
The Glucose–Succinate Pathway: A Crucial Anaerobic Metabolic Pathway in the Scallop Chlamys farreri Experiencing Heat Stress
Abstract
:1. Introduction
2. Results
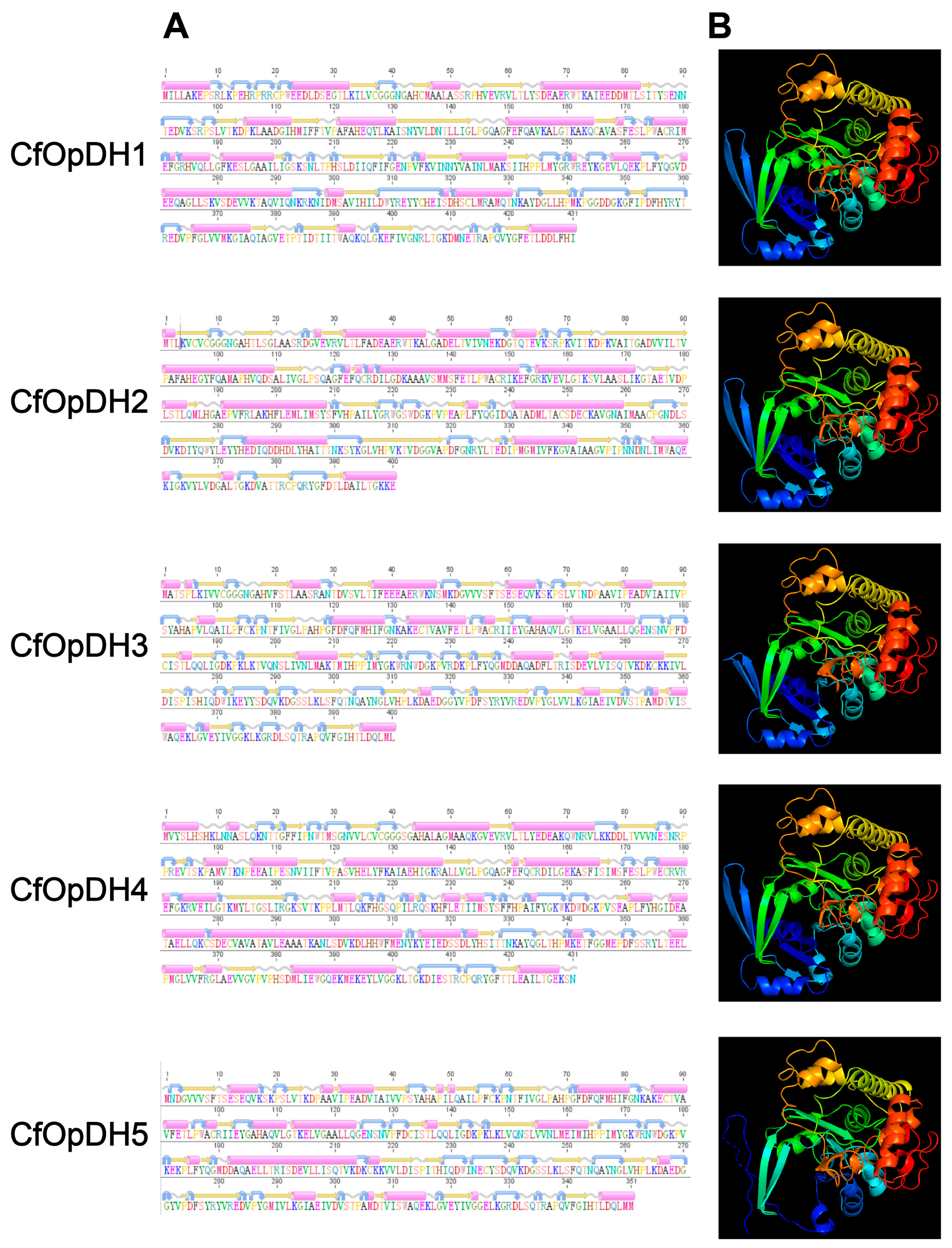
2.1. Gene Identification and Sequence Analysis
2.2. Phylogenetic Analysis
2.3. Spatiotemporal Expression
2.4. Expression Profiles of CfOpDHs, CfPEPCKs, and CfASTs in Response to Heat Stress
3. Discussion
3.1. Glucose–Lactate Pathway
3.2. Glucose–Opine Pathway
3.3. Aspartate–Succinate Pathway
3.4. Glucose–Succinate Pathway
4. Materials and Methods
4.1. Genome-Wide Identification and Sequence Analysis of LDH, OpDH, PEPCK, and AST Genes in C. farreri
4.2. Phylogenetic Analysis
4.3. Spatiotemporal Expression of OpDH, PEPCK, and AST Genes in C. farreri
4.4. Expression Analysis of CfOpDHs, CfPEPCKs, and CfASTs under Heat Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, X.; Luo, Y. Scallops and Scallop Aquaculture in China. Dev. Aquac. Fish. Sci. 2016, 40, 937–952. [Google Scholar]
- Venugopal, V.; Gopakumar, K. Shellfish: Nutritive value, health benefits, and consumer safety. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1219–1242. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.C.; Fan, Y.; Baker, G.L. Nutritional Value and Food Safety of Bivalve Molluscan Shellfish. J. Shellfish Res. 2018, 37, 695–708. [Google Scholar] [CrossRef]
- Huajing Industrial Research Institute. The Main Species, Breeding Area, Breeding Output, Import and Export Situation and Market Prospect of Scallop in China Were Analyzed; Huajing Industrial Research Institute: Huajing, China, 2022. [Google Scholar]
- Carroll, J.M.; Furman, B.T.; Tettelbach, S.T.; Peterson, B.J. Balancing the edge effects budget: Bay scallop settlement and loss along a seagrass edge. Ecology 2012, 93, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Giantsis, I.A.; Vratsistas, A.; Makri, S.; Pappa, A.Z.; Drosopoulou, E.; Anestis, A.; Mavridou, E.; Exadactylos, A.; Vafidis, D.; et al. Correlation between intermediary metabolism, Hsp gene expression, and oxidative stress-related proteins in long-term thermal-stressed Mytilus galloprovincialis. Am. J. Physiol. Reg. Integr. Comp. Physiol. 2020, 319, R264–R281. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Ding, J.; Ben, Y.; Chang, Y. Effects of high temperature on survival rate, oxygen consumption rate, and ammonia excretion rate of Mizuhopecten yessoensis. J. Dalian Ocean Univ. 2013, 28, 138–142. [Google Scholar]
- Wang, J.; Jiang, Z.; Tang, Q. Study on physiological energetics of Farrer’s scallop (Chlamys farreri). Mar. Fish. Res. 2004, 25, 46–53. [Google Scholar]
- Xu, D.; Zhang, J.; Wang, W.; Dong, M.; Wang, Z. Research on feeding physiology of scallop Patinopecten yessoensis. Prog. Fish. Sci. 2010, 31, 85–91. [Google Scholar]
- Falfushynska, H.I.; Phan, T.; Sokolova, I.M. Long-term acclimation to different thermal regimes affects molecular responses to heat stress in a freshwater clam Corbicula fluminea. Sci. Rep. 2016, 6, 39476. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Han, G.; Zhang, S.; Dong, Y. Anaerobic metabolism and thermal tolerance: The importance of opine pathways on survival of a gastropod after cardiac dysfunction. Integr. Zool. 2017, 12, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Harcet, M.; Perina, D.; Pleše, B. Opine dehydrogenases in marine invertebrates. Biochem. Genet. 2013, 51, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Keeling, R.E.; Körtzinger, A.; Gruber, N. Ocean deoxygenation in a warming world. Annu. Rev. Mar. Sci. 2010, 2, 199–229. [Google Scholar] [CrossRef] [PubMed]
- Schmidtko, S.; Stramma, L.; Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nature 2017, 542, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Liang, Y.S.; Gu, Y.; Yao, F.C.; Liu, Y.F.; Zhang, K.X.; Song, F.B.; Sun, J.L.; Luo, J. Different reoxygenation rates induce different metabolic, apoptotic and immune responses in Golden Pompano (Trachinotus blochii) after hypoxic stress. Fish Shellfish Immunol. 2023, 135, 108640. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Invertebrate and vertebrate pathways of anaerobic metabolism: Evolutionary considerations. J. Geol. Soc. 1983, 140, 27–37. [Google Scholar] [CrossRef]
- Huckabee, E.; Hospitals, M.M.; Pyruvate, D.I. Relationships of pyruvate and lactate during anaerobic metabolism. I. Effects of infusion of pyruvate or glucose and of hyperventilation. J. Clin. Investig. 1958, 37, 244–254. [Google Scholar] [CrossRef]
- Schöttler, U. On the Anaerobic Metabolism of Three Species of Nereis (Annelida). Mar. Ecol. Prog. Ser. 1979, 1, 249–254. [Google Scholar] [CrossRef]
- Regnouf, F.; van Thoai, N. Octopine and lactate dehydrogenases in mollusc muscles. Comp. Biochem. Physiol. 1970, 32, 411–416. [Google Scholar] [CrossRef]
- Zheng, Y.; Tian, Y.; Liu, Y.; Liu, J. Research on Opine dehydrogenase in commercial marine mollusks: A review. J. Dalian Ocean Univ. 2019, 34, 296–302. [Google Scholar]
- Hochachka, P.W. Living without Oxygen; Harvard University Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Livingstone, D.R. Origins and evolution of pathways of anaerobic metabolism in the animal kingdom. Am. Zool. 1991, 31, 522–534. [Google Scholar] [CrossRef]
- De Zwaan, A. Anaerobic energy metabolism in bivalve mollusks. In Oceanography and Marine Biology: An Annual Review; Barnes, H.B., Ed.; Routledge: London, UK, 1977; Volume 15, pp. 103–187. [Google Scholar]
- Ebberink, R.H.M.; Zurburg, W.; Zandee, D.I. The energy demand of the posterior adductor muscle of Mytilus edulis in catch during exposure to air. Mar. Biol. Lets. 1979, 1, 23–31. [Google Scholar]
- Fields, J.H.A. Alternatives to lactic acid: Possible advantages. J. Exp. Zool. 1983, 228, 445–457. [Google Scholar] [CrossRef]
- Livingstone, D.R.; De Zwaan, A.; Leopold, M.; Marteijn, E. Studies on the phylogenetic distribution of pyruvate oxidoreductases. Biochem. Syst. Ecol. 1983, 11, 415–425. [Google Scholar] [CrossRef]
- Livingstone, D.R.; De Zwaan, A.; Thompson, R.J. Aerobic metabolism, octopine production and phosphoarginine as sources of energy in the phasic and catch adductor muscles of the giant scallop Placopecten magellanicus during swimming and the subsequent recovery period. Comp. Biochem. Physiol. B 1981, 70, 35–44. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Bock, C.; Pörtner, H.O. Resistance to freshwater exposure in White Sea Littorina spp. I: Anaerobic metabolism and energetics. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2000, 170, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Li, M.; Cai, Y.; Qi, L.; Yuan, C.; Tian, Y. Examination of the effects of dry storage stress on mitochondrial energy synthesis in the scallop Mizuhopecten yessoensis. Aquaculture 2023, 575, 739770. [Google Scholar] [CrossRef]
- Reyes-Ramos, C.A.; Peregrino-Uriarte, A.B.; Cota-Ruiz, K.; Valenzuela-Soto, E.M.; Leyva-Carrillo, L.; Yepiz-Plascencia, G. Phosphoenolpyruvate carboxykinase cytosolic and mitochondrial isoforms are expressed and active during hypoxia in the white shrimp Litopenaeus vannamei. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 226, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rojas, B.E.; Hartman, M.D.; Figueroa, C.M.; Leaden, L.; Podestá, F.E.; Iglesias, A.A. Biochemical characterization of phosphoenolpyruvate carboxykinases from Arabidopsis thaliana. Biochem. J. 2019, 476, 2939–2952. [Google Scholar] [CrossRef]
- Utter, M.F.; Kurahashi, K. Mechanism of action of oxalacetic carboxylase from liver. J. Am. Chem. Soc. 1953, 75, 758. [Google Scholar] [CrossRef]
- Gäde, G.; Grieshaber, M.K. Pyruvate reductases catalyze the formation of lactate and opines in anaerobic invertebrates. Comp. Biochem. Physiol. Part B Comp. Biochem. 1986, 83, 255–272. [Google Scholar] [CrossRef]
- Hochachka, P.W. Fuels and pathways as designed systems for support of muscle work. J. Exp. Biol. 1985, 115, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.H.; de Bont, A.M.T.; Janus, J.; Wusman, T.C.M. Time dependent changes and tissue specificities in the accumulation of anaerobic fermentation products in the sea mussel Mytilus edulis L. Comp. Biochem. Physiol. Part B Comp. Biochem. 1997, 58, 81–87. [Google Scholar] [CrossRef]
- Kluytmans, J.H.; De Zwaan, A. Production of volatile fatty acids in bivalved molluscs. Biochem. Soc. Trans. 1976, 4, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Wongso, S.; Ushio, H.; Yamanaka, H. Glycolytic enzymes in the tissues of three species of scallop (Bivalvia: Pectinidae). Fish. Sci. 1999, 65, 123–128. [Google Scholar] [CrossRef]
- Shepherd, J.G.; Brewer, P.G.; Oschlies, A.; Watson, A.J. Ocean ventilation and deoxygenation in a warming world: Introduction and overview. Philos. Trans. Ser. A Math. Phys. Eng. Sci. 2017, 375, 20170240. [Google Scholar] [CrossRef] [PubMed]
- Boardman, L.; Sørensen, J.G.; Koštál, V.; Šimek, P.; Terblanche, J.S. Cold tolerance is unaffected by oxygen availability despite changes in anaerobic metabolism. Sci. Rep. 2016, 6, 32856. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, G.E.; Todgham, A.E. Living in the now: Physiological mechanisms to tolerate a rapidly changing environment. Annu. Rev. Physiol. 2010, 72, 127–145. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation; Princeton University Press: Princeton, NJ, USA, 1984. [Google Scholar]
- Kimura, T.; Nakano, T.; Yamaguchi, T.; Sato, M.; Ogawa, T.; Muramoto, K.; Yokoyama, T.; Kan-No, N.; Nagahisa, E.; Janssen, F.; et al. Complementary DNA cloning and molecular evolution of opine dehydrogenases in some marine invertebrates. Mar. Biotechnol. 2004, 6, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.T.; Rossmann, M.G. Comparison of super-secondary structures in proteins. J. Mol. Biol. 1973, 76, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Takeuchi, M.; Kanno, N.; Nagahisa, E.; Sato, Y. Distribution of opine dehydrogenases and lactate dehydrogenase activities in marine animals. Comp. Biochem. Physiol. Part B Comp. Biochem. 1993, 106, 955–960. [Google Scholar] [CrossRef]
- Vázquez-Dorado, S.; Sanjuan, A.; Comesaña, Á.S.; de Carlos, A. Identification of octopine dehydrogenase from Mytilus galloprovincialis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 160, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Endo, N.; Matsu-ura, H.; Kan-no, N. cDNA cloning and primary structure comparison of tauropine dehydrogenase and β-alanopine dehydrogenase from the limpet Cellana grata. Fish. Sci. 2009, 75, 1471–1479. [Google Scholar] [CrossRef]
- Choi, S.H.; Jee, B.Y.; Lee, S.J.; Cho, M.Y.; Lee, S.J.; Kim, J.W.; Jeong, H.D.; Kim, K.H. Effects of RNA interference-mediated knock-down of hypoxia-inducible factor-α on respiratory burst activity of the Pacific oyster Crassostrea gigas hemocytes. Fish Shellfish Immunol. 2013, 35, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Chille, E.; Strand, E.; Neder, M.; Schmidt, V.; Sherman, M.; Mass, T.; Putnam, H. Developmental series of gene expression clarifies maternal mRNA provisioning and maternal-to-zygotic transition in a reef-building coral. BMC Genom. 2021, 22, 815. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, B.; Xu, Q.; Xu, Q.; Wang, Q. The landscape of RNA Pol II binding reveals a stepwise transition during ZGA. Nature 2020, 587, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Vastenhouw, N.L.; Zhang, Y.; Woods, I.G.; Imam, F.; Regev, A.; Liu, X.S.; Rinn, J.; Schier, A.F. Chromatin signature of embryonic pluripotency is established during genome activation. Nature 2010, 464, 922–926. [Google Scholar] [CrossRef]
- Jia, Q.; Ni, Y.; Min, S.; Ming, L.; Qian, Y.; Cen, X.; Wang, J.; Tong, X. The ontogenesis of catabolic abilities and energy metabolism during endogenous nutritional periods of tongue sole, Cynoglossus semilaevis. J. Fish Biol. 2021, 99, 1708–1718. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Obluchinskaya, E.D.; Gorshenina, E.V.; Dvoretsky, V.G. Amino Acid Composition in Different Tissues of Iceland Scallop from the Barents Sea. Animals 2024, 14, 230. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, T.; Nakao, C.; Ohyama, H. Regulation of glycolysis during acclimation of scallops (Patinopecten yessoensis Jay) to anaerobiosis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 127, 45–52. [Google Scholar] [CrossRef] [PubMed]
- De Zwaan, A.; De Bont, A.M.T.; Zurburg, W.; Bayne, B.L.; Livingstone, D.R. On the role of strombine formation in the energy metabolism of the adductor muscle of a sessile bivalve. J. Comp. Physiol. A 1983, 149, 557–563. [Google Scholar] [CrossRef]
- Hayashi, H.J. Pyridoxal enzymes: Mechanistic diversity and uniformity. J. Biochem. 1995, 118, 463–473. [Google Scholar] [CrossRef]
- Toney, M.D. Reaction specificity in pyridoxal phosphate enzymes. Arch. Biochem. Biophys. 2005, 433, 279–287. [Google Scholar] [CrossRef]
- Boutet, I.; Meistertzheim, A.L.; Tanguy, A.; Thébault, M.T.; Moraga, D. Molecular characterization and expression of the gene encoding aspartate aminotransferase from the Pacific oyster Crassostrea gigas exposed to environmental stressors. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2005, 140, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Chia, F.S.; Bucklandnicks, J.; Young, C.M. Locomotion of marine invertebrate larvae—A review. Can. J. Zool./Rev. Can. Zool. 1984, 62, 1205–1222. [Google Scholar] [CrossRef]
- Fauquier, D.A.; Mazet, J.A.; Gulland, F.M.; Spraker, T.R.; Christopher, M.M. Distribution of tissue enzymes in three species of pinnipeds. J. Zoo Wildl. Med. Off. Publ. Am. Assoc. Zoo Vet. 2008, 39, 1–5. [Google Scholar] [CrossRef]
- Lewis, C.T.; Seyer, J.M.; Carlson, G.M. Cysteine 288: An essential hyperreactive thiol of cytosolic phosphoenolpyruvate carboxykinase (GTP). J. Biol. Chem. 1989, 264, 27–33. [Google Scholar] [CrossRef]
- Nordlie, R.C.; Lardy, H.A. Mammalian liver phosphoenolpyruvate carboxykinase activities. J. Biol. Chem. 1963, 238, 2259–2263. [Google Scholar] [CrossRef]
- Kniazkina, M.; Dyachuk, V. Neurogenesis of the scallop Azumapecten farreri: From the first larval sensory neurons to the definitive nervous system of juveniles. Front. Zool. 2022, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Morton, B.R. The Evolution of Eyes in the Bivalvia: New Insights. Am. Malacol. Bull. 2008, 26, 35–45. [Google Scholar] [CrossRef]
- Zammit, V.A.; Newsholme, E.A. Properties of pyruvate kinase and phosphoenolpyruvate carboxykinase in relation to the direction and regulation of phosphoenolpyruvate metabolism in muscles of the frog and marine invertebrates. Biochem. J. 1978, 174, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Duran-Encinas, Y.; Tremblay, R.; Genard, B.; Rivera-Pérez, C.; Lora-Vilchis, M.C.; Kraffe, E.; Palacios, E. Modulation of thermal stress response by prostaglandins in gills of the blue mussel Mytilus edulis. Aquaculture 2024, 582, 740478. [Google Scholar] [CrossRef]
- Hopkins, A.E. Activity of the adductor muscle in oysters. Physiol. Zool. 1936, 9, 498–507. [Google Scholar] [CrossRef]
- Beale, E.G.; Harvey, B.J.; Forest, C. PCK1 and PCK2 as candidate diabetes and obesity genes. Cell Biochem. Biophys. 2007, 48, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Hanson, R.W. Thematic minireview series: A perspective on the biology of phosphoenolpyruvate carboxykinase 55 years after its discovery. J. Biol. Chem. 2009, 284, 27021–27023. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Guebre-Egziabher, F.; Zhao, X.; Feriod, C.; Dong, J.; Alves, T.C.; Ioja, S.; Pongratz, R.L.; Bhanot, S.; Roden, M.; et al. A role for mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) in the regulation of hepatic gluconeogenesis. J. Biol. Chem. 2014, 289, 7257–7263. [Google Scholar] [CrossRef]
- Bryant, C. Metazoan Life without Oxygen, 1st ed.; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J. Scallop genome reveals molecular adaptations to semi-sessile life and neurotoxins. Nat. Commun. 2017, 8, 1721. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Peng, C.; Yang, Z.; Liu, Z.; Wang, S.; Yu, H.; Cui, C.; Hu, Y.; Xing, Q.; Hu, J.; Huang, X.; et al. A Systematical Survey on the TRP Channels Provides New Insight into Its Functional Diversity in Zhikong Scallop. Int. J. Mol. Sci. 2021, 22, 11075. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | CfOpDH1 | CfOpDH2 | CfOpDH3 | CfOpDH4 | CfOpDH5 | CfPEPCK1 | CfPEPCK2 | CfAST1 | CfAST2 |
|---|---|---|---|---|---|---|---|---|---|
| Gene ID | scaffold48249.6 | scaffold62993.3 | scaffold14657.35 | scaffold59443.2 | scaffold14657.34 | scaffold64265.9 | scaffold47297.11 | scaffold47499.10 | scaffold5663.76 |
| Total length (bp) | 20,552 | 11,244 | 45,172 | 14,543 | 7132 | 25,025 | 12,618 | 15,963 | 11,043 |
| ORF length (bp) | 1314 | 1203 | 1203 | 1212 | 1200 | 1977 | 1905 | 1248 | 1281 |
| 5′ UTR length (bp) | --- | 692 | 352 | 345 | 33 | 129 | 354 | 127 | 29 |
| 3′ UTR length (bp) | --- | 118 | 1169 | 1820 | --- | --- | 1123 | 61 | 1487 |
| Amino acids length | 437 | 400 | 400 | 403 | 399 | 658 | 634 | 415 | 426 |
| Weight (kDa) | 48.49 | 43.44 | 44.06 | 48.17 | 38.91 | 73.47 | 71.65 | 46.53 | 47.18 |
| Theoretical pI | 5.99 | 5.25 | 5.75 | 6.19 | 5.22 | 7.56 | 8.20 | 6.61 | 8.55 |
| Number of exons | 2 | 2 | 3 | 2 | 2 | 10 | 13 | 9 | 11 |
| Number of introns | 1 | 1 | 2 | 1 | 1 | 9 | 12 | 8 | 10 |
| Number of alpha helixes | 23 | 20 | 23 | 26 | 21 | 12 | 11 | 13 | 8 |
| Number of beta strands | 20 | 23 | 34 | 22 | 28 | 50 | 53 | 32 | 31 |
| Number of coils | 32 | 25 | 34 | 34 | 30 | 70 | 70 | 42 | 38 |
| Number of turns | 31 | 24 | 38 | 32 | 33 | 66 | 78 | 42 | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, L.; Liu, Z.; Sui, M.; Yang, Z.; Wang, H.; Chen, X.; Xu, Y.; Niu, Z.; Liu, N.; Xing, Q.; et al. The Glucose–Succinate Pathway: A Crucial Anaerobic Metabolic Pathway in the Scallop Chlamys farreri Experiencing Heat Stress. Int. J. Mol. Sci. 2024, 25, 4741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094741
Bao L, Liu Z, Sui M, Yang Z, Wang H, Chen X, Xu Y, Niu Z, Liu N, Xing Q, et al. The Glucose–Succinate Pathway: A Crucial Anaerobic Metabolic Pathway in the Scallop Chlamys farreri Experiencing Heat Stress. International Journal of Molecular Sciences. 2024; 25(9):4741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094741
Chicago/Turabian StyleBao, Lijingjing, Zhi Liu, Mingyi Sui, Zujing Yang, Haoran Wang, Xiaofei Chen, Yue Xu, Zehua Niu, Na Liu, Qiang Xing, and et al. 2024. "The Glucose–Succinate Pathway: A Crucial Anaerobic Metabolic Pathway in the Scallop Chlamys farreri Experiencing Heat Stress" International Journal of Molecular Sciences 25, no. 9: 4741. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094741