
Contributions of Non-Neuronal Cholinergic Systems to the Regulation of Immune Cell Function, Highlighting the Role of α7 Nicotinic Acetylcholine Receptors

Abstract
:1. Introduction
1.1. ACh
1.2. ChAT
1.3. mAChRs and nAChRs
1.3.1. mAChRs
1.3.2. nAChRs
2. Expression of ACh in a Wide Variety of Life Forms
3. Expression of Non-Neuronal Cholinergic Systems in Mammalian Species
4. Cholinergic System in Immune Cells
4.1. ACh Synthesis in Immune Cells
4.1.1. ChAT in Immune Cells
Determination of ACh Synthesizing Activity in Immune Cells
4.1.2. ChAT mRNA and Enzyme Expression in Immune Cells
4.1.3. Mechanisms Regulating ACh Synthesis and Release
4.1.4. Storage and Release of ACh in Immune Cells
4.2. Expression of AChRs in Immune Cells
4.2.1. mAChRs
4.2.2. nAChRs
4.3. Role of α7 nAChRs in Regulation of Immune Function and Inflammatory Responses
4.3.1. Role of α7 nAChRs in the Regulation of Antibody Production
4.3.2. Role of α7 nAChRs in the Regulation of T Cell Differentiation and Cytokine Production
4.3.3. Role of α7 nAChRs in the Promotion of Human CD4+ T Cell Differentiation into Tregs
4.4. Cholinergic Anti-Inflammatory Reflex
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ewins, A.J. Acetylcholine, a New Active Principle of Ergot. Biochem. J. 1914, 8, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Dale, H.H. The action of certain esters and ethers of choline, and their relation to muscarine. J. Pharmacol. Exp. Ther. 1914, 6, 147–190. [Google Scholar]
- Burgen, A.S. The background of the muscarinic system. Life Sci. 1995, 56, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Loewi, O. Über humorale Übertragbarkeit der Herznervenwirkung. Pflügers Arch. Ges. Physiol. 1921, 189, 239–242. [Google Scholar] [CrossRef]
- Loewi, O.; Navratil, E. Über humorale Übertragbarkeit der Herznervenwirkung. X. Mitteilung: Über das Schicksal des Vagusstoffs. Pflügers Arch. Ges. Physiol. 1926, 214, 678–688. [Google Scholar] [CrossRef]
- Dale, H.H.; Dudley, H.W. The presence of histamine and acetylcholine in the spleen of the ox and the horse. J. Physiol. 1929, 68, 97–123. [Google Scholar] [CrossRef]
- Dale, H. Pharmacology and Nerve-endings (Walter Ernest Dixon Memorial Lecture): (Section of Therapeutics and Pharmacology). Proc. R. Soc. Med. 1935, 28, 319–332. [Google Scholar]
- Bellinger, D.L.; Lorton, D.; Hamill, R.W.; Felten, S.Y.; Felten, D.L. Acetylcholinesterase staining and choline acetyltransferase activity in the young adult rat spleen: Lack of evidence for cholinergic innervation. Brain Behav. Immun. 1993, 7, 191–204. [Google Scholar] [CrossRef]
- Girard, E.; Bernard, V.; Minic, J.; Chatonnet, A.; Krejci, E.; Molgo, J. Butyrylcholinesterase and the control of synaptic responses in acetylcholinesterase knockout mice. Life Sci. 2007, 80, 2380–2385. [Google Scholar] [CrossRef]
- Spector, S.; Felix, A.; Semenuk, G.; Finberg, J.P. Development of a specific radioimmunoassay for acetylcholine. J. Neurochem. 1978, 30, 685–689. [Google Scholar] [CrossRef]
- Kawashima, K.; Ishikawa, H.; Mochizuki, M. Radioimmunoassay for acetylcholine in the rat brain. J. Pharmacol. Methods 1980, 3, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Damsma, G.; Westerink, B.H.; Horn, A.S. A simple, sensitive, and economic assay for choline and acetylcholine using HPLC, an enzyme reactor, and an electrochemical detector. J. Neurochem. 1985, 45, 1649–1652. [Google Scholar] [CrossRef] [PubMed]
- Tuček, S. The synthesis of acetylcholine in skeletal muscles of the rat. J. Physiol. 1982, 322, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Fonnum, F. A rapid radiochemical method for the determination of choline acetyltransferase. J. Neurochem. 1975, 24, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Tuček, S. Choline acetyltransferase and the synthesis of acetylcholine. In Handbook of Experimental Pharmacology, The Cholinergic Synapse; Springer: Berlin/Heidelberg, Germany, 1988; Volume 86, pp. 125–165. [Google Scholar] [CrossRef]
- Vijayaraghavan, S.; Karami, A.; Aeinehband, S.; Behbahani, H.; Grandien, A.; Nilsson, B.; Ekdahl, K.N.; Lindblom, R.P.; Piehl, F.; Darreh-Shori, T. Regulated Extracellular Choline Acetyltransferase Activity- The Plausible Missing Link of the Distant Action of Acetylcholine in the Cholinergic Anti-Inflammatory Pathway. PLoS ONE 2013, 8, e65936. [Google Scholar] [CrossRef] [PubMed]
- Gabalski, A.H.; Tynan, A.; Tsaava, T.; Li, J.H.; Lee, D.; Hepler, T.D.; Hide, D.; George, S.; Bravo Iniguez, C.E.; Thompson, D.A.; et al. Circulating extracellular choline acetyltransferase regulates inflammation. J. Intern. Med. 2024, 295, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Shurubor, Y.I.; D’Aurelio, M.; Clark-Matott, J.; Isakova, E.P.; Deryabina, Y.I.; Beal, M.F.; Cooper, A.J.L.; Krasnikov, B.F. Determination of Coenzyme A and Acetyl-Coenzyme A in Biological Samples Using HPLC with UV Detection. Molecules 2017, 22, 1388. [Google Scholar] [CrossRef]
- Speziale, R.; Montesano, C.; De Leonibus, M.L.; Bonelli, F.; Fezzardi, P.; Beconi, M.G.; Monteagudo, E.; Elbaum, D.; Orsatti, L. Determination of acetyl coenzyme A in human whole blood by ultra-performance liquid chromatography-mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1083, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Bonner, T.I.; Buckley, N.J.; Young, A.C.; Brann, M.R. Identification of a family of muscarinic acetylcholine receptor genes. Science 1987, 237, 527–532. [Google Scholar] [CrossRef]
- Bonner, T.I.; Young, A.C.; Brann, M.R.; Buckley, N.J. Cloning and expression of the human and rat m5 muscarinic acetylcholine receptor genes. Neuron 1988, 1, 403–410. [Google Scholar] [CrossRef]
- Hulme, E.C.; Birdsall, N.J.; Buckley, N.J. Muscarinic receptor subtypes. Annu. Rev. Pharmacol. Toxicol. 1990, 30, 633–673. [Google Scholar] [CrossRef]
- Grando, S.A.; Kawashima, K.; Kirkpatrick, C.J.; Kummer, W.; Wessler, I. Recent progress in revealing the biological and medical significance of the non-neuronal cholinergic system. Int. Immunopharmacol. 2015, 29, 1–7. [Google Scholar] [CrossRef]
- Grando, S.A.; Kawashima, K.; Kirkpatrick, C.J.; Meurs, H.; Wessler, I. The non-neuronal cholinergic system: Basic science, therapeutic implications and new perspectives. Life Sci. 2012, 91, 969–972. [Google Scholar] [CrossRef]
- Grando, S.A.; Kawashima, K.; Kirkpatrick, C.J.; Wessler, I. Recent progress in understanding the non-neuronal cholinergic system in humans. Life Sci. 2007, 80, 2181–2185. [Google Scholar] [CrossRef]
- Elgoyhen, A.B.; Katz, E. The efferent medial olivocochlear-hair cell synapse. J. Physiol. Paris 2012, 106, 47–56. [Google Scholar] [CrossRef]
- Zakrzewicz, A.; Richter, K.; Agne, A.; Wilker, S.; Siebers, K.; Fink, B.; Krasteva-Christ, G.; Althaus, M.; Padberg, W.; Hone, A.J.; et al. Canonical and novel non-canonical cholinergic agonists inhibit ATP-induced release of monocytic interleukin-1beta via different combinations of nicotinic acetylcholine receptor subunits alpha7, alpha9 and alpha10. Front. Cell. Neurosci. 2017, 11, 189. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J.; Racke, K. Non-neuronal acetylcholine, a locally acting molecule, widely distributed in biological systems: Expression and function in humans. Pharmacol. Ther. 1998, 77, 59–79. [Google Scholar] [CrossRef]
- Wessler, I.; Kilbinger, H.; Bittinger, F.; Kirkpatrick, C.J. The biological role of non-neuronal acetylcholine in plants and humans. Jpn. J. Pharmacol. 2001, 85, 2–10. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J.; Racke, K. The cholinergic ‘pitfall’: Acetylcholine, a universal cell molecule in biological systems, including humans. Clin. Exp. Pharmacol. Physiol. 1999, 26, 198–205. [Google Scholar] [CrossRef]
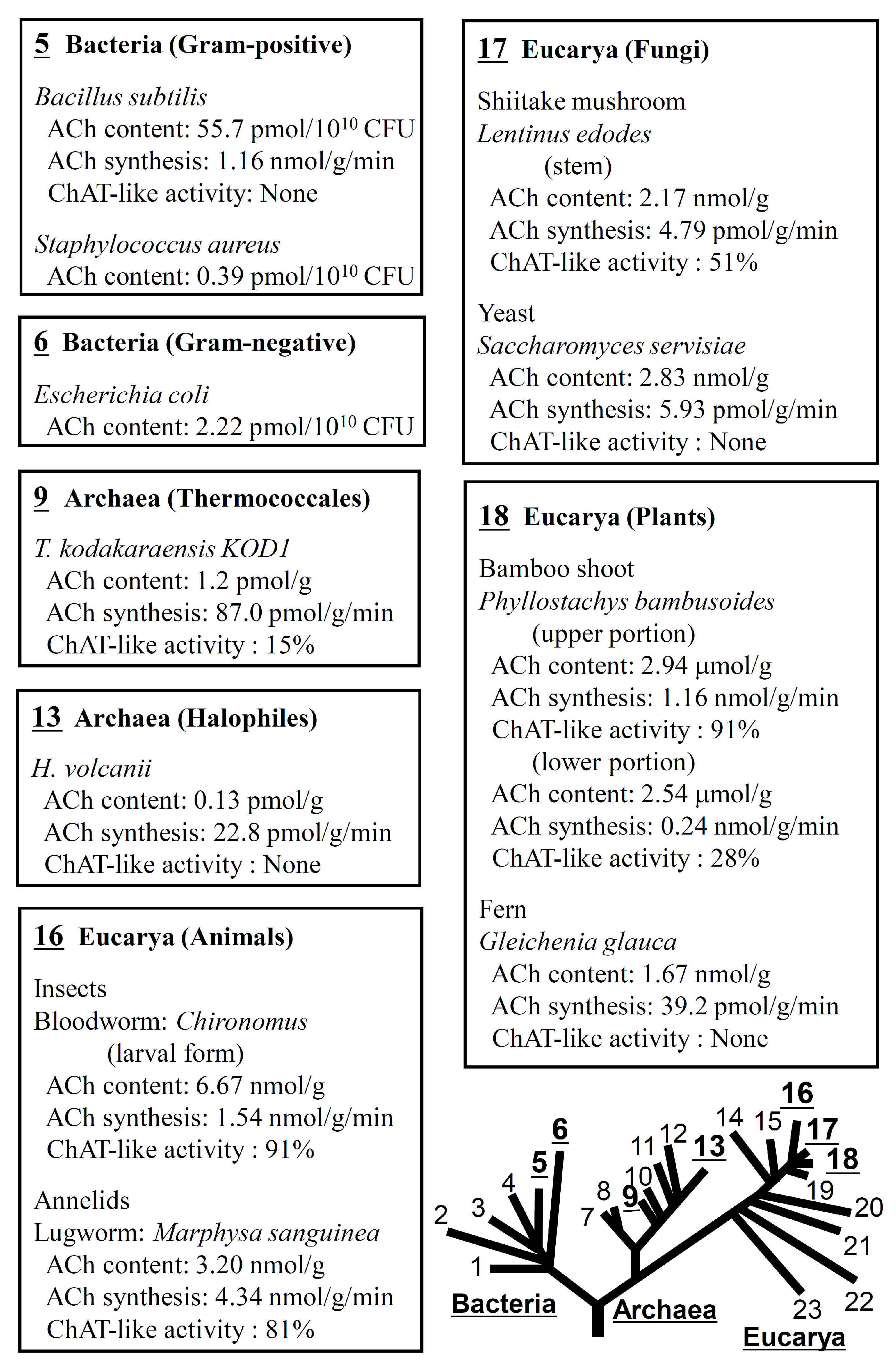
- Horiuchi, Y.; Kimura, R.; Kato, N.; Fujii, T.; Seki, M.; Endo, T.; Kato, T.; Kawashima, K. Evolutional study on acetylcholine expression. Life Sci. 2003, 72, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Fujii, T.; Kanai, T.; Amo, T.; Imanaka, T.; Nishimasu, H.; Wakagi, T.; Shoun, H.; Kamekura, M.; Kamagata, Y.; et al. Expression of acetylcholine (ACh) and ACh-synthesizing activity in Archaea. Life Sci. 2005, 77, 1935–1944. [Google Scholar] [CrossRef]
- Wheelis, M.L.; Kandler, O.; Woese, C.R. On the nature of global classification. Proc. Natl. Acad. Sci. USA 1992, 89, 2930–2934. [Google Scholar] [CrossRef]
- Kawashima, K.; Misawa, H.; Moriwaki, Y.; Fujii, Y.X.; Fujii, T.; Horiuchi, Y.; Yamada, T.; Imanaka, T.; Kamekura, M. Ubiquitous expression of acetylcholine and its biological functions in life forms without nervous systems. Life Sci. 2007, 80, 2206–2209. [Google Scholar] [CrossRef]
- Momonoki, Y.S. Asymmetric distribution of acetylcholinesterase in gravistimulated maize seedlings. Plant Physiol. 1997, 114, 47–53. [Google Scholar] [CrossRef]
- Sagane, Y.; Nakagawa, T.; Yamamoto, K.; Michikawa, S.; Oguri, S.; Momonoki, Y.S. Molecular characterization of maize acetylcholinesterase: A novel enzyme family in the plant kingdom. Plant Physiol. 2005, 138, 1359–1371. [Google Scholar] [CrossRef]
- Sastry, B.V.; Sadavongvivad, C. Cholinergic systems in non-nervous tissues. Pharmacol. Rev. 1978, 30, 65–132. [Google Scholar]
- Grando, S.A.; Kawashima, K.; Wessler, I. Introduction: The non-neuronal cholinergic system in humans. Life Sci. 2003, 72, 2009–2012. [Google Scholar] [CrossRef]
- Grando, S.A.; Kawashima, K.; Wessler, I. A historic perspective on the current progress in elucidation of the biologic significance of non-neuronal acetylcholine. Int. Immunopharmacol. 2020, 81, 106289. [Google Scholar] [CrossRef]
- Grando, S.A.; Kist, D.A.; Qi, M.; Dahl, M.V. Human keratinocytes synthesize, secrete, and degrade acetylcholine. J. Investig. Dermatol. 1993, 101, 32–36. [Google Scholar] [CrossRef]
- Grando, S.A. Biological functions of keratinocyte cholinergic receptors. J. Investig. Dermatol. Symp. Proc. 1997, 2, 41–48. [Google Scholar] [CrossRef]
- Song, P.; Sekhon, H.S.; Jia, Y.; Keller, J.A.; Blusztajn, J.K.; Mark, G.P.; Spindel, E.R. Acetylcholine is synthesized by and acts as an autocrine growth factor for small cell lung carcinoma. Cancer Res. 2003, 63, 214–221. [Google Scholar]
- Cheng, K.; Samimi, R.; Xie, G.; Shant, J.; Drachenberg, C.; Wade, M.; Davis, R.J.; Nomikos, G.; Raufman, J.P. Acetylcholine release by human colon cancer cells mediates autocrine stimulation of cell proliferation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G591–G597. [Google Scholar] [CrossRef]
- Yu, H.; Xia, H.; Tang, Q.; Xu, H.; Wei, G.; Chen, Y.; Dai, X.; Gong, Q.; Bi, F. Acetylcholine acts through M3 muscarinic receptor to activate the EGFR signaling and promotes gastric cancer cell proliferation. Sci. Rep. 2017, 7, 40802. [Google Scholar] [CrossRef]
- Kakinuma, Y.; Akiyama, T.; Sato, T. Cholinoceptive and cholinergic properties of cardiomyocytes involving an amplification mechanism for vagal efferent effects in sparsely innervated ventricular myocardium. FEBS J. 2009, 276, 5111–5125. [Google Scholar] [CrossRef]
- Kawashima, K.; Watanabe, N.; Oohata, H.; Fujimoto, K.; Suzuki, T.; Ishizaki, Y.; Morita, I.; Murota, S. Synthesis and release of acetylcholine by cultured bovine arterial endothelial cells. Neurosci. Lett. 1990, 119, 156–158. [Google Scholar] [CrossRef]
- Ikeda, C.; Morita, I.; Mori, A.; Fujimoto, K.; Suzuki, T.; Kawashima, K.; Murota, S. Phorbol ester stimulates acetylcholine synthesis in cultured endothelial cells isolated from porcine cerebral microvessels. Brain Res. 1994, 655, 147–152. [Google Scholar] [CrossRef]
- Fujii, T.; Tsuchiya, T.; Yamada, S.; Fujimoto, K.; Suzuki, T.; Kasahara, T.; Kawashima, K. Localization and synthesis of acetylcholine in human leukemic T cell lines. J. Neurosci. Res. 1996, 44, 66–72. [Google Scholar] [CrossRef]
- Rinner, I.; Kawashima, K.; Schauenstein, K. Rat lymphocytes produce and secrete acetylcholine in dependence of differentiation and activation. J. Neuroimmunol. 1998, 81, 31–37. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Hall, L.L.; Gallacher, G.; Ndoye, A.; Jolkovsky, D.L.; Webber, R.J.; Buchli, R.; Grando, S.A. Choline acetyltransferase, acetylcholinesterase, and nicotinic acetylcholine receptors of human gingival and esophageal epithelia. J. Dent. Res. 2000, 79, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Klapproth, H.; Reinheimer, T.; Metzen, J.; Munch, M.; Bittinger, F.; Kirkpatrick, C.J.; Hohle, K.D.; Schemann, M.; Racke, K.; Wessler, I. Non-neuronal acetylcholine, a signalling molecule synthezised by surface cells of rat and man. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 355, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Sakuragawa, N.; Elwan, M.A.; Uchida, S.; Fujii, T.; Kawashima, K. Non-neuronal neurotransmitters and neurotrophic factors in amniotic epithelial cells: Expression and function in humans and monkey. Jpn. J. Pharmacol. 2001, 85, 20–23. [Google Scholar] [CrossRef]
- Sastry, B.V. Human placental cholinergic system. Biochem. Pharmacol. 1997, 53, 1577–1586. [Google Scholar] [CrossRef]
- Hamann, M.; Chamoin, M.C.; Portalier, P.; Bernheim, L.; Baroffio, A.; Widmer, H.; Bader, C.R.; Ternaux, J.P. Synthesis and release of an acetylcholine-like compound by human myoblasts and myotubes. J Physiol 1995, 489, 791–803. [Google Scholar] [CrossRef]
- Danielson, P.; Andersson, G.; Alfredson, H.; Forsgren, S. Extensive expression of markers for acetylcholine synthesis and of M2 receptors in tenocytes in therapy-resistant chronic painful patellar tendon tendinosis-a pilot study. Life Sci. 2007, 80, 2235–2238. [Google Scholar] [CrossRef]
- Kurzen, H.; Wessler, I.; Kirkpatrick, C.J.; Kawashima, K.; Grando, S.A. The non-neuronal cholinergic system of human skin. Horm. Metab. Res. 2007, 39, 125–135. [Google Scholar] [CrossRef]
- Yoshida, M.; Inadome, A.; Maeda, Y.; Satoji, Y.; Masunaga, K.; Sugiyama, Y.; Murakami, S. Non-neuronal cholinergic system in human bladder urothelium. Urology 2006, 67, 425–430. [Google Scholar] [CrossRef]
- Hanna-Mitchell, A.T.; Beckel, J.M.; Barbadora, S.; Kanai, A.J.; de Groat, W.C.; Birder, L.A. Non-neuronal acetylcholine and urinary bladder urothelium. Life Sci. 2007, 80, 2298–2302. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T. Extraneuronal cholinergic system in lymphocytes. Pharmacol. Ther. 2000, 86, 29–48. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T. The lymphocytic cholinergic system and its contribution to the regulation of immune activity. Life Sci. 2003, 74, 675–696. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T. Expression of non-neuronal acetylcholine in lymphocytes and its contribution to the regulation of immune function. Front. Biosci. 2004, 9, 2063–2085. [Google Scholar] [CrossRef]
- Bany, U.; Laskowska-Bozek, H.; Ryzewski, J. Accumulation of inositol 1,4,5-triphosphate after cholinergic stimulation of human lymphocytes. Arch. Immunol. Ther. Exp. 1995, 43, 51–54. [Google Scholar]
- Maslinski, W. Cholinergic receptors of lymphocytes. Brain Behav. Immun. 1989, 3, 1–14. [Google Scholar] [CrossRef]
- Rinner, I.; Schauenstein, K. The parasympathetic nervous system takes part in the immuno-neuroendocrine dialogue. J. Neuroimmunol. 1991, 34, 165–172. [Google Scholar] [CrossRef]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Expression and Function of the Cholinergic System in Immune Cells. Front. Immunol. 2017, 8, 1085. [Google Scholar] [CrossRef]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Physiological functions of the cholinergic system in immune cells. J. Pharmacol. Sci. 2017, 134, 1–21. [Google Scholar] [CrossRef]
- Kawashima, K.; Oohata, H.; Fujimoto, K.; Suzuki, T. Plasma concentration of acetylcholine in young women. Neurosci. Lett. 1987, 80, 339–342. [Google Scholar] [CrossRef]
- Kawashima, K.; Oohata, H.; Fujimoto, K.; Suzuki, T. Extraneuronal localization of acetylcholine and its release upon nicotinic stimulation in rabbits. Neurosci. Lett. 1989, 104, 336–339. [Google Scholar] [CrossRef]
- Rinner, I.; Schauenstein, K. Detection of choline-acetyltransferase activity in lymphocytes. J. Neurosci. Res. 1993, 35, 188–191. [Google Scholar] [CrossRef]
- Kawashima, K.; Kajiyama, K.; Fujimoto, K.; Oohata, H.; Suzuki, T. Presence of acetylcholine in human blood and its localization in circulating mononuclear leukocytes. Biog. Amine 1993, 9, 251–258. [Google Scholar]
- Fujii, T.; Yamada, S.; Misawa, H.; Tajima, S.; Fujimoto, K.; Suzuki, T.; Kawashima, K. Expression of choline acetyltransferase mRNA and protein in t-lymphocytes. Proc. Jpn. Acad. 1995, 71B, 231–235. [Google Scholar] [CrossRef]
- Fujii, T.; Yamada, S.; Watanabe, Y.; Misawa, H.; Tajima, S.; Fujimoto, K.; Kasahara, T.; Kawashima, K. Induction of choline acetyltransferase mRNA in human mononuclear leukocytes stimulated by phytohemagglutinin, a T-cell activator. J. Neuroimmunol. 1998, 82, 101–107. [Google Scholar] [CrossRef]
- Fujii, T.; Tajima, S.; Yamada, S.; Watanabe, Y.; Sato, K.Z.; Matsui, M.; Misawa, H.; Kasahara, T.; Kawashima, K. Constitutive expression of mRNA for the same choline acetyltransferase as that in the nervous system, an acetylcholine-synthesizing enzyme, in human leukemic T-cell lines. Neurosci. Lett. 1999, 259, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Matsui, M.; Fujii, T.; Kawashima, K. Decreased acetylcholine content and choline acetyltransferase mRNA expression in circulating mononuclear leukocytes and lymphoid organs of the spontaneously hypertensive rat. Life Sci. 2001, 69, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Yoshikawa, K.; Fujii, Y.X.; Moriwaki, Y.; Misawa, H. Expression and function of genes encoding cholinergic components in murine immune cells. Life Sci. 2007, 80, 2314–2319. [Google Scholar] [CrossRef] [PubMed]
- Salamone, G.; Lombardi, G.; Gori, S.; Nahmod, K.; Jancic, C.; Amaral, M.M.; Vermeulen, M.; Espanol, A.; Sales, M.E.; Geffner, J. Cholinergic modulation of dendritic cell function. J. Neuroimmunol. 2011, 236, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Tallini, Y.N.; Shui, B.; Greene, K.S.; Deng, K.Y.; Doran, R.; Fisher, P.J.; Zipfel, W.; Kotlikoff, M.I. BAC transgenic mice express enhanced green fluorescent protein in central and peripheral cholinergic neurons. Physiol. Genom. 2006, 27, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Gautron, L.; Rutkowski, J.M.; Burton, M.D.; Wei, W.; Wan, Y.; Elmquist, J.K. Neuronal and nonneuronal cholinergic structures in the mouse gastrointestinal tract and spleen. J. Comp. Neurol. 2013, 521, 3741–3767. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Takada-Takatori, Y.; Kawashima, K. Roles played by lymphocyte function-associated antigen-1 in the regulation of lymphocytic cholinergic activity. Life Sci. 2007, 80, 2320–2324. [Google Scholar] [CrossRef]
- Fujii, T.; Takada-Takatori, Y.; Kawashima, K. Regulatory mechanisms of acetylcholine synthesis and release by T cells. Life Sci. 2012, 91, 981–985. [Google Scholar] [CrossRef]
- Fujii, T.; Watanabe, Y.; Inoue, T.; Kawashima, K. Upregulation of mRNA encoding the M5 muscarinic acetylcholine receptor in human T- and B-lymphocytes during immunological responses. Neurochem. Res. 2003, 28, 423–429. [Google Scholar] [CrossRef]
- Fujii, T.; Horiguchi, K.; Sunaga, H.; Moriwaki, Y.; Misawa, H.; Kasahara, T.; Tsuji, S.; Kawashima, K. SLURP-1, an endogenous alpha7 nicotinic acetylcholine receptor allosteric ligand, is expressed in CD205(+) dendritic cells in human tonsils and potentiates lymphocytic cholinergic activity. J. Neuroimmunol. 2014, 267, 43–49. [Google Scholar] [CrossRef]
- Cox, M.A.; Duncan, G.S.; Lin, G.H.Y.; Steinberg, B.E.; Yu, L.X.; Brenner, D.; Buckler, L.N.; Elia, A.J.; Wakeham, A.C.; Nieman, B.; et al. Choline acetyltransferase-expressing T cells are required to control chronic viral infection. Science 2019, 363, 639–644. [Google Scholar] [CrossRef]
- Usdin, T.B.; Eiden, L.E.; Bonner, T.I.; Erickson, J.D. Molecular biology of the vesicular ACh transporter. Trends Neurosci. 1995, 18, 218–224. [Google Scholar] [CrossRef]
- Alfonso, A.; Grundahl, K.; McManus, J.R.; Asbury, J.M.; Rand, J.B. Alternative splicing leads to two cholinergic proteins in Caenorhabditis elegans. J. Mol. Biol. 1994, 241, 627–630. [Google Scholar] [CrossRef]
- Bejanin, S.; Cervini, R.; Mallet, J.; Berrard, S. A unique gene organization for two cholinergic markers, choline acetyltransferase and a putative vesicular transporter of acetylcholine. J. Biol. Chem. 1994, 269, 21944–21947. [Google Scholar] [CrossRef]
- Erickson, J.D.; Varoqui, H.; Schafer, M.K.; Modi, W.; Diebler, M.F.; Weihe, E.; Rand, J.; Eiden, L.E.; Bonner, T.I.; Usdin, T.B. Functional identification of a vesicular acetylcholine transporter and its expression from a “cholinergic” gene locus. J. Biol. Chem. 1994, 269, 21929–21932. [Google Scholar] [CrossRef]
- Berrard, S.; Varoqui, H.; Cervini, R.; Israel, M.; Mallet, J.; Diebler, M.F. Coregulation of two embedded gene products, choline acetyltransferase and the vesicular acetylcholine transporter. J. Neurochem. 1995, 65, 939–942. [Google Scholar] [CrossRef]
- Berse, B.; Blusztajn, J.K. Coordinated up-regulation of choline acetyltransferase and vesicular acetylcholine transporter gene expression by the retinoic acid receptor alpha, cAMP, and leukemia inhibitory factor/ciliary neurotrophic factor signaling pathways in a murine septal cell line. J. Biol. Chem. 1995, 270, 22101–22104. [Google Scholar] [CrossRef]
- Misawa, H.; Takahashi, R.; Deguchi, T. Coordinate expression of vesicular acetylcholine transporter and choline acetyltransferase in sympathetic superior cervical neurones. Neuroreport 1995, 6, 965–968. [Google Scholar] [CrossRef]
- Tian, X.; Sun, X.; Suszkiw, J.B. Developmental age-dependent upregulation of choline acetyltransferase and vesicular acetylcholine transporter mRNA expression in neonatal rat septum by nerve growth factor. Neurosci. Lett. 1996, 209, 134–136. [Google Scholar] [CrossRef]
- Cavalli, A.; Eder-Colli, L.; Dunant, Y.; Loctin, F.; Morel, N. Release of acetylcholine by Xenopus oocytes injected with mRNAs from cholinergic neurons. EMBO J. 1991, 10, 1671–1675. [Google Scholar] [CrossRef] [PubMed]
- Israël, M.; Dunant, Y. Acetylcholine release and the cholinergic genomic locus. Mol. Neurobiol. 1998, 16, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Malo, M.; Israël, M. Expression of the acetylcholine release mechanism in various cells and reconstruction of the release mechanism in non-releasing cells. Life Sci. 2003, 72, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Dunant, Y.; Cordeiro, J.M.; Goncalves, P.P. Exocytosis, mediatophore, and vesicular Ca2+/H+ antiport in rapid neurotransmission. Ann. N. Y. Acad. Sci. 2009, 1152, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Takada-Takatori, Y.; Horiguchi, K.; Kawashima, K. Mediatophore regulates acetylcholine release from T cells. J. Neuroimmunol. 2012, 244, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.Z.; Fujii, T.; Watanabe, Y.; Yamada, S.; Ando, T.; Kazuko, F.; Kawashima, K. Diversity of mRNA expression for muscarinic acetylcholine receptor subtypes and neuronal nicotinic acetylcholine receptor subunits in human mononuclear leukocytes and leukemic cell lines. Neurosci. Lett. 1999, 266, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Fujii, T.; Moriwaki, Y.; Misawa, H.; Horiguchi, K. Reconciling neuronally and nonneuronally derived acetylcholine in the regulation of immune function. Ann. N. Y. Acad. Sci. 2012, 1261, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Kawashima, K. Calcium signaling and c-Fos gene expression via M3 muscarinic acetylcholine receptors in human T- and B-cells. Jpn. J. Pharmacol. 2000, 84, 124–132. [Google Scholar] [CrossRef]
- Fujii, T.; Kawashima, K. Ca2+ oscillation and c-fos gene expression induced via muscarinic acetylcholine receptor in human T- and B-cell lines. Naunyn Schmiedebergs Arch. Pharmacol. 2000, 362, 14–21. [Google Scholar] [CrossRef]
- Fujii, Y.X.; Tashiro, A.; Arimoto, K.; Fujigaya, H.; Moriwaki, Y.; Misawa, H.; Fujii, T.; Matsui, M.; Kasahara, T.; Kawashima, K. Diminished antigen-specific IgG1 and interleukin-6 production and acetylcholinesterase expression in combined M1 and M5 muscarinic acetylcholine receptor knockout mice. J. Neuroimmunol. 2007, 188, 80–85. [Google Scholar] [CrossRef]
- Koarai, A.; Traves, S.L.; Fenwick, P.S.; Brown, S.M.; Chana, K.K.; Russell, R.E.; Nicholson, A.G.; Barnes, P.J.; Donnelly, L.E. Expression of muscarinic receptors by human macrophages. Eur. Respir. J. 2012, 39, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Galitovskiy, V.; Chernyavsky, A.I.; Marchenko, S.; Grando, S.A. Plasticity of the murine spleen T-cell cholinergic receptors and their role in in vitro differentiation of naive CD4 T cells toward the Th1, Th2 and Th17 lineages. Genes Immun. 2011, 12, 222–230. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T.; Moriwaki, Y.; Misawa, H.; Horiguchi, K. Non-neuronal cholinergic system in regulation of immune function with a focus on alpha7 nAChRs. Int. Immunopharmacol. 2015, 29, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, M.; Fujii, T.; Ono, S.; Moriwaki, Y.; Misawa, H.; Kawashima, K. Minireview: Divergent roles of alpha7 nicotinic acetylcholine receptors expressed on antigen-presenting cells and CD4(+) T cells in the regulation of T cell differentiation. Int. Immunopharmacol. 2020, 82, 106306. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, M.; Komori, M.; Matsui, Y.Y.; Murase, M.X.; Fujii, T.; Takeshima, S.; Okuyama, H.; Ono, S.; Moriwaki, Y.; Misawa, H.; et al. Distinct Roles of alpha7 nAChRs in Antigen-Presenting Cells and CD4(+) T Cells in the Regulation of T Cell Differentiation. Front. Immunol. 2019, 10, 1102. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.X.; Fujigaya, H.; Moriwaki, Y.; Misawa, H.; Kasahara, T.; Grando, S.A.; Kawashima, K. Enhanced serum antigen-specific IgG1 and proinflammatory cytokine production in nicotinic acetylcholine receptor alpha7 subunit gene knockout mice. J. Neuroimmunol. 2007, 189, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Skok, M.V.; Grailhe, R.; Agenes, F.; Changeux, J.P. The role of nicotinic receptors in B-lymphocyte development and activation. Life Sci. 2007, 80, 2334–2336. [Google Scholar] [CrossRef] [PubMed]
- Nizri, E.; Irony-Tur-Sinai, M.; Lory, O.; Orr-Urtreger, A.; Lavi, E.; Brenner, T. Activation of the cholinergic anti-inflammatory system by nicotine attenuates neuroinflammation via suppression of Th1 and Th17 responses. J. Immunol. 2009, 183, 6681–6688. [Google Scholar] [CrossRef]
- Nizri, E.; Brenner, T. Modulation of inflammatory pathways by the immune cholinergic system. Amino Acids 2013, 45, 73–85. [Google Scholar] [CrossRef]
- Galitovskiy, V.; Qian, J.; Chernyavsky, A.I.; Marchenko, S.; Gindi, V.; Edwards, R.A.; Grando, S.A. Cytokine-induced alterations of alpha7 nicotinic receptor in colonic CD4 T cells mediate dichotomous response to nicotine in murine models of Th1/Th17- versus Th2-mediated colitis. J. Immunol. 2011, 187, 2677–2687. [Google Scholar] [CrossRef]
- Gault, J.; Robinson, M.; Berger, R.; Drebing, C.; Logel, J.; Hopkins, J.; Moore, T.; Jacobs, S.; Meriwether, J.; Choi, M.J.; et al. Genomic organization and partial duplication of the human alpha7 neuronal nicotinic acetylcholine receptor gene (CHRNA7). Genomics 1998, 52, 173–185. [Google Scholar] [CrossRef]
- Riley, B.; Williamson, M.; Collier, D.; Wilkie, H.; Makoff, A. A 3-Mb map of a large Segmental duplication overlapping the alpha7-nicotinic acetylcholine receptor gene (CHRNA7) at human 15q13-q14. Genomics 2002, 79, 197–209. [Google Scholar] [CrossRef]
- de Lucas-Cerrillo, A.M.; Maldifassi, M.C.; Arnalich, F.; Renart, J.; Atienza, G.; Serantes, R.; Cruces, J.; Sanchez-Pacheco, A.; Andres-Mateos, E.; Montiel, C. Function of partially duplicated human alpha7 nicotinic receptor subunit CHRFAM7A gene: Potential implications for the cholinergic anti-inflammatory response. J. Biol. Chem. 2011, 286, 594–606. [Google Scholar] [CrossRef]
- Araud, T.; Graw, S.; Berger, R.; Lee, M.; Neveu, E.; Bertrand, D.; Leonard, S. The chimeric gene CHRFAM7A, a partial duplication of the CHRNA7 gene, is a dominant negative regulator of alpha7*nAChR function. Biochem. Pharmacol. 2011, 82, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Lasala, M.; Corradi, J.; Bruzzone, A.; Esandi, M.D.; Bouzat, C. A human-specific, truncated 7 nicotinic receptor subunit assembles with full-length 7 and forms functional receptors with different stoichiometries. J. Biol. Chem. 2018, 293, 10707–10717. [Google Scholar] [CrossRef]
- Sinkus, M.L.; Graw, S.; Freedman, R.; Ross, R.G.; Lester, H.A.; Leonard, S. The human CHRNA7 and CHRFAM7A genes: A review of the genetics, regulation, and function. Neuropharmacology 2015, 96, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, C.; Indersmitten, T.; Freedman, R.; Leonard, S.; Lester, H.A. The duplicated alpha7 subunits assemble and form functional nicotinic receptors with the full-length alpha7. J. Biol. Chem. 2014, 289, 26451–26463. [Google Scholar] [CrossRef] [PubMed]
- Costantini, T.W.; Dang, X.; Yurchyshyna, M.V.; Coimbra, R.; Eliceiri, B.P.; Baird, A. A Human-Specific alpha7-Nicotinic Acetylcholine Receptor Gene in Human Leukocytes: Identification, Regulation and the Consequences of CHRFAM7A Expression. Mol. Med. 2015, 21, 323–336. [Google Scholar] [CrossRef]
- Kabbani, N.; Nordman, J.C.; Corgiat, B.A.; Veltri, D.P.; Shehu, A.; Seymour, V.A.; Adams, D.J. Are nicotinic acetylcholine receptors coupled to G proteins? Bioessays 2013, 35, 1025–1034. [Google Scholar] [CrossRef]
- de Jonge, W.J.; Ulloa, L. The alpha7 nicotinic acetylcholine receptor as a pharmacological target for inflammation. Br. J. Pharmacol. 2007, 151, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Razani-Boroujerdi, S.; Boyd, R.T.; Davila-Garcia, M.I.; Nandi, J.S.; Mishra, N.C.; Singh, S.P.; Pena-Philippides, J.C.; Langley, R.; Sopori, M.L. T cells express alpha7-nicotinic acetylcholine receptor subunits that require a functional TCR and leukocyte-specific protein tyrosine kinase for nicotine-induced Ca2+ response. J. Immunol. 2007, 179, 2889–2898. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, M.; Takeshima, S.; Okuyama, H.; Matsurida, A.; Murase, M.; Ono, S.; Kawashima, K.; Fujii, T. alpha7 nAChRs expressed on antigen presenting cells are insensitive to the conventional antagonists alpha-bungarotoxin and methyllycaconitine. Int. Immunopharmacol. 2020, 81, 106276. [Google Scholar] [CrossRef] [PubMed]
- King, J.R.; Nordman, J.C.; Bridges, S.P.; Lin, M.K.; Kabbani, N. Identification and characterization of a G protein-binding cluster in alpha7 nicotinic acetylcholine receptors. J. Biol. Chem. 2015, 290, 20060–20070. [Google Scholar] [CrossRef]
- Papke, R.L.; Lindstrom, J.M. Nicotinic acetylcholine receptors: Conventional and unconventional ligands and signaling. Neuropharmacology 2020, 168, 108021. [Google Scholar] [CrossRef]
- Hao, J.; Simard, A.R.; Turner, G.H.; Wu, J.; Whiteaker, P.; Lukas, R.J.; Shi, F.D. Attenuation of CNS inflammatory responses by nicotine involves alpha7 and non-alpha7 nicotinic receptors. Exp. Neurol. 2011, 227, 110–119. [Google Scholar] [CrossRef]
- Reale, M.; Costantini, E. Cholinergic Modulation of the Immune System in Neuroinflammatory Diseases. Diseases 2021, 9, 29. [Google Scholar] [CrossRef]
- Benfante, R.; Antonini, R.A.; De Pizzol, M.; Gotti, C.; Clementi, F.; Locati, M.; Fornasari, D. Expression of the alpha7 nAChR subunit duplicate form (CHRFAM7A) is down-regulated in the monocytic cell line THP-1 on treatment with LPS. J. Neuroimmunol. 2011, 230, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Villiger, Y.; Szanto, I.; Jaconi, S.; Blanchet, C.; Buisson, B.; Krause, K.H.; Bertrand, D.; Romand, J.A. Expression of an alpha7 duplicate nicotinic acetylcholine receptor-related protein in human leukocytes. J. Neuroimmunol. 2002, 126, 86–98. [Google Scholar] [CrossRef]
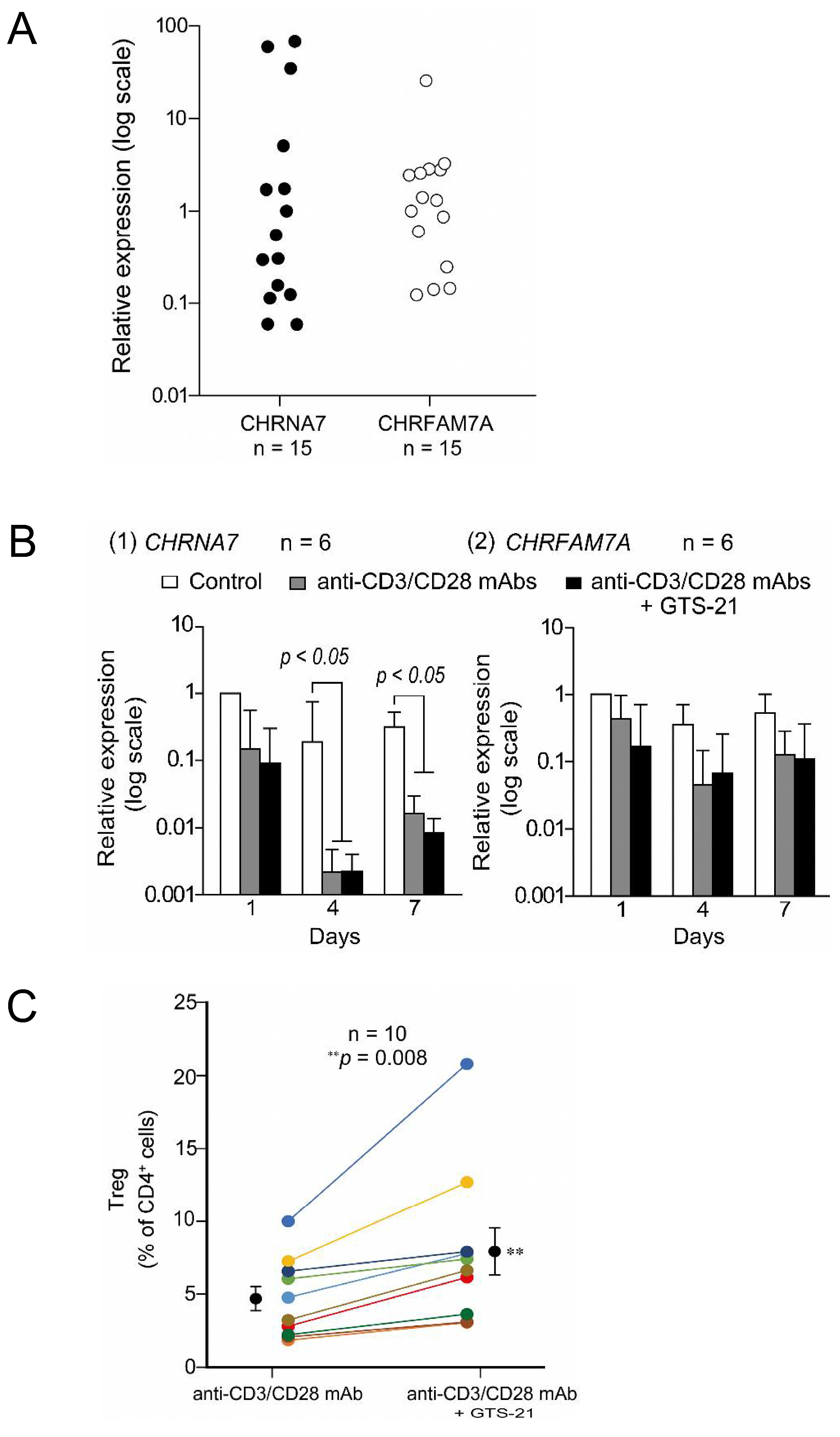
- Mashimo, M.; Fujii, T.; Ono, S.; Moriwaki, Y.; Misawa, H.; Azami, T.; Kasahara, T.; Kawashima, K. GTS-21 Enhances Regulatory T Cell Development from T Cell Receptor-Activated Human CD4(+) T Cells Exhibiting Varied Levels of CHRNA7 and CHRFAM7A Expression. Int. J. Mol. Sci. 2023, 24, 12257. [Google Scholar] [CrossRef]
- Dang, X.; Eliceiri, B.P.; Baird, A.; Costantini, T.W. CHRFAM7A: A human-specific alpha7-nicotinic acetylcholine receptor gene shows differential responsiveness of human intestinal epithelial cells to LPS. FASEB J. 2015, 29, 2292–2302. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Nance, D.M.; Sanders, V.M. Autonomic innervation and regulation of the immune system (1987–2007). Brain Behav. Immun. 2007, 21, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Ballina, M.; Olofsson, P.S.; Ochani, M.; Valdes-Ferrer, S.I.; Levine, Y.A.; Reardon, C.; Tusche, M.W.; Pavlov, V.A.; Andersson, U.; Chavan, S.; et al. Acetylcholine-synthesizing T cells relay neural signals in a vagus nerve circuit. Science 2011, 334, 98–101. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, W.J.; van der Zanden, E.P.; The, F.O.; Bijlsma, M.F.; van Westerloo, D.J.; Bennink, R.J.; Berthoud, H.R.; Uematsu, S.; Akira, S.; van den Wijngaard, R.M.; et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nat. Immunol. 2005, 6, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Parrish, W.R.; Rosas-Ballina, M.; Gallowitsch-Puerta, M.; Ochani, M.; Ochani, K.; Yang, L.H.; Hudson, L.; Lin, X.; Patel, N.; Johnson, S.M.; et al. Modulation of TNF release by choline requires alpha7 subunit nicotinic acetylcholine receptor-mediated signaling. Mol. Med. 2008, 14, 567–574. [Google Scholar] [CrossRef]
- Besedovsky, H.O.; del Rey, A. Immune-neuro-endocrine interactions: Facts and hypotheses. Endocr. Rev. 1996, 17, 64–102. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| 1. Cancer cells |
| (1) Lung cancer cells [42] |
| (2) Colon cancer cells [43] |
| (3) Stomach cancer cells [44] |
| 2. Cardiovascular cells |
| (1) Cardiomyocytes [45] |
| (2) Vascular endothelial cells [46,47] |
| 3. Immune cells (T cells, B cells, and Monocytes) [48,49] |
| 4. Digestive epithelial cells |
| (1) Gingival and esophageal epithelial cells [50] |
| (2) Small intestinal epithelial cells [51] |
| 5. Reproductive organs |
| (1) Amniotic membrane [52] |
| (2) Placenta [53] |
| 6. Respiratory epithelial cells |
| (1) Bronchial epithelial cells [51] |
| 7. Myogenic cells and tendon |
| (1) Myogenic cells [54] |
| (2) Tendon [55] |
| 8. Skin |
| (1) Keratinocytes [41]; see also a review by Kurzen et al. [56] |
| 9. Urinary bladder [57,58] |
| (A) mAChR | ||||||
| Sample | Cell Type | M1 | M2 | M3 | M4 | M5 |
| 1 (F) | MNLs | + | + | + | + | + |
| 2 (F) | MNLs | − | + | − | + | + |
| 3 (F) | MNLs | + | + | + | + | + |
| 4 (F) | MNLs | + | − | + | + | + |
| 5 (M) | MNLs | + | + | − | + | + |
| 6 (M) | MNLs | + | − | + | + | + |
| 7 (M) | MNLs | − | + | + | + | + |
| (B) nAChR | ||||||
| Sample | Cell Type | α3 | α5 | α7 | α9 | α10 |
| 1 (F) | T | + | + | + | + | + |
| B | + | + | + | − | + | |
| 2 (F) | T | + | + | − | + | − |
| B | + | + | + | + | + | |
| 3 (F) | T | + | + | − | − | − |
| B | + | + | + | − | − | |
| 4 (F) | T | + | + | + | − | − |
| B | + | + | + | + | + | |
| 5 (F) | T | + | + | + | + | − |
| B | + | − | − | + | + | |
| 6 (F) | T | − | + | − | + | − |
| B | + | + | + | + | + | |
| 7 (F) | T | + | + | + | + | + |
| B | − | + | + | + | + | |
| 8 (F) | T | − | + | + | + | + |
| B | + | + | + | + | + | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawashima, K.; Mashimo, M.; Nomura, A.; Fujii, T. Contributions of Non-Neuronal Cholinergic Systems to the Regulation of Immune Cell Function, Highlighting the Role of α7 Nicotinic Acetylcholine Receptors. Int. J. Mol. Sci. 2024, 25, 4564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084564
Kawashima K, Mashimo M, Nomura A, Fujii T. Contributions of Non-Neuronal Cholinergic Systems to the Regulation of Immune Cell Function, Highlighting the Role of α7 Nicotinic Acetylcholine Receptors. International Journal of Molecular Sciences. 2024; 25(8):4564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084564
Chicago/Turabian StyleKawashima, Koichiro, Masato Mashimo, Atsuo Nomura, and Takeshi Fujii. 2024. "Contributions of Non-Neuronal Cholinergic Systems to the Regulation of Immune Cell Function, Highlighting the Role of α7 Nicotinic Acetylcholine Receptors" International Journal of Molecular Sciences 25, no. 8: 4564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25084564