
Cinnamaldehyde Protects against P. gingivalis Induced Intestinal Epithelial Barrier Dysfunction in IEC-6 Cells via the PI3K/Akt-Mediated NO/Nrf2 Signaling Pathway
 , ,
, ,
Abstract
:1. Introduction
2. Results
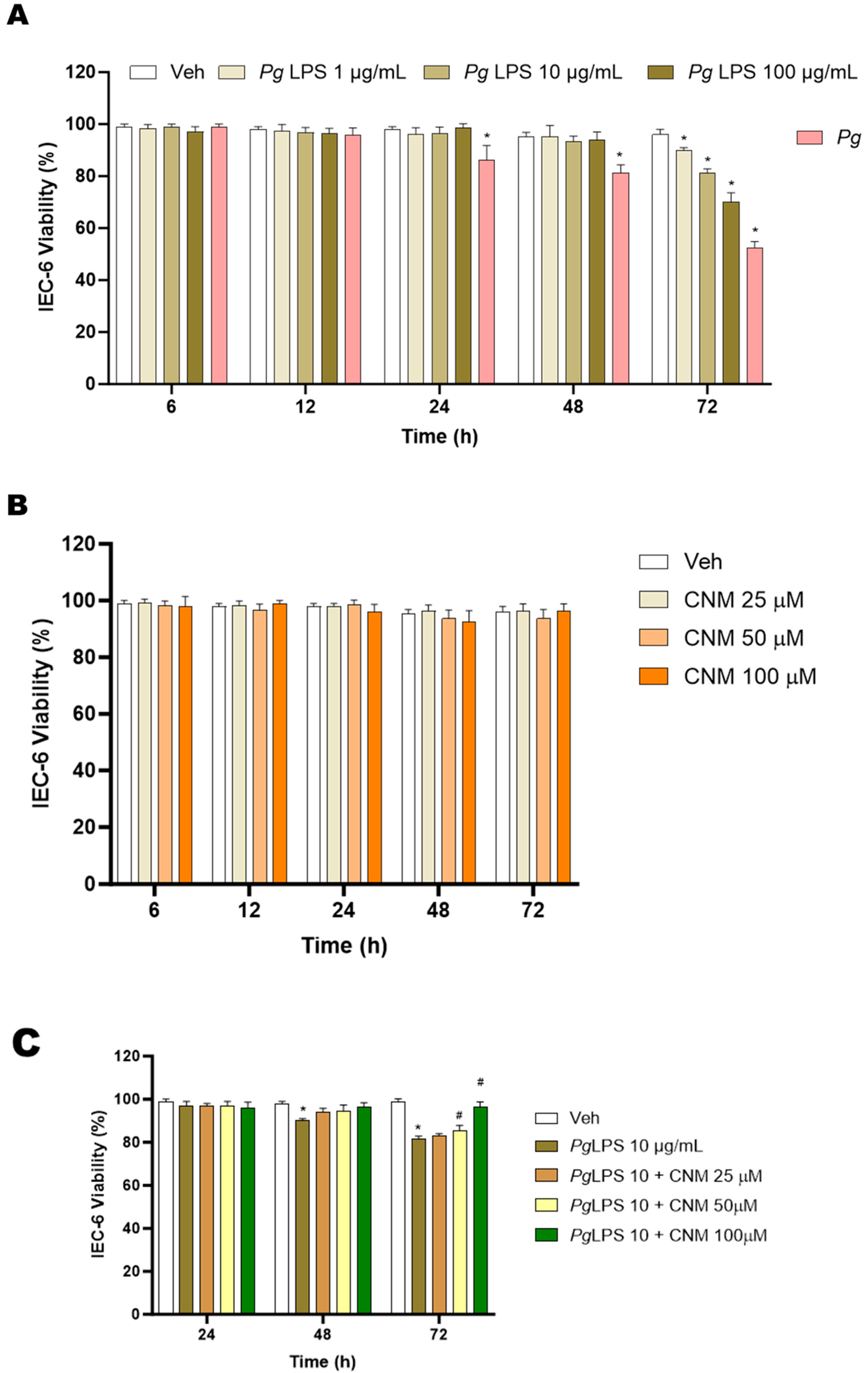
2.1. Protective Effects of Cinnamaldehyde on Pg, Pg-LPS-Induced Cytotoxicity in IEC-6 Cells
2.2. Cinnamaldehyde Inhibited Pg or Pg-LPS-Induced Oxidative Stress in IEC-6 Cells
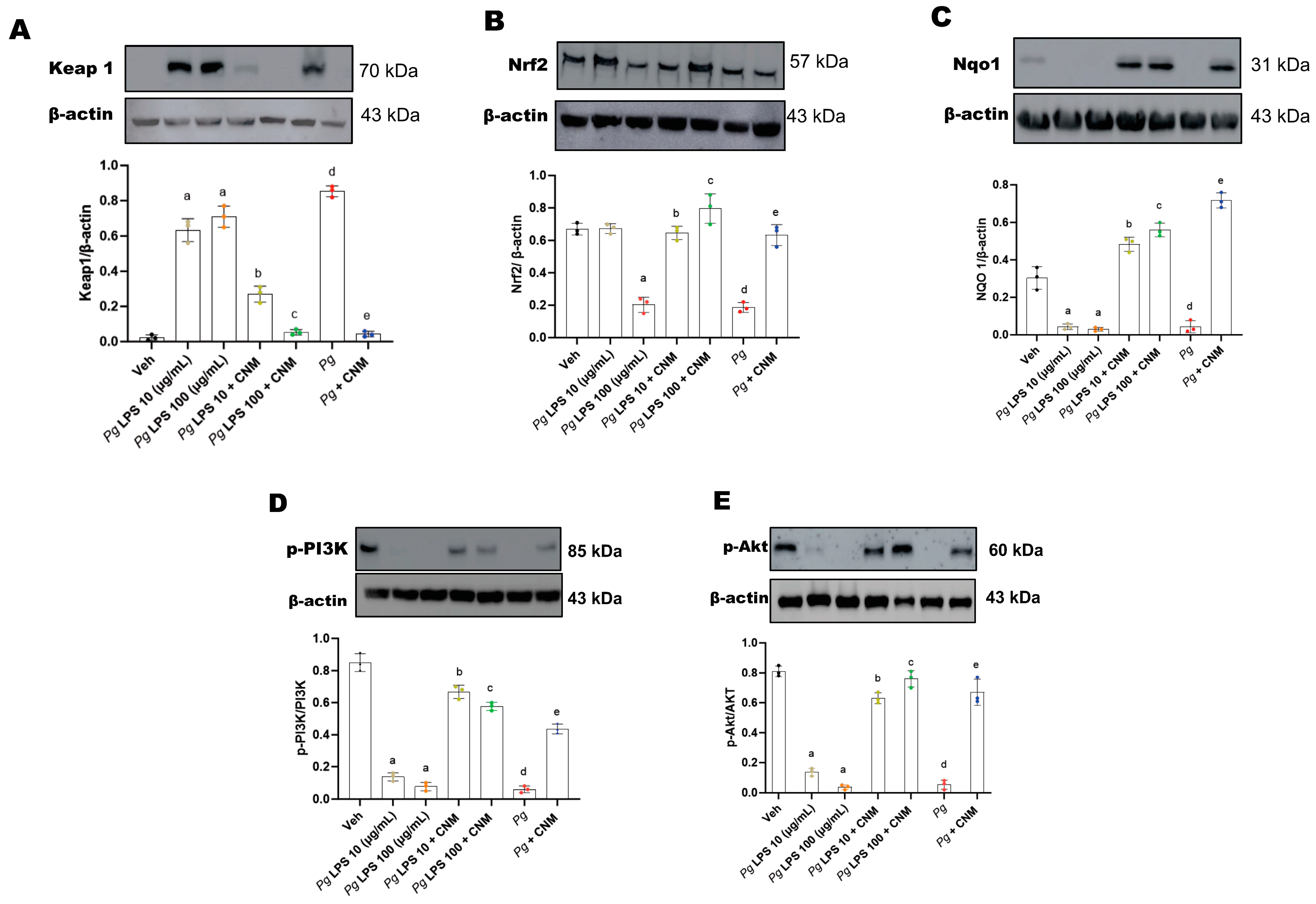
2.3. Cinnamaldehyde Activates Nrf2 Translocation via PI3/Akt Pathway
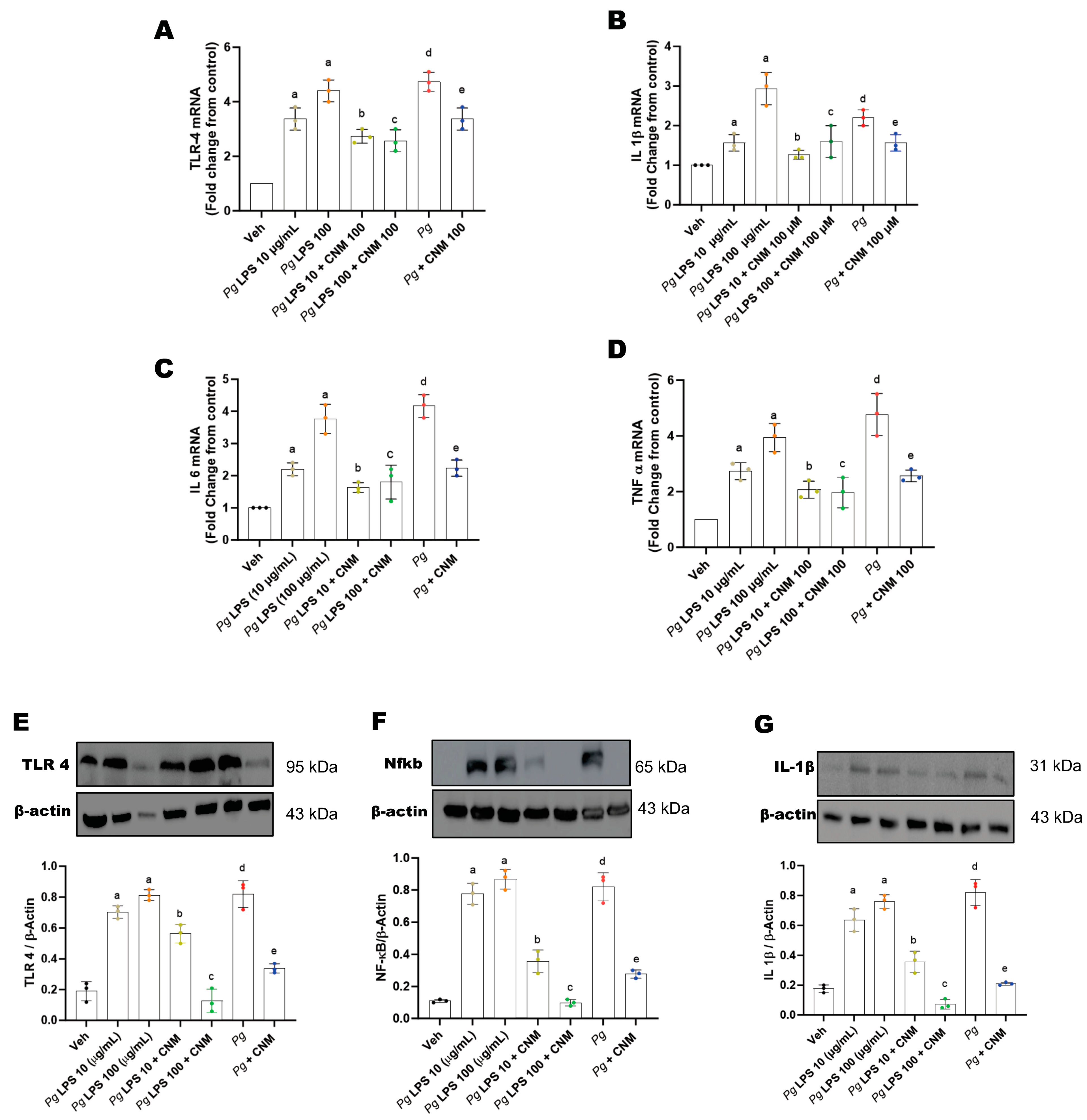
2.4. Cinnamaldehyde Inhibits the Expression of Inducible Nitric Oxide Synthase (iNOS)
2.5. Cinnamaldehyde Reduces the Expression Levels of Proinflammatory Cytokines
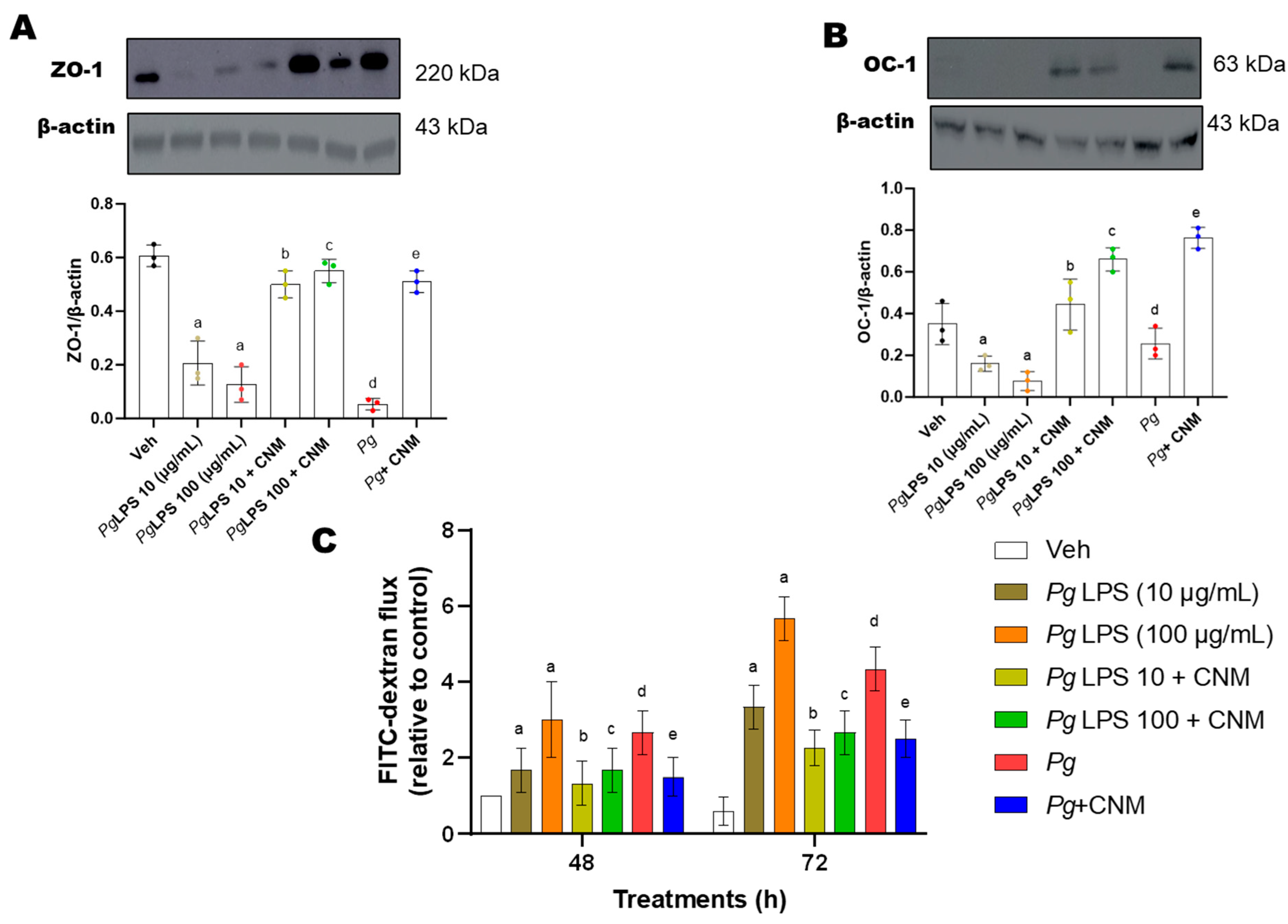
2.6. Cinnamaldehyde Increased Occludin and ZO-1 Protein Expression and Intestinal Epithelial Barrier Function in IEC-6 Cells
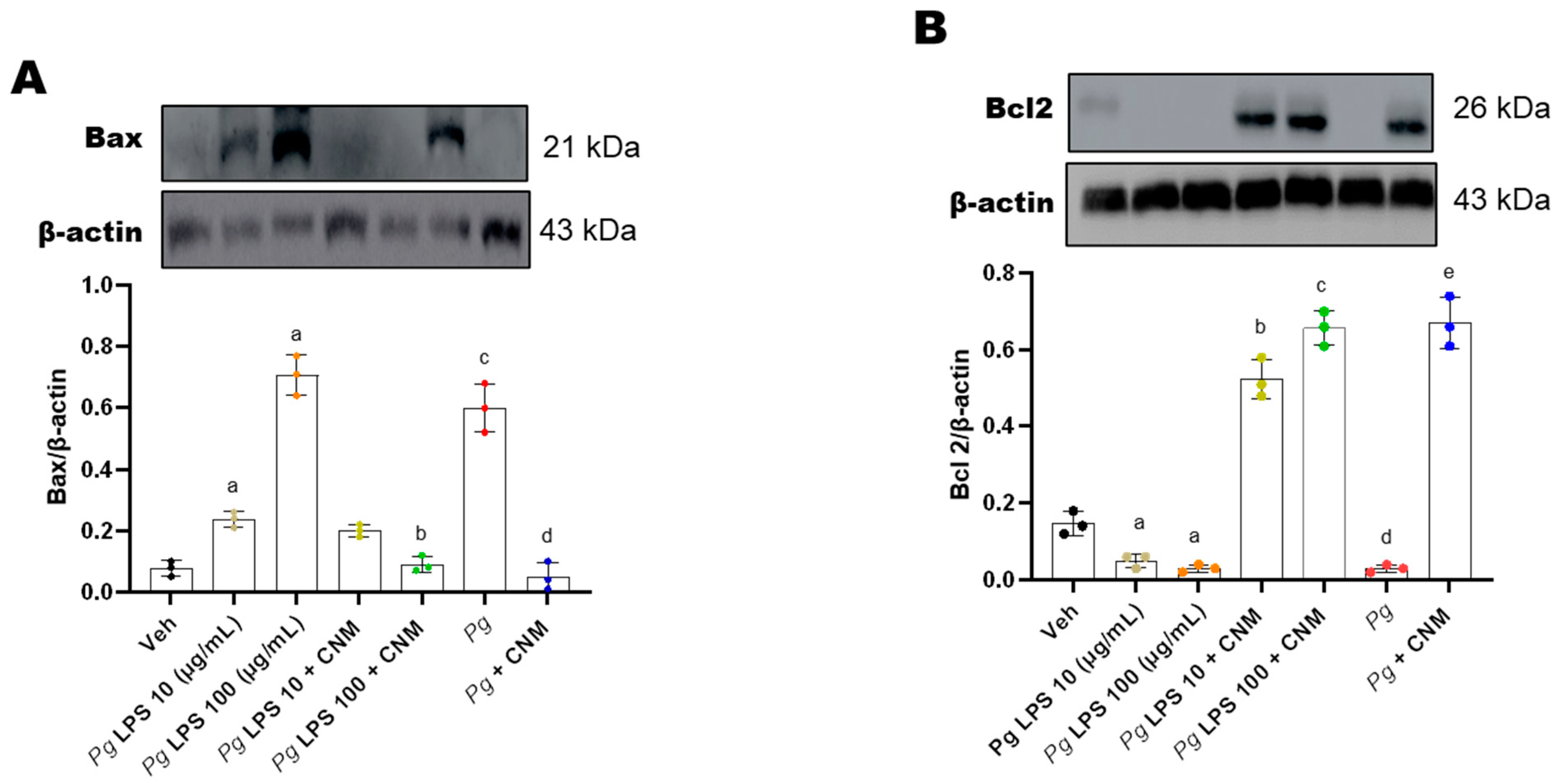
2.7. Cinnamaldehyde Inhibits Bax/Caspase-3 Signaling Pathway in IEC-6 Cells Infected with Pg-LPS/P. gingivalis
3. Discussion
4. Materials and Methods
4.1. Pre-Culture of Pg
4.2. IEC-6 Cell Culture
4.3. Human Oxygen-Bacteria Anaerobic (HoxBan) Coculture and Treatment
4.4. Cell Viability Assay
4.5. Cinnamaldehyde Treatment
4.6. Intracellular ROS Determination
4.7. Nitric Oxide and Malondialdehyde Determination
4.8. Intestinal Epithelial Barrier Function Determination
4.9. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction Analysis
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Srivastava, R.; Srivastava, M.; Verma, P.; Gautam, A. Metabolic syndrome and periodontal disease: An overview for physicians. J. Fam. Med. Prim. Care 2019, 8, 3492. [Google Scholar] [CrossRef] [PubMed]
- Lira-Junior, R.; Boström, E.A. Oral-gut connection: One step closer to an integrated view of the gastrointestinal tract? Mucosal Immunol. 2018, 11, 316–318. [Google Scholar] [CrossRef]
- Damgaard, C.; Danielsen, A.K.; Enevold, C.; Massarenti, L.; Nielsen, C.H.; Holmstrup, P.; Belstrøm, D. Porphyromonas gingivalis in saliva associates with chronic and aggressive periodontitis. J. Oral Microbiol. 2019, 11, 1653123. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Huang, W.; Pan, Z.; Cui, H.; Qi, G.; Zhou, X.; Chen, H. Quantitative analysis of microbiota in saliva, supragingival, and subgingival plaque of Chinese adults with chronic periodontitis. Clin. Oral Investig. 2012, 16, 1579–1588. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, W.; Wang, J.; Ma, J.; Zhang, M.; Lu, X.; Liu, J.; Kou, Y. Multifaceted Impacts of Periodontal Pathogens in Disorders of the Intestinal Barrier. Front. Immunol. 2021, 12, 693479. [Google Scholar] [CrossRef]
- Buckley, A.; Turner, J.R. Cell Biology of Tight Junction Barrier Regulation and Mucosal Disease. Cold Spring Harb. Perspect. Biol. 2018, 10, a029314. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Buckley, A.G.; Looi, K.; Iosifidis, T.; Ling, K.-M.; Sutanto, E.N.; Martinovich, K.M.; Kicic-Starcevich, E.; Garratt, L.W.; Shaw, N.C.; Lannigan, F.J.; et al. Visualisation of Multiple Tight Junctional Complexes in Human Airway Epithelial Cells. Biol. Proced. Online 2018, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.; Kim, H.J. Intestinal barrier dysfunction orchestrates the onset of inflammatory host–microbiome cross-talk in a human gut inflammation-on-a-chip. Proc. Natl. Acad. Sci. USA 2018, 115, E10539–E10547. [Google Scholar] [CrossRef]
- Wen, Z.; Liu, W.; Li, X.; Chen, W.; Liu, Z.; Wen, J.; Liu, Z. A Protective Role of the NRF2-Keap1 Pathway in Maintaining Intestinal Barrier Function. Oxidative Med. Cell. Longev. 2019, 2019, 1759149. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Goteri, G.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D. The Role of NQO1 in Ovarian Cancer. Int. J. Mol. Sci. 2023, 24, 7839. [Google Scholar] [CrossRef] [PubMed]
- Chumpitazi, B.P.; Kearns, G.L.; Shulman, R.J. Review article: The physiological effects and safety of peppermint oil and its efficacy in irritable bowel syndrome and other functional disorders. Aliment. Pharmacol. Ther. 2018, 47, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Sprouse, J.C.; Freeman, M.L.; Gangula, P.R. Activation of Nrf2 attenuates delayed gastric emptying in obesity induced diabetic (T2DM) female mice. Free Radic. Biol. Med. 2019, 135, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Wilus, D.; Tabatabai, M.; Freeman, M.L.; Gangula, P.R. Mechanistic role of antioxidants in rescuing delayed gastric emptying in high fat diet induced diabetic female mice. Biomed. Pharmacother. 2021, 137, 111370. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Raju, A.V.; Freeman, M.L.; Srinivasan, S.; Gangula, P.R. Nrf2 attenuates hyperglycemia-induced nNOS impairment in adult mouse primary enteric neuronal crest cells and normalizes stomach function. Am. J. Physiol. Liver Physiol. 2022, 322, G368–G382. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Mao, H.; Lu, X.; Shi, T.; Wang, J. Cinnamaldehyde Promotes the Intestinal Barrier Functions and Reshapes Gut Microbiome in Early Weaned Rats. Front. Nutr. 2021, 8, 748503. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, S.; Yin, P.; Li, D.; Mei, C.; Yu, X.; Shi, Y.; Jiang, L.; Liu, F. Punicalagin induces Nrf2 translocation and HO-1 expression via PI3K/Akt, protecting rat intestinal epithelial cells from oxidative stress. Int. J. Hyperth. 2016, 32, 465–473. [Google Scholar] [CrossRef]
- Crabtree, M.J.; Tatham, A.L.; Hale, A.B.; Alp, N.J.; Channon, K.M. Critical Role for Tetrahydrobiopterin Recycling by Dihydrofolate Reductase in Regulation of Endothelial Nitric-oxide Synthase Coupling. J. Biol. Chem. 2009, 284, 28128–28136. [Google Scholar] [CrossRef]
- Andrews, C.; McLean, M.H.; Durum, S.K. Cytokine Tuning of Intestinal Epithelial Function. Front. Immunol. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Hatok, J.; Racay, P. Bcl-2 family proteins: Master regulators of cell survival. Biomol. Concepts 2016, 7, 259–270. [Google Scholar] [CrossRef]
- Bao, J.; Li, L.; Zhang, Y.; Wang, M.; Chen, F.; Ge, S.; Chen, B.; Yan, F. Periodontitis may induce gut microbiota dysbiosis via salivary microbiota. Int. J. Oral Sci. 2022, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, Y.; Ouhara, K.; Munenaga, S.; Shoji, M.; Ozawa, T.; Hisatsune, J.; Kado, I.; Kajiya, M.; Matsuda, S.; Kawai, T.; et al. Effect of Porphyromonas gingivalis infection on gut dysbiosis and resultant arthritis exacerbation in mouse model. Arthritis Res. Ther. 2020, 22, 249. [Google Scholar] [CrossRef] [PubMed]
- Vona, R.; Pallotta, L.; Cappelletti, M.; Severi, C.; Matarrese, P. The Impact of Oxidative Stress in Human Pathology: Focus on Gastrointestinal Disorders. Antioxidants 2021, 10, 201. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.; Zhang, X.; Lu, Y.; Chen, H. New insights in intestinal oxidative stress damage and the health intervention effects of nutrients: A review. J. Funct. Foods 2020, 75, 104248. [Google Scholar] [CrossRef]
- Charoensaensuk, V.; Chen, Y.-C.; Lin, Y.-H.; Ou, K.-L.; Yang, L.-Y.; Lu, D.-Y. Porphyromonas gingivalis Induces Proinflammatory Cytokine Expression Leading to Apoptotic Death through the Oxidative Stress/NF-κB Pathway in Brain Endothelial Cells. Cells 2021, 10, 3033. [Google Scholar] [CrossRef] [PubMed]
- Cherian, D.; Peter, T.; Narayanan, A.; Madhavan, S.S.; Achammada, S.; Vynat, G.P. Malondialdehyde as a marker of oxidative stress in periodontitis patients. J. Pharm. Bioallied Sci. 2019, 11, 297. [Google Scholar] [CrossRef] [PubMed]
- Peron, D.; Prates, R.A.; Antonio, E.L.; Teixeira, I.L.A.; de Oliveira, H.A.; Mansano, B.S.D.M.; Bergamo, A.; Almeida, D.R.; Dariolli, R.; Tucci, P.J.F.; et al. A common oral pathogen Porphyromonas gingivalis induces myocarditis in rats. J. Clin. Periodontol. 2022, 49, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Okoro, E.U.; Gipson, M.J.; Chukkapalli, S.S.; Farmer-Dixon, C.M.; Gangula, P.R. Porphyromonas gingivalis infection alters Nrf2-phase II enzymes and nitric oxide in primary human aortic endothelial cells. J. Periodontol. 2021, 92, 54–65. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef]
- Tu, W.; Wang, H.; Li, S.; Liu, Q.; Sha, H. The Anti-Inflammatory and Anti-Oxidant Mechanisms of the Keap1/Nrf2/ARE Signaling Pathway in Chronic Diseases. Aging Dis. 2019, 10, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Ohara, N. Novel function of Porphyromonas gingivalis gingipains in the PI3K/Akt signaling pathway. J. Oral Biosci. 2017, 59, 131–134. [Google Scholar] [CrossRef]
- Idrizaj, E.; Traini, C.; Vannucchi, M.G.; Baccari, M.C. Nitric Oxide: From Gastric Motility to Gastric Dysmotility. Int. J. Mol. Sci. 2021, 22, 9990. [Google Scholar] [CrossRef]
- Yan, M.; Hou, M.; Liu, J.; Zhang, S.; Liu, B.; Wu, X.; Liu, G. Regulation of iNOS-Derived ROS Generation by HSP90 and Cav-1 in Porcine Reproductive and Respiratory Syndrome Virus-Infected Swine Lung Injury. Inflammation 2017, 40, 1236–1244. [Google Scholar] [CrossRef]
- Poljakovic, M.; Karpman, D.; Svanborg, C.; Persson, K. Human renal epithelial cells express iNOS in response to cytokines but not bacteria. Kidney Int. 2002, 61, 444–455. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, R.; Yi, Z.; Li, Y.; Fu, Y.; Zhang, Y.; Li, P.; Li, X.; Pan, Y. Porphyromonas gingivalis induced inflammatory responses and promoted apoptosis in lung epithelial cells infected with H1N1 via the Bcl-2/Bax/Caspase-3 signaling pathway. Mol. Med. Rep. 2018, 18, 97–104. [Google Scholar] [CrossRef]
- Nativel, B.; Couret, D.; Giraud, P.; Meilhac, O.; D’Hellencourt, C.L.; Viranaïcken, W.; Da Silva, C.R. Porphyromonas gingivalis lipopolysaccharides act exclusively through TLR4 with a resilience between mouse and human. Sci. Rep. 2017, 7, 15789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Huang, Q.; Deng, S.; Yang, Y.; Li, J.; Wang, S. Mechanisms of TLR4-Mediated Autophagy and Nitroxidative Stress. Front. Cell. Infect. Microbiol. 2021, 11, 766590. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Shen, Y.; Wang, M.; Wang, X.; Yang, Y. Suppression of miR-21 and miR-155 of macrophage by cinnamaldehyde ameliorates ulcerative colitis. Int. Immunopharmacol. 2019, 67, 22–34. [Google Scholar] [CrossRef]
- Tomofuji, T.; Irie, K.; Sanbe, T.; Azuma, T.; Ekuni, D.; Tamaki, N.; Yamamoto, T.; Morita, M. Periodontitis and increase in circulating oxidative stress. Jpn. Dent. Sci. Rev. 2009, 45, 46–51. [Google Scholar] [CrossRef]
- Yamamoto, N.; Kan-O, K.; Tatsuta, M.; Ishii, Y.; Ogawa, T.; Shinozaki, S.; Fukuyama, S.; Nakanishi, Y.; Matsumoto, K. Incense smoke-induced oxidative stress disrupts tight junctions and bronchial epithelial barrier integrity and induces airway hyperresponsiveness in mouse lungs. Sci. Rep. 2021, 11, 7222. [Google Scholar] [CrossRef] [PubMed]
- Komarova, Y.A.; Kruse, K.; Mehta, D.; Malik, A.B. Protein Interactions at Endothelial Junctions and Signaling Mechanisms Regulating Endothelial Permeability. Circ. Res. 2017, 120, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Stathopoulou, P.G.; Galicia, J.C.; Benakanakere, M.R.; Garcia, C.A.; Potempa, J.; Kinane, D.F. Porphyromonas gingivalis induce apoptosis in human gingival epithelial cells through a gingipain-dependent mechanism. BMC Microbiol. 2009, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Bugueno, I.M.; Batool, F.; Korah, L.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis Differentially Modulates Apoptosome Apoptotic Peptidase Activating Factor 1 in Epithelial Cells and Fibroblasts. Am. J. Pathol. 2018, 188, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Meghil, M.M.; Tawfik, O.K.; Elashiry, M.; Rajendran, M.; Arce, R.M.; Fulton, D.J.; Schoenlein, P.V.; Cutler, C.W. Disruption of Immune Homeostasis in Human Dendritic Cells via Regulation of Autophagy and Apoptosis by Porphyromonas gingivalis. Front. Immunol. 2019, 10, 2286. [Google Scholar] [CrossRef] [PubMed]
- Sadaghian Sadabad, M.; von Martels, J.Z.H.; Khan, M.T.; Blokzijl, T.; Paglia, G.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N. A simple coculture system shows mutualism between anaerobic faecalibacteria and epithelial Caco-2 cells. Sci. Rep. 2016, 5, 17906. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb.prot095505. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| IL-1β | 5′-CTTTGAAGCTGATGGCCCTAAA-3′ | 5′-AGTGGTGGTCGGAGATTCGT-3′ |
| IL-6 | 5′-TCAATGAGGAGACTTGCCTG-3′ | 5′-GATGAGTTGTCATGTCCTGC-3′ |
| TLR-4 | 5′-GATCTGTCTCATAATGGCTTG-3′ | 5′-GACAGATTCCGAATGCTTGTG-3′ |
| β-actin | 5′-ATCCTCACCCTGAAGTACCC-3′ | 5′-TAGAAGGTGTGGTGCCAGAT-3′ |
| TNF α | 5′- CACGCTCTTCTGTCTACTGA-3′ | 5′-ATCTGAGTGTGAGGGTCTGG-3′ |
| Sl. No. | Antibody | Source |
|---|---|---|
| 1 | Rabbit anti-occludin | #71–1500, Invitrogen |
| 2 | Rabbit anti-ZO-1 | #61–7300, Invitrogen |
| 3 | Rabbit anti-p-Akt | #9271, Cell Signaling Technology (Danvers, MA, USA) |
| 4 | Mouse anti-Tlr4 | sc-293072, Santa Cruz Biotechnologies (Dallas, TX, USA) |
| 5 | Rabbit anti-β-actin | #4970, Cell Signaling Technology |
| 6 | Mouse anti-Kelch-like ECH-associated protein 1 (Keap-1) | sc-365626, Santa Cruz Biotechnologies |
| 7 | Mouse anti-Nrf2 | sc-365949, Santa Cruz |
| 8 | Mouse anti-GTP cyclohydrolase 1 (GCH-1) | sc-271482, Santa Cruz |
| 9 | Mouse anti-NQO-1 | #sc-376023, Santa Cruz |
| 10 | Rabbit anti-Bax | #2772, Cell Signaling |
| 11 | Rabbit anti-Bcl2 | #2876, Cell Signaling |
| 12 | Rabbit anti-Nfkb (p65) | #ab16502, Abcam (Cambridge, UK) |
| 13 | Mouse anti-IL-1β | #sc-12742, Santa Cruz Biotechnologies |
| 14 | Rabbit anti-iNOS | #ab15323, Abcam |
| 15 | Rabbit anti-PI3K | #4228, Cell Signaling |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampath, C.; Chukkapalli, S.S.; Raju, A.V.; Alluri, L.S.C.; Srisai, D.; Gangula, P.R. Cinnamaldehyde Protects against P. gingivalis Induced Intestinal Epithelial Barrier Dysfunction in IEC-6 Cells via the PI3K/Akt-Mediated NO/Nrf2 Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 4734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094734
Sampath C, Chukkapalli SS, Raju AV, Alluri LSC, Srisai D, Gangula PR. Cinnamaldehyde Protects against P. gingivalis Induced Intestinal Epithelial Barrier Dysfunction in IEC-6 Cells via the PI3K/Akt-Mediated NO/Nrf2 Signaling Pathway. International Journal of Molecular Sciences. 2024; 25(9):4734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094734
Chicago/Turabian StyleSampath, Chethan, Sasanka S. Chukkapalli, Abhinav V. Raju, Leela Subhashini C. Alluri, Dollada Srisai, and Pandu R. Gangula. 2024. "Cinnamaldehyde Protects against P. gingivalis Induced Intestinal Epithelial Barrier Dysfunction in IEC-6 Cells via the PI3K/Akt-Mediated NO/Nrf2 Signaling Pathway" International Journal of Molecular Sciences 25, no. 9: 4734. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094734