Development of the 12-Base Short Dimeric Myogenetic Oligodeoxynucleotide That Induces Myogenic Differentiation

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oligonucleotides
2.2. Myoblast Culture
2.3. Immunocytochemistry
2.4. qPCR
2.5. Protein Precipitation and Western Blotting
2.6. Agarose Gel Electrophoresis
2.7. NMR
2.8. Native PAGE
2.9. Multicanonical Molecular Dynamics (McMD) Simulation
2.10. Statistical Analysis
3. Results
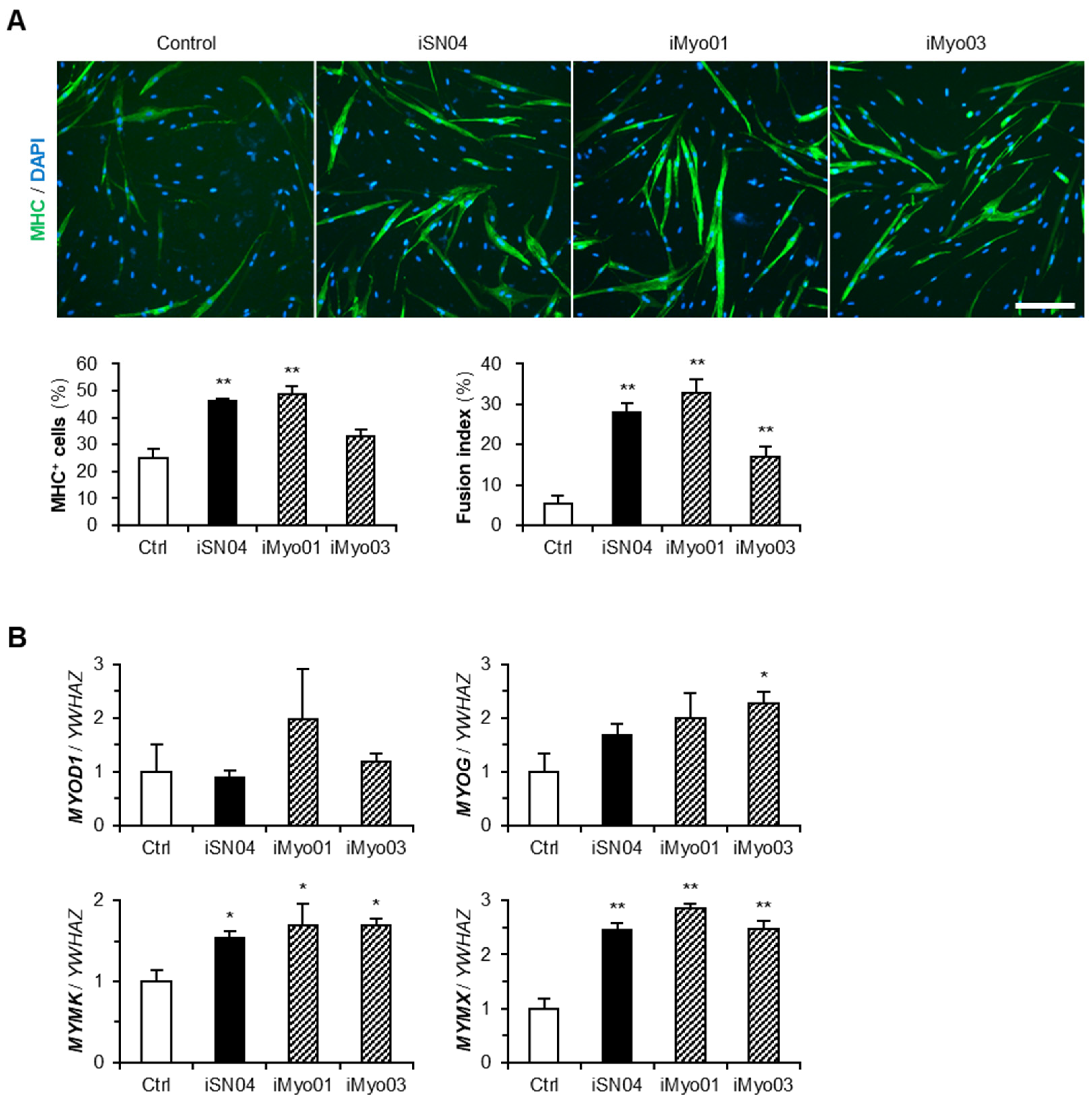
3.1. Designing and Screening of iMyo-ODNs
3.2. iMyo01 and iMyo03 Bind to Nucleolin but Not to Berberine
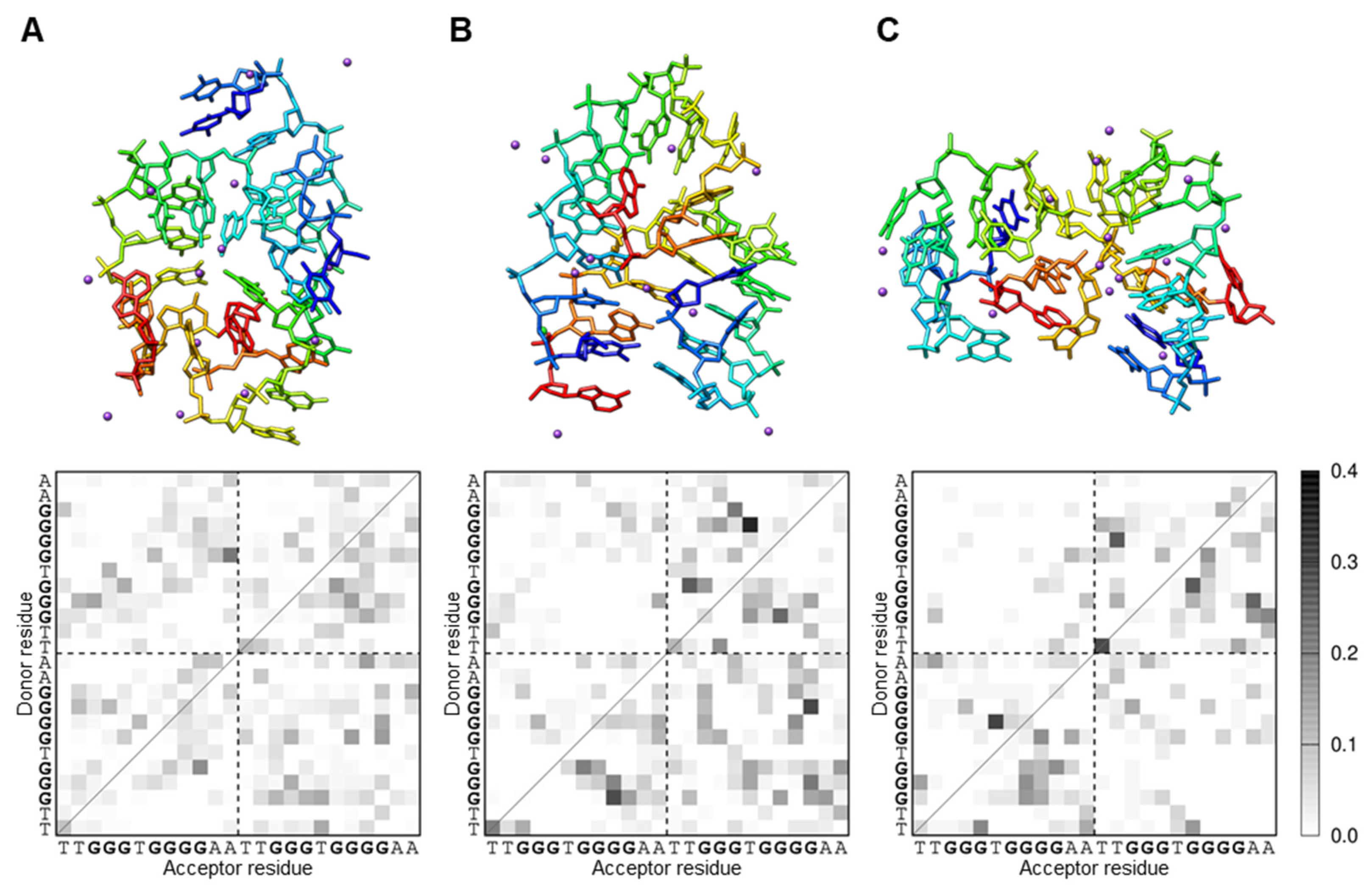
3.3. iMyo01 Forms a G-Quadruplex Structure
3.4. iMyo01 Forms a Homodimer in the Presence of K+
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.H.; Wang, H.; Du, J.; Mitch, W.E. Satellite cell dysfunction and impaired IGF-1 signaling cause CKD-induced muscle atrophy. J. Am. Soc. Nephrol. 2010, 21, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Marchildon, F.; Lamarche, E.; Lala-Tabbert, N.; St-Louis, C.; Wiper-Bergeron, N. Expression of CCAAT/enhancer binding protein beta in muscle satellite cells inhibits myogenesis in cancer cachexia. PLoS ONE 2015, 10, e0145583. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, T.I.; Davidsen, P.K.; Pedersen, M.; Schultz, H.S.; Hansen, N.S.; Larsen, T.J.; Vaag, A.; Pedersen, B.K.; Nielsen, S.; Scheele, C. Dysregulation of a novel miR-23b/27b-p53 axis impairs muscle stem cell differentiation of humans with type 2 diabetes. Mol. Metab. 2017, 6, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, C.; Larcher, L.M.; Barrero, R.A.; Veedu, R.N. Three decades of nucleic acid aptamer technologies: Lessons learned, progress and opportunities on aptamer development. Biotechnol. Adv. 2019, 37, 28–50. [Google Scholar] [CrossRef] [PubMed]
- Shinji, S.; Umezawa, K.; Nihashi, Y.; Nakamura, S.; Shimosato, T.; Takaya, T. Identification of the myogenetic oligodeoxynucleotides (myoDNs) that promote differentiation of skeletal muscle myoblasts by targeting nucleolin. Front. Cell Dev. Biol. 2021, 8, 616706. [Google Scholar] [CrossRef] [PubMed]
- Nohira, N.; Shinji, S.; Nakamura, S.; Nihashi, Y.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotides as anti-nucleolin aptamers inhibit the growth of embryonal rhabdomyosarcoma cells. Biomedicines 2022, 10, 2691. [Google Scholar] [CrossRef] [PubMed]
- Nihashi, Y.; Shinji, S.; Umezawa, K.; Shimosato, T.; Ono, T.; Kagami, H.; Ono, T.; Kagami, H.; Takaya, T. Myogenetic oligodeoxynucleotide complexed with berberine promotes differentiation of chicken myoblasts. Anim. Sci. J. 2021, 92, e13597. [Google Scholar] [CrossRef]
- Nakamura, S.; Yonekura, S.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotide (myoDN) recovers the differentiation of skeletal muscle myoblasts deteriorated by diabetes mellitus. Front. Physiol. 2021, 12, 679152. [Google Scholar] [CrossRef]
- Nihashi, Y.; Yamamoto, M.; Shimosato, T.; Takaya, T. Myogenetic oligodeoxynucleotide restores differentiation and reverses inflammation of myoblasts aggravated by cancer-conditioned medium. Muscles 2022, 1, 111–120. [Google Scholar] [CrossRef]
- Yamamoto, M.; Miyoshi, M.; Morioka, K.; Mitani, T.; Takaya, T. Anti-nucleolin aptamer, iSN04, inhibits the inflammatory responses in C2C12 myoblasts by modulating the β-catenin/NF-κB signaling pathway. Biochem. Biophys. Res. Commun. 2023, 664, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ishioka, M.; Nihashi, Y.; Sunagawa, Y.; Umezawa, K.; Shimosato, T.; Kagami, H.; Morimoto, T.; Takaya, T. Myogenetic oligodeoxynucleotide induces myocardial differentiation of murine pluripotent stem cells. Int. J. Mol. Sci. 2023, 24, 14380. [Google Scholar] [CrossRef] [PubMed]
- Juliano, R.L. Intracellular trafficking and endosomal release of oligonucleotides: What we know and what we don’t. Nucleic Acid Ther. 2018, 28, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Rajput, Y.S.; Singh, D.; Sharma, R. Prediction of shorter oligonucleotide sequences recognizing aflatoxin M1. Biotechnol. Appl. Biochem. 2018, 65, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Nihashi, Y.; Miyoshi, M.; Umezawa, K.; Shimosato, T.; Takaya, T. Identification of a novel osteogenetic oligodeoxynucleotide (osteoDN) that promotes osteoblast differentiation in a TLR9-independent manner. Nanomaterials 2022, 12, 1680. [Google Scholar] [CrossRef] [PubMed]
- Straarup, E.M.; Fisker, N.; Hedtjarn, M.; Lindholm, M.W.; Rosenbohm, C.; Aarup, V.; Hansen, H.F.; Orum, H.; Hansen, J.B.; Koch, T. Short locked nucleic acid antisense oligonucleotides potently reduce apolipoprotein B mRNA and serum cholesterol in mice and non-human primates. Nucleic Acids Res. 2010, 38, 7100–7111. [Google Scholar] [CrossRef]
- Takaya, T.; Nihashi, Y.; Kojima, S.; Ono, T.; Kagami, H. Autonomous xenogenic cell fusion of murine and chick skeletal muscle myoblasts. Anim. Sci. J. 2017, 88, 1880–1885. [Google Scholar] [CrossRef]
- Nihashi, Y.; Ono, T.; Kagami, H.; Takaya, T. Toll-like receptor ligand-dependent inflammatory responses in chick skeletal muscle myoblasts. Dev. Comp. Immunol. 2019, 91, 115–122. [Google Scholar] [CrossRef]
- Nihashi, Y.; Umezawa, K.; Shinji, S.; Hamaguchi, Y.; Kobayashi, H.; Kono, T.; Ono, T.; Kagami, H.; Takaya, T. Distinct cell proliferation, myogenic differentiation, and gene expression in skeletal muscle myoblasts of layer and broiler chickens. Sci. Rep. 2019, 9, 16527. [Google Scholar] [CrossRef]
- Takaya, T.; Nihashi, Y.; Ono, T.; Kagami, H. Transcription of endogenous retrovirus group K members and their neighboring genes in chicken skeletal muscle myoblasts. J. Poult. Sci. 2021, 58, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Macke, T.J.; Case, D.A. Modeling unusual nucleic acid structures. In Molecular Modeling of Nucleic Acids; Leontis, N.B., SantaLucia, J., Eds.; American Chemical Society: Washington, DC, USA, 1998; pp. 379–393. [Google Scholar]
- Galindo-Murillo, R.; Robertson, J.C.; Zgarbova, M.; Sponer, J.; Otyepka, M.; Jurecka, P.; Cheatham, T.E., 3rd. Assessing the current state of Amber force field modifications for DNA. J. Chem. Theory. Comput. 2016, 12, 4114–4127. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Ikebe, J.; Umezawa, K.; Kamiya, N.; Sugihara, T.; Yonezawa, Y.; Takano, Y.; Nakamura, H.; Higo, J. Theory for trivial trajectory parallelization of multicanonical molecular dynamics and application to a polypeptide in water. J. Comput. Chem. 2011, 32, 1286–1297. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.Y.; Berryman, J.T.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., 3rd; Cisneros, G.A.; Cruzeiro, V.W.D.; et al. Amber 2023; University of California: San Francisco, CA, USA, 2023. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupi, C.; Ferraroni, M.; Billa, A.R.; Scheggi, F.; Gratteri, P. The crystal structure of human telomeric DNA complexed with berberine: An interesting case of stacked ligand to G-tetrad ratio higher than 1:1. Nucleic Acids Res. 2013, 41, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, J.; Nomura, Y.; Tanaka, Y.; Torigoe, H.; Nakamura, Y.; Sakamoto, T.; Kozu, T. A G-quadruplex-forming RNA ap-tamer binds to the MTG8 TAFH domain and dissociates the leukemic AML1-MTG8 fusion protein from DNA. FEBS Lett. 2020, 594, 3477–3489. [Google Scholar] [CrossRef] [PubMed]
- Awadasseid, A.; Ma, X.; Wu, Y.; Zhang, W. G-quadruplex stabilization via small-molecules as a potential anti-cancer strategy. Biomed. Pharmacother. 2021, 139, 111550. [Google Scholar] [CrossRef] [PubMed]
- Guedin, A.; Gros, J.; Alberti, P.; Mergny, J.L. How long is too long? Effects of loop size on G-quadruplex stability. Nucleic Acids Res. 2010, 38, 7858–7868. [Google Scholar] [CrossRef]
- Zavyalova, E.; Golovin, A.; Timoshenko, T.; Babiy, A.; Pavlova, G.; Kopylov, A. DNA aptamers for human thrombin with high anticoagulant activity demonstrate target- and species-specificity. Curr. Med. Chem. 2012, 19, 5232–5237. [Google Scholar] [CrossRef] [PubMed]
- Lakhin, A.V.; Tarantul, V.Z.; Gening, L.V. Aptamers: Problems, solutions and prospects. Acta Naturae 2013, 5, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef]
- Chen, J.; Guo, K.; Kastan, M.B. Interactions of nucleolin and ribosomal protein L26 (RPL26) in translational control of human p53 mRNA. J. Biol. Chem. 2012, 287, 16467–16476. [Google Scholar] [CrossRef]
- Cerone, M.A.; Marchetti, A.; Bossi, G.; Blandino, G.; Sacchi, A.; Soddu, S. p53 is involved in the differentiation but not in the differentiation-associated apoptosis of myoblasts. Cell Death Differ. 2000, 7, 506–508. [Google Scholar] [CrossRef]
- Porrello, A.; Cerone, M.A.; Coen, S.; Gurtner, A.; Fontemaggi, G.; Cimino, L.; Piaggio, G.; Sacchi, A.; Soddu, S. p53 regulates myogenesis by triggering the differentiation activity of pRb. J. Cell Biol. 2000, 151, 1295–1304. [Google Scholar] [CrossRef]
- Harford, T.J.; Kliment, G.; Shukla, G.C.; Weyman, C.M. The muscle regulatory transcription factor MyoD participates with p53 to directly increase the expression of the pro-apoptotic Bcl2 family member PUMA. Apoptosis 2017, 22, 1532–1542. [Google Scholar] [CrossRef]
- Abdelmohsen, K.; Gorospe, M. RNA-binding protein nucleolin in disease. RNA Biol. 2012, 9, 799–808. [Google Scholar] [CrossRef]
- Tang, Z.; Qiu, H.; Luo, L.; Liu, N.; Zhong, J.; Kang, K.; Gou, D. miR-34b modulates skeletal muscle cell proliferation and differentiation. J. Cell. Biol. 2017, 118, 4285–4295. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Jang, G.H.; Kang, H.Y.; Song, G. Predicting aptamer sequences that interact with target proteins using an aptamer-protein interaction classifier and a Monte Carlo tree search approach. PLoS ONE 2021, 16, e0253760. [Google Scholar] [CrossRef]
- Molina, A.G.; Sanghvi, Y.S. Liquid-phase oligonucleotide synthesis: Past, present, and future predictions. Curr. Protoc. Nucleic Acid Chem. 2019, 77, e82. [Google Scholar] [CrossRef] [PubMed]
- Sacca, B.; Lacroix, L.; Mergny, J.L. The effect of chemical modifications on the thermal stability of different G-quadruplex-forming oligonucleotides. Nucleic Acids Res. 2005, 33, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Hwang, A.B.; Brack, A.S. Muscle stem cells and aging. Curr. Top. Dev. Biol. 2018, 126, 299–322. [Google Scholar] [PubMed]
- McCormick, R.; Vasilaki, A. Age-related changes in skeletal muscle: Changes to life-style as a therapy. Biogerontology 2018, 19, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Rolland, Y.; Dray, C.; Vellas, B.; Barreto, P.S. Current and investigational medications for the treatment of sarcopenia. Metabolism 2023, 149, 155597. [Google Scholar] [CrossRef] [PubMed]
- Domaniku, A.; Bilgic, S.N.; Kir, S. Muscle wasting: Emerging pathways and potential drug targets. Trends Pharmacol. Sci. 2023, 44, 705–718. [Google Scholar] [CrossRef]
- Golan, T.; Geva, R.; Richards, D.; Madhusudan, S.; Lin, B.K.; Wang, H.T.; Walgren, R.A.; Stemmer, S.M. LY2495655, an antimyostatin antibody, in pancreatic cancer: A randomized, phase 2 trial. J. Cachexia Sarcopenia Muscle 2018, 9, 871–879. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Nucleotide | Sequence (5′-3′) | Reference |
|---|---|---|---|
| iMyo01 | PS-ODN | TTGGGTGGGGAA | This study |
| iMyo02 | PS-ODN | TTTGGGTGGGGAAA | This study |
| iMyo03 | PS-ODN | TTGGGTGAGGGGAA | This study |
| iMyo04 | PS-ODN | TTTGGGTGAGGGGAAA | This study |
| iMyo05 | PS-ODN | TTGGGTGGGAA | This study |
| iMyo06 | PS-ODN | TTTGGGTGGGAAA | This study |
| iMyo07 | PS-ODN | TTGGGTGAGGGAA | This study |
| iMyo08 | PS-ODN | TTTGGGTGAGGGAAA | This study |
| iSN04 | PS-ODN | AGATTAGGGTGAGGGTGA | [7] |
| iSN14 | PS-ODN | ACATTAGGGTGAAAATGA | [7] |
| iSN45 | PS-ODN | GATCCTCAAGCTTAGGTC | [7] |
| iMyo01-DNA | DNA | TTGGGTGGGGAA | This study |
| iMyo01-CS | DNA | TTCCCCACCCAA | This study |
| 12nt-Ctrl | DNA | GCATTGGTATTC | [17] |
| 12nt-Ctrl-CS | RNA | GAAUACCAAUGC | This study |
| RBD | Human vs. Mouse | Human vs. Chicken | Mouse vs. Chicken |
|---|---|---|---|
| RBD1 | 85.9% | 56.4% | 56.4% |
| RBD2 | 83.8% | 69.3% | 64.5% |
| RBD3 | 89.3% | 71.1% | 67.1% |
| RBD4 | 93.7% | 88.2% | 89.5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umezawa, K.; Ikeda, R.; Sakamoto, T.; Enomoto, Y.; Nihashi, Y.; Shinji, S.; Shimosato, T.; Kagami, H.; Takaya, T. Development of the 12-Base Short Dimeric Myogenetic Oligodeoxynucleotide That Induces Myogenic Differentiation. BioTech 2024, 13, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech13020011
Umezawa K, Ikeda R, Sakamoto T, Enomoto Y, Nihashi Y, Shinji S, Shimosato T, Kagami H, Takaya T. Development of the 12-Base Short Dimeric Myogenetic Oligodeoxynucleotide That Induces Myogenic Differentiation. BioTech. 2024; 13(2):11. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech13020011
Chicago/Turabian StyleUmezawa, Koji, Rena Ikeda, Taiichi Sakamoto, Yuya Enomoto, Yuma Nihashi, Sayaka Shinji, Takeshi Shimosato, Hiroshi Kagami, and Tomohide Takaya. 2024. "Development of the 12-Base Short Dimeric Myogenetic Oligodeoxynucleotide That Induces Myogenic Differentiation" BioTech 13, no. 2: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/biotech13020011