
Berberine Improves Cancer-Derived Myocardial Impairment in Experimental Cachexia Models by Targeting High-Mobility Group Box-1

Abstract
:1. Introduction
2. Results
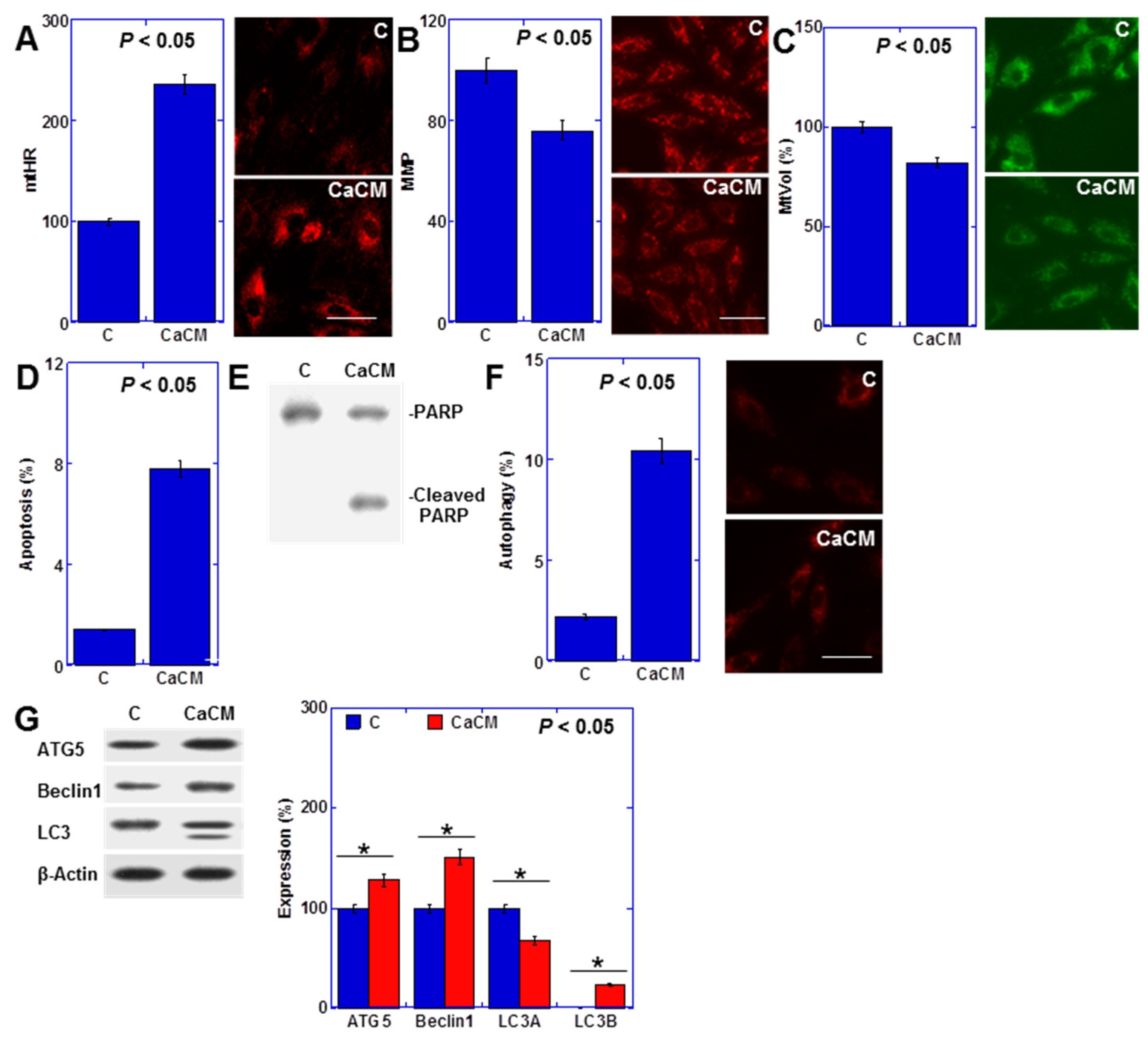
2.1. Impact of Rat Cancerous Ascites on Myocardial Cells
2.2. Inflammatory Cytokines in Cancerous Ascites
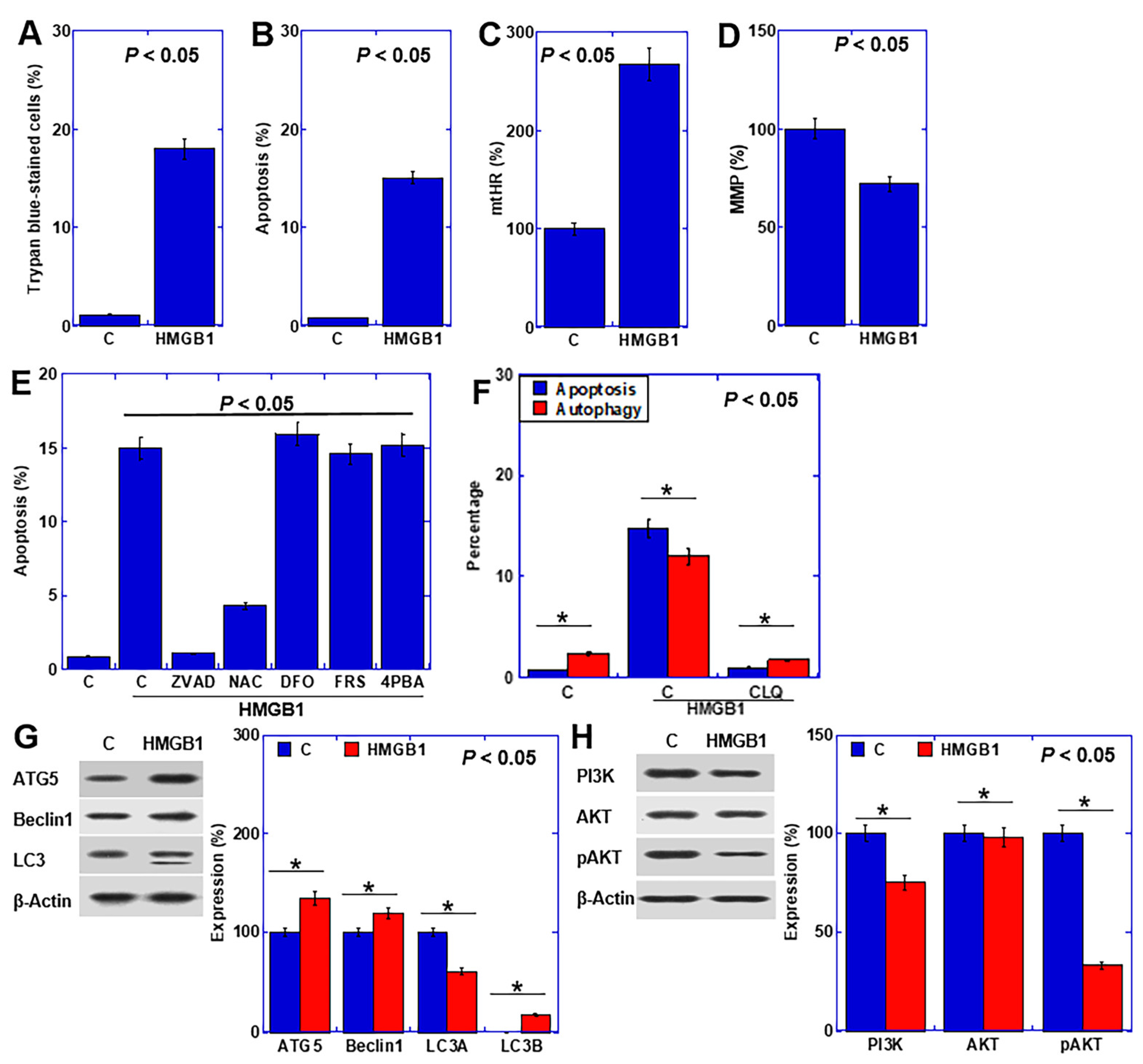
2.3. Effect of HMGB1 on Cardiomyocytes
2.4. HMGB1 Signal in Cardiomyocytes
2.5. Effect of BBR on CRC Cancer Cells
2.6. Effect of BBR on Cardiomyocytes
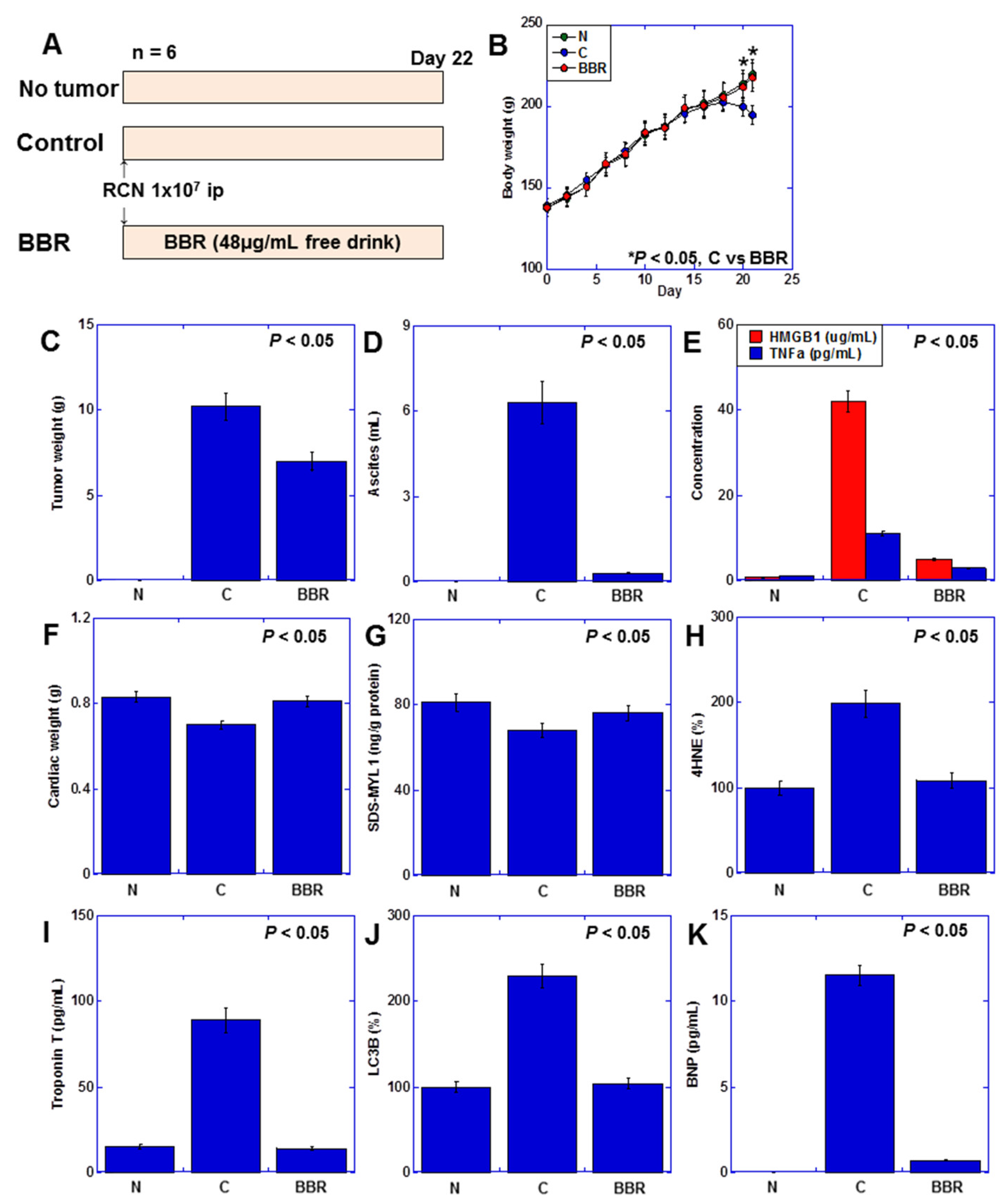
2.7. Effect of BBR on Myocardial Damage Using a Rat Cancer Cachexia Model
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. In Vitro Cachexia Model
4.3. Cell Growth, Cell Death and Apoptosis
4.4. Mitochondrial Imaging
4.5. Enzyme-Linked Immunosorbent Assay (ELISA) and Fluorometric Assay
4.6. Western Blot
4.7. Cell Death Inhibitor Assay
4.8. Reverse Transcription–Polymerase Chain Reaction
4.9. Small Interfering RNA
4.10. Detection of miRNA
4.11. Animals
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Blum, D.; Stene, G.B.; Solheim, T.S.; Fayers, P.; Hjermstad, M.J.; Baracos, V.E.; Fearon, K.; Strasser, F.; Kaasa, S. Validation of the Consensus-Definition for Cancer Cachexia and evaluation of a classification model—A study based on data from an international multicentre project (EPCRC-CSA). Ann. Oncol. 2014, 25, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Fearon, K.C.; Voss, A.C.; Hustead, D.S. Definition of cancer cachexia: Effect of weight loss, reduced food intake, and systemic inflammation on functional status and prognosis. Am. J. Clin. Nutr. 2006, 83, 1345–1350. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Mariean, C.R.; Tiucă, O.M.; Mariean, A.; Cotoi, O.S. Cancer Cachexia: New Insights and Future Directions. Cancers 2023, 15, 5590. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.M.; Sullivan, E.S. Impact of musculoskeletal degradation on cancer outcomes and strategies for management in clinical practice. Proc. Nutr. Soc. 2021, 80, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; Stemmler, B.; López-Soriano, F.J.; Busquets, S. Nonmuscle Tissues Contribution to Cancer Cachexia. Mediat. Inflamm. 2015, 2015, 182872. [Google Scholar] [CrossRef] [PubMed]
- Kazemi-Bajestani, S.M.; Becher, H.; Fassbender, K.; Chu, Q.; Baracos, V.E. Concurrent evolution of cancer cachexia and heart failure: Bilateral effects exist. J. Cachexia Sarcopenia Muscle 2014, 5, 95–104. [Google Scholar] [CrossRef]
- Florido, R.; Daya, N.R.; Ndumele, C.E.; Koton, S.; Russell, S.D.; Prizment, A.; Blumenthal, R.S.; Matsushita, K.; Mok, Y.; Felix, A.S.; et al. Cardiovascular Disease Risk Among Cancer Survivors: The Atherosclerosis Risk In Communities (ARIC) Study. J. Am. Coll. Cardiol. 2022, 80, 22–32. [Google Scholar] [CrossRef]
- Ausoni, S.; Calamelli, S.; Saccà, S.; Azzarello, G. How progressive cancer endangers the heart: An intriguing and underestimated problem. Cancer Metastasis Rev. 2020, 39, 535–552. [Google Scholar] [CrossRef]
- Mamidanna, R.; Nachiappan, S.; Bottle, A.; Aylin, P.; Faiz, O. Defining the timing and causes of death amongst patients undergoing colorectal resection in England. Color. Dis. 2016, 18, 586–593. [Google Scholar] [CrossRef]
- Herrmann, J. Adverse cardiac effects of cancer therapies: Cardiotoxicity and arrhythmia. Nat. Rev. Cardiol. 2020, 17, 474–502. [Google Scholar] [CrossRef] [PubMed]
- Perez, I.E.; Taveras Alam, S.; Hernandez, G.A.; Sancassani, R. Cancer Therapy-Related Cardiac Dysfunction: An Overview for the Clinician. Clin. Med. Insights Cardiol. 2019, 13, 1179546819866445. [Google Scholar] [CrossRef] [PubMed]
- Kamphuis, J.A.M.; Linschoten, M.; Cramer, M.J.; Gort, E.H.; van Rhenen, A.; Asselbergs, F.W.; Doevendans, P.A.; Teske, A.J. Cancer Therapy-Related Cardiac Dysfunction of Nonanthracycline Chemotherapeutics: What Is the Evidence? JACC Cardio Oncol. 2019, 1, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.T. The pathogenesis and treatment of cardiac atrophy in cancer cachexia. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H466–H477. [Google Scholar] [CrossRef] [PubMed]
- Anker, M.S.; Sanz, A.P.; Zamorano, J.L.; Mehra, M.R.; Butler, J.; Riess, H.; Coats, A.J.S.; Anker, S.D. Advanced cancer is also a heart failure syndrome: A hypothesis. Eur. J. Heart Fail. 2021, 23, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, Y.; Nukaga, S.; Mori, T.; Fujiwara-Tani, R.; Fujii, K.; Mori, S.; Goto, K.; Kishi, S.; Sasaki, T.; Nakashima, C.; et al. Evaluation of cancer-derived myocardial impairments using a mouse model. Oncotarget 2020, 11, 3712–3722. [Google Scholar] [CrossRef]
- Nukaga, S.; Mori, T.; Miyagawa, Y.; Fujiwara-Tani, R.; Sasaki, T.; Fujii, K.; Mori, S.; Goto, K.; Kishi, S.; Nakashima, C.; et al. Combined administration of lauric acid and glucose improved cancer-derived cardiac atrophy in a mouse cachexia model. Cancer Sci. 2020, 111, 4605–4615. [Google Scholar] [CrossRef]
- Ohmori, H.; Fujiwara-Tani, R.; Nukaga, S.; Nishida, R.; Fujii, K.; Mori, S.; Ogata, R.; Ikemoto, A.; Sasaki, R.; Kishi, S.; et al. Investigation of cancer-induced myocardial damage in autopsy cases-A comparison of cases with and without chemotherapy. Pathol. Int. 2024, 74, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, H.; Luo, Y.; Kuniyasu, H. Non-histone nuclear factor HMGB1 as a therapeutic target in colorectal cancer. Expert. Opin. Ther. Targets 2011, 15, 183–193. [Google Scholar] [CrossRef]
- Luo, Y.; Yoneda, J.; Ohmori, H.; Sasaki, T.; Shimbo, K.; Eto, S.; Kato, Y.; Miyano, H.; Kobayashi, T.; Sasahira, T.; et al. Cancer usurps skeletal muscle as an energy repository. Cancer Res. 2014, 74, 330–340. [Google Scholar] [CrossRef]
- Ohmori, H.; Kawahara, I.; Mori, T.; Nukaga, S.; Luo, Y.; Kishi, S.; Fujiwara-Tani, R.; Mori, S.; Goto, K.; Sasaki, T.; et al. Evaluation of Parameters for Cancer-Induced Sarcopenia in Patients Autopsied after Death from Colorectal Cancer. Pathobiology 2019, 86, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Jin, Y.; Gao, Z.; Zeng, Y.; Du, J.; Yan, H.; Chen, X.; Ping, L.; Lin, N.; Yang, B.; et al. Autophagic degradation of CCN2 (cellular communication network factor 2) causes cardiotoxicity of sunitinib. Autophagy 2022, 18, 1152–1173. [Google Scholar] [CrossRef] [PubMed]
- Raucci, A.; Di Maggio, S.; Scavello, F.; D’Ambrosio, A.; Bianchi, M.E.; Capogrossi, M.C. The Janus face of HMGB1 in heart disease: A necessary update. Cell Mol. Life Sci. 2019, 76, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J Clin 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Wakao, F. CANCER STATISTICS IN JAPAN 2021; Foundation for Promotion of Cancer Research: Tokyo, Japan, 2021; p. 146. [Google Scholar]
- Kasprzak, A. The Role of Tumor Microenvironment Cells in Colorectal Cancer (CRC) Cachexia. Int. J. Mol. Sci. 2021, 22, 1565. [Google Scholar] [CrossRef] [PubMed]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. 2019, 33, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Hu, M.; Huang, Z.; Fang, K.; Wang, D.; Chen, Q.; Li, J.; Yang, D.; Zou, X.; Xu, L.; et al. Berberine Attenuates Intestinal Mucosal Barrier Dysfunction in Type 2 Diabetic Rats. Front. Pharmacol. 2017, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, Y.; Huang, W.; Selwyn, F.P.; Klaassen, C.D. Dose-response effect of berberine on bile acid profile and gut microbiota in mice. BMC Complement. Altern. Med. 2016, 16, 394. [Google Scholar] [CrossRef]
- Mori, S.; Fujiwara-Tani, R.; Gyoten, M.; Nukaga, S.; Sasaki, R.; Ikemoto, A.; Ogata, R.; Kishi, S.; Fujii, K.; Kuniyasu, H. Berberine Induces Combined Cell Death in Gastrointestinal Cell Lines. Int. J. Mol. Sci. 2023, 24, 6588. [Google Scholar] [CrossRef]
- Zhu, J.R.; Lu, H.D.; Guo, C.; Fang, W.R.; Zhao, H.D.; Zhou, J.S.; Wang, F.; Zhao, Y.L.; Li, Y.M.; Zhang, Y.D.; et al. Berberine attenuates ischemia-reperfusion injury through inhibiting HMGB1 release and NF-κB nuclear translocation. Acta Pharmacol. Sin. 2018, 39, 1706–1715. [Google Scholar] [CrossRef]
- Shi, J.; Xu, H.; Cavagnaro, M.J.; Li, X.; Fang, J. Blocking HMGB1/RAGE Signaling by Berberine Alleviates A1 Astrocyte and Attenuates Sepsis-Associated Encephalopathy. Front. Pharmacol. 2021, 12, 760186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Peng, L.; Zhang, J.; Dong, Y.P.; Wang, C.J.; Liu, C.; Xia, D.Y.; Zhang, X.S. Berberine Ameliorates Subarachnoid Hemorrhage Injury via Induction of Sirtuin 1 and Inhibiting HMGB1/Nf-κB Pathway. Front. Pharmacol. 2020, 11, 1073. [Google Scholar] [CrossRef] [PubMed]
- Lucia, S.; Esposito, M.; Rossi Fanelli, F.; Muscaritoli, M. Cancer cachexia: From molecular mechanisms to patient’s care. Crit. Rev. Oncog. 2012, 17, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Belloum, Y.; Rannou-Bekono, F.; Favier, F.B. Cancer-induced cardiac cachexia: Pathogenesis and impact of physical activity (Review). Oncol. Rep. 2017, 37, 2543–2552. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.W.; Liu, M.M.; Duan, R.; Tao, Y.F.; Zhou, J.S.; Fang, W.R.; Zhu, J.R.; Niu, L.; Sun, J.G. The lncRNA Malat1 functions as a ceRNA to contribute to berberine-mediated inhibition of HMGB1 by sponging miR-181c-5p in poststroke inflammation. Acta Pharmacol. Sin. 2020, 41, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Long, T.; Pan, W.; Li, F.; Sheikh, S.A.; Xie, Q.; Zhang, C. Berberine up-regulates miR-340-5p to protect myocardial ischaemia/reperfusion from HMGB1-mediated inflammatory injury. ESC Heart Fail. 2023, 10, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Livesey, K.M.; Cheh, C.W.; Farkas, A.; Loughran, P.; Hoppe, G.; Bianchi, M.E.; Tracey, K.J.; Zeh, H.J.; et al. Endogenous HMGB1 regulates autophagy. J. Cell Biol. 2010, 190, 881–892. [Google Scholar] [CrossRef]
- Xie, G.; Jin, H.; Mikhail, H.; Pavel, V.; Yang, G.; Ji, B.; Lu, B.; Li, Y. Autophagy in sarcopenia: Possible mechanisms and novel therapies. Biomed. Pharmacother. 2023, 165, 115147. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Gao, M.; Liu, B.; Qin, Y.; Chen, L.; Liu, H.; Wu, H.; Gong, G. Mitochondrial autophagy: Molecular mechanisms and implications for cardiovascular disease. Cell Death Dis. 2022, 13, 444. [Google Scholar] [CrossRef]
- Sun, Y.; Yao, X.; Zhang, Q.J.; Zhu, M.; Liu, Z.P.; Ci, B.; Xie, Y.; Carlson, D.; Rothermel, B.A.; Sun, Y.; et al. Beclin-1-Dependent Autophagy Protects the Heart During Sepsis. Circulation 2018, 138, 2247–2262. [Google Scholar] [CrossRef]
- Yamaguchi, O. Autophagy in the Heart. Circ. J. 2019, 83, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Kalfalah, F.; Sobek, S.; Bornholz, B.; Götz-Rösch, C.; Tigges, J.; Fritsche, E.; Krutmann, J.; Köhrer, K.; Deenen, R.; Ohse, S.; et al. Inadequate mito-biogenesis in primary dermal fibroblasts from old humans is associated with impairment of PGC1A-independent stimulation. Exp. Gerontol. 2014, 56, 59–68. [Google Scholar] [CrossRef]
- Chen, R.; Zou, J.; Zhong, X.; Li, J.; Kang, R.; Tang, D. HMGB1 in the interplay between autophagy and apoptosis in cancer. Cancer Lett. 2024, 581, 216494. [Google Scholar] [CrossRef]
- Livesey, K.M.; Kang, R.; Vernon, P.; Buchser, W.; Loughran, P.; Watkins, S.C.; Zhang, L.; Manfredi, J.J.; Zeh, H.J., III; Li, L.; et al. p53/HMGB1 complexes regulate autophagy and apoptosis. Cancer Res. 2012, 72, 1996–2005. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.S.; Huang, Y.; Qu, J.F.; Wang, Y.J.; Huang, Z.Y.; Wang, F.Y.; Yi, W.J.; Liu, X.X. Panaxynol ameliorates cardiac ischemia/reperfusion injury by suppressing NLRP3-induced pyroptosis and apoptosis via HMGB1/TLR4/NF-κB axis. Int. Immunopharmacol. 2023, 121, 110222. [Google Scholar] [CrossRef]
- Wang, W.K.; Lu, Q.H.; Zhang, J.N.; Wang, B.; Liu, X.J.; An, F.S.; Qin, W.D.; Chen, X.Y.; Dong, W.Q.; Zhang, C.; et al. HMGB1 mediates hyperglycaemia-induced cardiomyocyte apoptosis via ERK/Ets-1 signalling pathway. J. Cell Mol. Med. 2014, 18, 2311–2320. [Google Scholar] [CrossRef] [PubMed]
- He, D.W.; Liu, D.Z.; Luo, X.Z.; Chen, C.B.; Lu, C.H.; Na, N.; Huang, F. HMGB1-RAGE axis contributes to myocardial ischemia/reperfusion injury via regulation of cardiomyocyte autophagy and apoptosis in diabetic mice. Biol. Chem. 2024, 405, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. The Anti-Cancer Mechanisms of Berberine: A Review. Cancer Manag. Res. 2020, 12, 695–702. [Google Scholar] [CrossRef]
- Huang, C.; Liu, H.; Gong, X.L.; Wu, L.Y.; Wen, B. Effect of evodiamine and berberine on the interaction between DNMTs and target microRNAs during malignant transformation of the colon by TGF-β1. Oncol. Rep. 2017, 37, 1637–1645. [Google Scholar] [CrossRef]
- Li, X.; Su, Y.; Li, N.; Zhang, F.R.; Zhang, N. Berberine Attenuates MPP+-Induced Neuronal Injury by Regulating LINC00943/miR-142-5p/KPNA4/NF-κB Pathway in SK-N-SH Cells. Neurochem. Res. 2021, 46, 3286–3300. [Google Scholar] [CrossRef]
- Andersson, U.; Yang, H. HMGB1 is a critical molecule in the pathogenesis of Gram-negative sepsis. J. Intensive Med. 2022, 2, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Huang, K.H.; Yeh, T.S.; Chi, C.W. Somatic alterations in mitochondrial DNA and mitochondrial dysfunction in gastric cancer progression. World J. Gastroenterol. 2014, 20, 3950–3959. [Google Scholar] [CrossRef] [PubMed]
- Javed Iqbal, M.; Quispe, C.; Javed, Z.; Sadia, H.; Qadri, Q.R.; Raza, S.; Salehi, B.; Cruz-Martins, N.; Abdulwanis Mohamed, Z.; Sani Jaafaru, M.; et al. Nanotechnology-Based Strategies for Berberine Delivery System in Cancer Treatment: Pulling Strings to Keep Berberine in Power. Front. Mol. Biosci. 2020, 7, 624494. [Google Scholar] [CrossRef] [PubMed]
- Vahedi-Mazdabadi, Y.; Shahinfar, H.; Toushih, M.; Shidfar, F. Effects of berberine and barberry on selected inflammatory biomarkers in adults: A systematic review and dose-response meta-analysis of randomized clinical trials. Phytother. Res. 2023, 37, 5541–5557. [Google Scholar] [CrossRef]
- Tsai, P.L.; Tsai, T.H. Hepatobiliary excretion of berberine. Drug Metab. Dispos. 2004, 32, 405–412. [Google Scholar] [CrossRef]
- Choksey, A.; Carter, R.D.; Thackray, B.D.; Ball, V.; Kennedy, B.W.C.; Ha, L.H.T.; Sharma, E.; Broxholme, J.; Castro-Guarda, M.; Murphy, M.P.; et al. AICAR confers prophylactic cardioprotection in doxorubicin-induced heart failure in rats. J. Mol. Cell Cardiol. 2024. online ahead of print. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Primer | |||
|---|---|---|---|
| Gene Symbol | Gene Bank ID | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
| miR-181c-5p | NR_031897.1 | agaacttgccaagggtttg | gcagttccaggcctcggg |
| miR-340-5p | NR_106669.1 | cacttgtactcggtgtga | taagataccaggtatggc |
| U6 | K00784.1 | gtgcctgct tcggcagca | cgcttcacgaatttgcgtg |
| rat HMGB1 | NM_012963.3 | cacaagaagaagcacccgga | catcctcctcgtcgtcttcc |
| rat ACTB | NM_031144.3 | tcaacaccccagccatgtac | aatgcctgggtacatggtgg |
| Antibody | |||
| Target | Clone or Cat# | Company | |
| ATG5 | EPR1755(2) | Abcam, Waltham, MA, USA | |
| Beclin1 | Clone 669922 | R&D Systems, Minneapolis, MN, USA | |
| LC3 | ab51520 | Abcam, Waltham, MA, USA | |
| β-actin | ab8227 | Abcam, Waltham, MA, USA | |
| PARP | GTX100573 | GeneTex, Irvine, CA, USA | |
| HMGB1 | 3E8 | Biolegend, San Diego, CA, USA | |
| rat TNFα | 52B83 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | |
| rat IL6 | 1.2-2B11-2G10 | Abcam, Waltham, MA, USA | |
| PI3K | #4292 | Cell Signaling, Danvers, MA, USA | |
| AKT | ab8805 | Abcam, Waltham, MA, USA | |
| pAKT | ab38449 | Abcam, Waltham, MA, USA | |
| RAGE | EPR21171 | Abcam, Waltham, MA, USA | |
| TLR4 | 76B357.1 | Abcam, Waltham, MA, USA | |
| IκBα | 66418-1-Ig | Proteintech, Rosemont, IL, USA | |
| pIκBα | 14D4 | Cell Signaling, Danvers, MA, USA | |
| NFκBp65 | D14E12 | Cell Signaling, Danvers, MA, USA | |
| Lamin | ab65986 | Abcam, Waltham, MA, USA | |
| BCL2 | 26593-1-AP | Proteintech, Rosemont, IL, USA | |
| BAX | E63 | Abcam, Waltham, MA, USA | |
| ELISA | |||
| Target | Catalog# | Company | |
| HMGB1 | LS-F4039 | Shino-Test, Sagamihara, Japan | |
| rat TNFα | ab100785 | Abcam, Waltham, MA, USA | |
| rat IL6 | ab234570 | Abcam, Waltham, MA, USA | |
| MYL1 | orb1211541 | Biorbyt, Cambridge, UK | |
| 4HNE | ab238538 | Abcam, Waltham, MA, USA | |
| Troponin T | ABIN6970886 | antibodies-online.com, Limerick, PA, USA | |
| LC3B | #35172 | Cell Signaling, Danvers, MA, USA | |
| rat BNP | ab108816 | Abcam, Waltham, MA, USA | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, K.; Fujiwara-Tani, R.; Nukaga, S.; Miyagawa, Y.; Kawahara, I.; Nishida, R.; Ikemoto, A.; Sasaki, R.; Ogata, R.; Kishi, S.; et al. Berberine Improves Cancer-Derived Myocardial Impairment in Experimental Cachexia Models by Targeting High-Mobility Group Box-1. Int. J. Mol. Sci. 2024, 25, 4735. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094735
Goto K, Fujiwara-Tani R, Nukaga S, Miyagawa Y, Kawahara I, Nishida R, Ikemoto A, Sasaki R, Ogata R, Kishi S, et al. Berberine Improves Cancer-Derived Myocardial Impairment in Experimental Cachexia Models by Targeting High-Mobility Group Box-1. International Journal of Molecular Sciences. 2024; 25(9):4735. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094735
Chicago/Turabian StyleGoto, Kei, Rina Fujiwara-Tani, Shota Nukaga, Yoshihiro Miyagawa, Isao Kawahara, Ryoichi Nishida, Ayaka Ikemoto, Rika Sasaki, Ruiko Ogata, Shingo Kishi, and et al. 2024. "Berberine Improves Cancer-Derived Myocardial Impairment in Experimental Cachexia Models by Targeting High-Mobility Group Box-1" International Journal of Molecular Sciences 25, no. 9: 4735. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094735