Increases the Efficacy of Anti-PD-1 through Enhanced Antitumor T-cell Responses. Cancer Immunol. Res. 2015, 3, 506–517. [Google Scholar] [CrossRef]
- Willingham, S.B.; Ho, P.Y.; Hotson, A.; Hill, C.; Piccione, E.C.; Hsieh, J.; Liu, L.; Buggy, J.J.; McCaffery, I.; Miller, R.A. A2AR Antagonism with CPI-444 Induces Antitumor Responses and Augments Efficacy to Anti-PD-(L)1 and Anti-CTLA-4 in Preclinical Models. Cancer Immunol. Res. 2018, 6, 1136–1149. [Google Scholar] [CrossRef]
- Ohta, A. A Metabolic Immune Checkpoint: Adenosine in Tumor Microenvironment. Front. Immunol. 2016, 7, 109. [Google Scholar] [CrossRef]
- Sek, K.; Molck, C.; Stewart, G.D.; Kats, L.; Darcy, P.K.; Beavis, P.A. Targeting Adenosine Receptor Signaling in Cancer Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3837. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Diao, L.; Yang, Y.; Yi, X.; Rodriguez, B.L.; Li, Y.; Villalobos, P.A.; Cascone, T.; Liu, X.; Tan, L.; et al. CD38-Mediated Immunosuppression as a Mechanism of Tumor Cell Escape from PD-1/PD-L1 Blockade. Cancer Discov. 2018, 8, 1156–1175. [Google Scholar] [CrossRef]
- Yang, R.; Elsaadi, S.; Misund, K.; Abdollahi, P.; Vandsemb, E.N.; Moen, S.H.; Kusnierczyk, A.; Slupphaug, G.; Standal, T.; Waage, A.; et al. Conversion of ATP to adenosine by CD39 and CD73 in multiple myeloma can be successfully targeted together with adenosine receptor A2A blockade. J. Immunother. Cancer 2020, 8, e000610. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Mayer, A. Hypoxia-Driven Adenosine Accumulation: A Crucial Microenvironmental Factor Promoting Tumor Progression. Adv. Exp. Med. Biol. 2016, 876, 177–183. [Google Scholar]
- Ma, X.L.; Shen, M.N.; Hu, B.; Wang, B.L.; Yang, W.J.; Lv, L.H.; Wang, H.; Zhou, Y.; Jin, A.L.; Sun, Y.F.; et al. CD73 promotes hepatocellular carcinoma progression and metastasis via activating PI3K/AKT signaling by inducing Rap1-mediated membrane localization of P110beta and predicts poor prognosis. J. Hematol. Oncol. 2019, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Ngiow, S.F.; Barkauskas, D.S.; Sult, E.; Hay, C.; Blake, S.J.; Huang, Q.; Liu, J.; Takeda, K.; Teng, M.W.L.; et al. Co-inhibition of CD73 and A2AR Adenosine Signaling Improves Anti-tumor Immune Responses. Cancer Cell 2016, 30, 391–403. [Google Scholar] [CrossRef]
- Sun, C.; Wang, B.; Hao, S. Adenosine-A2A Receptor Pathway in Cancer Immunotherapy. Front. Immunol. 2022, 13, 837230. [Google Scholar] [CrossRef]
- Allard, B.; Jacoberger-Foissac, C.; Cousineau, I.; Bareche, Y.; Buisseret, L.; Chrobak, P.; Allard, D.; Pommey, S.; Ah-Pine, F.; Duquenne, S.; et al. Adenosine A2A receptor is a tumor suppressor of NASH-associated hepatocellular carcinoma. Cell Rep. Med. 2023, 4, 101188. [Google Scholar] [CrossRef]
- Patel, S.P.; Kurzrock, R. PD-L1 Expression as a Predictive Biomarker in Cancer Immunotherapy. Mol. Cancer Ther. 2015, 14, 847–856. [Google Scholar] [CrossRef]
- Nesline, M.K.; Previs, R.A.; Dy, G.K.; Deng, L.; Lee, Y.H.; DePietro, P.; Zhang, S.; Meyers, N.; Severson, E.; Ramkissoon, S.; et al. PD-L1 Expression by RNA-Sequencing in Non-Small Cell Lung Cancer: Concordance with Immunohistochemistry and Associations with Pembrolizumab Treatment Outcomes. Cancers 2023, 15, 4789. [Google Scholar] [CrossRef] [PubMed]
- Bevins, N.J.; Okamura, R.; Montesion, M.; Adashek, J.J.; Goodman, A.M.; Kurzrock, R. Tumor Infiltrating Lymphocyte Expression of PD-1 Predicts Response to Anti-PD-1/PD-L1 Immunotherapy. J. Immunother. Precis. Oncol. 2022, 5, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Ruppin, E. Multiomics Prediction of Response Rates to Therapies to Inhibit Programmed Cell Death 1 and Programmed Cell Death 1 Ligand 1. JAMA Oncol. 2019, 5, 1614–1618. [Google Scholar] [CrossRef] [PubMed]
- Kamai, T.; Kijima, T.; Tsuzuki, T.; Nukui, A.; Abe, H.; Arai, K.; Yoshida, K.I. Increased expression of adenosine 2A receptors in metastatic renal cell carcinoma is associated with poorer response to anti-vascular endothelial growth factor agents and anti-PD-1/Anti-CTLA4 antibodies and shorter survival. Cancer Immunol. Immunother. 2021, 70, 2009–2021. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.; Hotson, A.; Powderly, J.D.; Sznol, M.; Heist, R.S.; Choueiri, T.K.; George, S.; Hughes, B.G.M.; Hellmann, M.D.; Shepard, D.R.; et al. Adenosine 2A Receptor Blockade as an Immunotherapy for Treatment-Refractory Renal Cell Cancer. Cancer Discov. 2020, 10, 40–53. [Google Scholar] [CrossRef]
- Sitkovsky, M.V. Lessons from the A2A Adenosine Receptor Antagonist-Enabled Tumor Regression and Survival in Patients with Treatment-Refractory Renal Cell Cancer. Cancer Discov. 2020, 10, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Yin, S.; To, K.K.W.; Fu, L. CD39/CD73/A2AR pathway and cancer immunotherapy. Mol. Cancer 2023, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Liu, C.; Wang, Z.; Wu, W.; Zhang, N.; Zhang, L.; Hu, J.; Luo, P.; Zhang, J.; et al. Immune checkpoint modulators in cancer immunotherapy: Recent advances and emerging concepts. J. Hematol. Oncol. 2022, 15, 111. [Google Scholar] [CrossRef] [PubMed]
- Nishizaki, D.; Kurzrock, R.; Miyashita, H.; Adashek, J.J.; Lee, S.; Nikanjam, M.; Eskander, R.N.; Patel, H.; Botta, G.P.; Nesline, M.K.; et al. Viewing the immune checkpoint VISTA: Landscape and outcomes across cancers. ESMO Open 2024, 9, 102942. [Google Scholar] [CrossRef]
- Kato, S.; Okamura, R.; Kumaki, Y.; Ikeda, S.; Nikanjam, M.; Eskander, R.; Goodman, A.; Lee, S.; Glenn, S.T.; Dressman, D.; et al. Expression of TIM3/VISTA checkpoints and the CD68 macrophage-associated marker correlates with anti-PD1/PDL1 resistance: Implications of immunogram heterogeneity. Oncoimmunology 2020, 9, 1708065. [Google Scholar] [CrossRef]
- Turcotte, M.; Spring, K.; Pommey, S.; Chouinard, G.; Cousineau, I.; George, J.; Chen, G.M.; Gendoo, D.M.A.; Haibe-Kains, B.; Karn, T.; et al. CD73 is associated with poor prognosis in high-grade serous ovarian cancer. Cancer Res. 2015, 75, 4494–4503. [Google Scholar] [CrossRef]
- Sicklick, J.K.; Kato, S.; Okamura, R.; Schwaederle, M.; Hahn, M.E.; Williams, C.B.; De, P.; Krie, A.; Piccioni, D.E.; Miller, V.A.; et al. Molecular profiling of cancer patients enables personalized combination therapy: The I-PREDICT study. Nat. Med. 2019, 25, 744–750. [Google Scholar] [CrossRef]
- Adashek, J.J.; Goloubev, A.; Kato, S.; Kurzrock, R. Missing the target in cancer therapy. Nat. Cancer 2021, 2, 369–371. [Google Scholar] [CrossRef]
- Conroy, J.M.; Pabla, S.; Glenn, S.T.; Burgher, B.; Nesline, M.; Papanicolau-Sengos, A.; Andreas, J.; Giamo, V.; Lenzo, F.L.; Hyland, F.C.L.; et al. Analytical Validation of a Next-Generation Sequencing Assay to Monitor Immune Responses in Solid Tumors. J. Mol. Diagn. 2018, 20, 95–109. [Google Scholar] [CrossRef]
- Adashek, J.J.; Kato, S.; Nishizaki, D.; Miyashita, H.; De, P.; Lee, S.; Pabla, S.; Nesline, M.; Conroy, J.M.; DePietro, P.; et al. LAG-3 transcriptomic expression patterns across malignancies: Implications for precision immunotherapeutics. Cancer Med. 2023, 12, 13155–13166. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Kato, S.; Nesline, M.K.; Conroy, J.M.; DePietro, P.; Pabla, S.; Kurzrock, R. Indoleamine 2,3-dioxygenase (IDO) inhibitors and cancer immunotherapy. Cancer Treat. Rev. 2022, 110, 102461. [Google Scholar] [CrossRef]
- Jou, J.; Kato, S.; Miyashita, H.; Thangathurai, K.; Pabla, S.; DePietro, P.; Nesline, M.K.; Conroy, J.M.; Rubin, E.; Eskander, R.N.; et al. Cancer-Immunity Marker RNA Expression Levels across Gynecologic Cancers: Implications for Immunotherapy. Mol. Cancer Ther. 2023, 22, 1352–1362. [Google Scholar] [CrossRef]
- Miyashita, H.; Kurzrock, R.; Bevins, N.J.; Thangathurai, K.; Lee, S.; Pabla, S.; Nesline, M.; Glenn, S.T.; Conroy, J.M.; DePietro, P.; et al. T-cell priming transcriptomic markers: Implications of immunome heterogeneity for precision immunotherapy. NPJ Genom. Med. 2023, 8, 19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Univariable | Multivariable | Comment | |||
|---|---|---|---|---|---|---|
| Condition (N = Total No. of Cases) | Proportion of High ADORA2A among All Cases with (%) | Odds Ratio (95% CI) | p-Value | Odds Ratio (95% CI) | p-Value | |
| Gender Men (N = 204) Women (N = 310) | Men (46/204, 22.5%) Women (61/310, 19.6%) | 1.19 (0.77–1.83) | 0.433 | |||
| Age above 61 (N = 256) and below median of 61 years (N = 258) | Age above median 61 (60/256, 23.4%) Age below median 61 (72/258, 27.9%) | 0.82 (0.53–1.25) | 0.351 | |||
| + PD-L1 (CPS ≥ 1%) IHC (N = 156) PD-L1 (<1%) IHC (N = 357) | + PDL1 (≥1%) IHC (31/156, 19.8%) PDL1 (<1%) IHC (76/357, 21.2%) | 0.92 (0.57–1.45) | 0.716 | One patient was missing PD-L1 IHC evaluation | ||
| High 1 CTLA-4 (N = 87) Low/Moderate CTLA-4 (N = 427) | High 1 CTLA4 (33/87, 37.9%) Low/Moderate CTLA4 (74/427, 17.3%) | 2.92 (1.76–4.80) | <0.001 | 0.79 (0.37–1.64) | 0.538 | |
| High 1 LAG-3 (N = 116) Low/Moderate LAG-3 (N = 398) | High 1 LAG3 (41/116, 35.3%) Low/Moderate LAG3 (66/398, 16.5%) | 2.75 (1.72–4.36) | <0.001 | 0.86 (0.42–1.70) | 0.665 | |
| High 1 PD-1 (N = 93) Low/Moderate PD-1 (N = 421) | High 1 PD-1 (41/93, 44.08%) Low/Moderate PD-1 (66/421, 15.6%) | 4.24 (2.60–6.90) | <0.001 | 2.55 (1.19–5.45) | 0.015 | High ADORA2A RNA (≥75th percentile rank) was positively associated with high PD1 |
| High 1 PD-L1 (N = 67) Low/Moderate PD-L1 (N = 447) | High 1 PD-L1 (25/67, 37.3%) Low/Moderate PD-L1 (82/447, 18.3%) | 2.65 (1.51–4.57) | <0.001 | 0.198 (0.74–3.86) | 0.198 | |
| High 1 PD-L2 (N = 100) Low/Moderate PD-L2 (N = 414) | High 1 PD-L2 (37/100, 37.0%) Low/Moderate PD-L2 (70/414, 16.9%) | 2.89 (1.78–4.66) | <0.001 | 1.07 (0.51–2.21) | 0.858 | |
| High 1 TIM-3 (N = 90) Low/Moderate TIM-3 (N = 424) | High 1 TIM3 (28/90, 31.1%) Low/Moderate TIM3 (79/424, 18.6%) | 1.97 (1.17–3.26) | 0.009 | 0.50 (0.23–1.05) | 0.073 | |
| High 1 VISTA (N = 166) Low/Moderate VISTA (N = 348) | High 1 VISTA (59/166, 35.5%) Low/Moderate VISTA (48/348, 13.7%) | 3.45 (2.22–5.37) | <0.001 | 3.05 (1.75–5.36) | 0.001 | High ADORA2A RNA (≥75th percentile rank) was positively associated with high VISTA |
| High 1 CD38 (N = 79) Low/Moderate CD38 (N = 435) | High 1 CD38 (35/79, 44.3%) Low/Moderate CD38 (72/435, 16.5%) | 4.01 (2.40–6.69) | <0.001 | 2.24 (1.07–4.63) | 0.031 | High ADORA2A RNA (≥75th percentile rank) was positively associated with high CD38 |
| High 1 CD39 (N = 131) Low/Moderate CD39 (N = 383) | High 1 CD39 (54/131, 41.2%) Low/Moderate CD39 (53/383, 13.8%) | 4.37 (2.78–6.89) | <0.001 | 3.54 (1.93 -6.54) | <0.001 | High ADORA2A RNA (≥75th percentile rank) was positively associated with high CD39 |
| High 1 IDO1 (N = 91) Low/Moderate IDO1 (N = 423) | High 1 IDO1 (19/91, 20.8%) Low/Moderate IDO1 (88/423, 20.8%) | 1.00 (0.56–1.72) | 0.987 | |||
| Microsatellite unstable (N = 15) Microsatellite stable (N = 425) | Unstable (3/15, 20.0%) Stable (85/425, 20.0%) | 1.00 (0.22–3.23) | 0.999 | |||
| TMB (N = 450 total samples) a TMB ≥ 10 mutations/megabase (N = 33) TMB < 10 mutations/megabase (N = 417) | TMB ≥10 mutations/megabase (8/33, 24.2%) TMB < 10 mutations/megabase (81/417, 19.4%) | 1.33 (0.54–2.93) | 0.505 | |||
| Neuroendocrine cancer (N = 15) | NET (11/15, 73%) Not NET (96/499, 19.2%) | 11.5 (3.86–42.4) | <0.001 | 19.3 (5.53–80.4) | <0.001 | High ADORA2A RNA (≥75th percentile rank) was positively associated with neuroendocrine cancer |
| Sarcoma (N = 24) | Sarcoma (9/24, 37.5%) Not Sarcoma (98/490, 20.0%) | 2.40 (0.98–5.56) | 0.045 | 3.34 (1.21–8.91) | 0.017 | High ADORA2A RNA (≥75th percentile rank) was positively associated with sarcoma |
| Breast cancer (N = 49) | Breast (19/49, 38.8%) Not Breast (88/465, 18.9%) | 2.71 (1.44–5.01) | 0.002 | 4.43 (2.01–9.78) | <0.001 | High ADORA2A RNA (≥75th percentile rank) was positively associated with breast cancer |
| Pancreas cancer (N = 55) | Pancreas (13/55, 23.6%) Not Pancreas (94/459, 20.4%) | 1.20 (0.60–2.27) | 0.586 | |||
| Carcinoma Unknown Primary or CUP (N = 13) | CUP (3/13, 23.1%) Not CUP (104/501, 20.7%) | 1.15 (0.25–3.82) | 0.839 | |||
| Esophageal cancer (N = 17) | Esophageal ca (2/17, 11.8%) Not esophageal ca (105/497, 21.1%) | 0.50 (0.08–1.80) | 0.359 | |||
| Lung cancer (N = 20) | Lung ca (3/20, 15%) Not Lung CA (104/494, 21%) | 0.66 (0.15–2.02) | 0.516 | |||
| Head and neck cancer (N = 12) | Head & Neck cancer (2/12, 16.7%) Not head & neck cancer (105/502, 20.9%) | 0.76 (0.12–2.92) | 0.721 | |||
| Stomach cancer (N = 25) | Stomach ca (5/25, 20%) Not stomach ca (102/489, 20.8%) | 0.95 (0.31–2.41) | 0.918 | |||
| Liver and bile duct cancer (N = 19) | Liver ca (4/19, 21.1%) Not liver Ca (103/495, 20.8%) | 1.01 (0.28–2.87) | 0.979 | |||
| Uterine cancer (N = 24) | Uterine cancer (3/24, 12.5%) Not uterine cancer (104/490, 21.2%) | 0.53 (0.12–1.58) | 0.312 | |||
| Small intestine cancer (N = 12) | Small intestine (2/12, 16.7%) Not small intestine cancer (105/502, 20.9%) | 0.76 (0.12–2.92) | 0.721 | |||
| Colorectal cancer (N = 140) | Colorectal (15/140, 10.7%) Not colorectal (92/374, 24.5%) | 0.37 (0.20–0.64) | 0.001 | 0.49 (0.24–0.96) | 0.044 | High ADORA2A RNA (≥75th percentile rank) was negatively associated with colorectal cancer |
| Ovarian cancer (N = 43) | Ovarian cancer (2/43, 4.7%) Not ovarian cancer (105/471, 22.2%) | 0.17 (0.03–0.57) | 0.005 | 0.17(0.02–0.67) | 0.028 | High ADORA2A RNA (≥75th percentile rank) was negatively associated with ovarian cancer |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shreenivas, A.; Nishizaki, D.; Lee, S.; Pabla, S.; Nesline, M.; Conroy, J.M.; DePietro, P.; Kato, S.; Kurzrock, R. Clinical and Biologic Correlates of ADORA2A Transcriptomic Expression in Cancer. Int. J. Mol. Sci. 2024, 25, 4742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094742
Shreenivas A, Nishizaki D, Lee S, Pabla S, Nesline M, Conroy JM, DePietro P, Kato S, Kurzrock R. Clinical and Biologic Correlates of ADORA2A Transcriptomic Expression in Cancer. International Journal of Molecular Sciences. 2024; 25(9):4742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094742
Chicago/Turabian StyleShreenivas, Aditya, Daisuke Nishizaki, Suzanna Lee, Sarabjot Pabla, Mary Nesline, Jeffrey M. Conroy, Paul DePietro, Shumei Kato, and Razelle Kurzrock. 2024. "Clinical and Biologic Correlates of ADORA2A Transcriptomic Expression in Cancer" International Journal of Molecular Sciences 25, no. 9: 4742. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094742