
IL17A Suppresses IGFBP1 in Human Endometrial Stromal Cells
by
 , and
, and
Susumu Tanaka
1,* ,
,
Misa Sawachika
1,
Namika Yoshida
1,
Kensuke Futani
1,
Hiromi Murata
2 and
Hidetaka Okada
2 1
Department of Nutrition Science, University of Nagasaki, Siebold, Nagasaki 851-2195, Japan
2
Department of Obstetrics and Gynecology, Kansai Medical University, 2-5-1 Shin-Machi, Hirakata 573-1010, Japan
*
Author to whom correspondence should be addressed.
Reprod. Med. 2024, 5(2), 43-56; https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed5020006
Submission received: 6 March 2024
/
Revised: 22 April 2024
/
Accepted: 24 April 2024
/
Published: 26 April 2024
Abstract
:Interleukin (IL) 17A has been implicated in preeclampsia, preterm labor, and miscarriage. IL17A production in non-lymphoid tissues is mainly carried out by unconventional γδ17T cells. Innate lymphoid cells (ILCs) 3, a subgroup of innate lymphocytes, can also be a source of IL17A in the endometrium and are required from implantation to early pregnancy, with their regulation ensuring that pregnancy continues. Herein, we examined the expression of γδ17T cells and ILC3 regulators IL1B, IL23A, and IL17D and IL17A receptors (IL17RA/IL17RC) in human endometrial stromal cells (EnSCs) and cell lines (KC02-44D). Accordingly, quantitative polymerase chain reaction and immunoblotting were employed. IL1B, IL23A, and IL17D were significantly upregulated in decidualized EnSCs and KC02-44D cells. A significant augmentation in IL17RA/IL17RC was also observed in decidualization. IL17A stimulation of KC02-44D cells during decidualization suppressed the decidualization marker IGFBP1. The involvement of transcription factor Forkhead box protein O1 (FOXO1) in this repression was reflected by its translocation from the nucleus into the cytoplasm. A role for IkB kinase alpha in FOXO1 phosphorylation-mediated migration was also suggested. Taken together, our findings indicate that the secretion of IL17A by γδ17T and ILC3 cells in the uterus contributes to EnSCs function and may play critical roles in regulating IGFBP1-mediated implantation and fetal growth.
1. Introduction
The human menstrual cycle takes approximately 28 days, during which the proliferation of the functional layer, secretory changes in epithelial cells, stromal cell decidualization, and menstruation occur periodically within the endometrium [1]. After the follicle transforms into the corpus luteum, the uterine gland secretion is activated by progesterone, which is produced by the corpus luteum. Progesterone also acts on endometrial stromal cells (EnSCs) to induce morphological and functional differentiation, known as decidualization, thus providing a microenvironment suitable for implantation of the embryo in humans. Upon decidualization, EnSCs produce and secrete cytokines, chemokines, and growth factors, such as insulin-like growth factor binding protein 1 (IGFBP1), angiogenic factors, and prolactin (PRL) [2]. Abnormal decidualization leads to placental dysplasia and underdevelopment of the spiral artery, which can cause subsequent implantation failure and miscarriage [2,3,4].
In early pregnancy, the embryonic and maternal cells have been found to interact [5]. The prediction of receptor–ligand molecular pairs in the placenta and decidua of women at 6–14 weeks of gestation indicated the presence of pregnancy-specific cell subsets and revealed that EnSCs and immune cells in the decidua interact with embryo-derived extravillous trophoblasts. Data suggest that the inflammatory cytokine interleukin (IL)17A may have a pathological role in preeclampsia, premature birth, and miscarriage, with previous studies having focused on IL17A produced by immune cells at the maternal–fetal interface [6,7,8]. IL17A is part of the IL17 family, which plays important roles in immune responses and host defense [9,10]. Interestingly, members of this family have been proposed to have unique and non-overlapping functions against cancer, autoimmune disease, and infection. IL17A, the first member of this family to be identified, plays a critical role during infection. Persistently high levels of IL17A and homologous proteinIL17F induce inflammation. Meanwhile, IL17B has been implicated in tumorigenesis [11]. IL17C influences mucosal barrier integrity and is implicated in autoimmune diseases [12,13]. IL-17D is a newly identified cytokine, whose receptor was recently identified as CD93 [14,15]. IL-17E (IL-25) enhances allergic inflammatory responses and is involved in host defense [16,17,18]. A dysregulation of IL17A has been reported in unexplained recurrent pregnancy loss [19,20,21,22]. There are emerging reports that endometrial IL17A promotes trophoblast migration [23,24].
Originally, IL17A was thought to be produced mainly by Th17 cells, with a subsequent report showing that IL17A-secreting Th17 cells induced human trophocyte invasion in first-trimester trophocytes [25]. However, γδ17T cells have recently attracted attention as an alternative source of IL17A in non-lymphoid tissues, including the uterus [26,27,28]. Murine uterine γδT cells produce high levels of IL17A; therefore, they are called γδ17T cells [23]. Further, the reduction of Th17 cell reactivity and reversal of the Th17/Treg imbalance did not improve pregnancy outcomes in recurrent implantation failure, suggesting that Th17 cell-derived IL17A exerts minimal effects in the endometrium. Innate lymphoid cells (ILCs) 3 are another potential source of IL17A within the endometrium, as they express RORɤt [29,30,31,32]. Although ILC3 almost disappear from the decidua in late pregnancy [30,33], an increase in ILC3 numbers in the decidua causes early delivery [34], suggesting the need for ILC3 production between implantation and early pregnancy as well as its suppression during continued pregnancy. Decreased ILC production or abnormal functional interactions of ILCs with the human decidua may cause fetal death [35]. Despite a marked increase in ILC3 within the endometrium during the implantation window being associated with autoimmune thyroid disease and female infertility, the function of ILC3 has been reported to be suppressed in that period [36]. Thus, we hypothesized that γδ17T cells and ILC3 may be regulated by endometrium-derived cells, as well as other immune cells, including uterine natural killer (uNK) cells specifically found in the endometrium [33,37].
In this study, we observed for the first time the upregulation of IL1B, IL23A (which enhances the production of IL17A by γδ17T [38,39,40] and ILC3 [41]), and IL17D (which specifically regulates ILC3 function via the CD93 receptor [14,15]) in decidualized EnSCs. Based on the increase in IL17A receptor expression during decidualization, we investigated whether IL17A secreted by activated-γδ17T and ILC3 could affect decidualization through a feedback mechanism. IL17A downregulated decidualization marker IGFBP1 in decidualized EnSCs and the KC02-44D cell line and regulatory mechanisms for IGFBP1 downregulation by Forkhead box protein O1 (FOXO1) phosphorylation.
2. Materials and Methods
2.1. Ethical Statement
The study was explained to all eligible patients, and we obtained informed consent from all participants. The Kansai Medical University review board approved this study (ID: 2006101), and it was conducted in accordance with the Helsinki Declaration. Human uteri with a benign myoma were donated by six patients (42–50 years) with regular menstrual cycles (Table 1). Patients who received preoperative hormone therapies were not included in this study. Histologically normal endometria were obtained from all subjects.
2.2. Decidualization of Human EnSCs
Human EnSCs were purified from the endometrium through a previously described method [42]. For EnSCs culture, DMEM-F12 medium without phenol red was purchased from Thermo Fisher Scientific (Waltham, MA, USA) and supplemented with Glutamax (2 mmol/L, Thermo Fisher Scientific), 10% Charcoal Stripped-Fetal Bovine Serum(CS-FBS; Biowest, Nuaillé, France), streptomycin (100 μg/mL), and penicillin (100 IU/mL) (DMEM-F12/CS-FBS medium). The cells were then cultured in a 37 °C humidified environment with 5% CO2 until near confluence, changing the medium every 3 days. EnSCs were seeded anew, and grown until confluence prior to experiments. EnSCs were then stimulated with 10−8 mol/L estradiol (E2) and 10−7 mol/L medroxyprogesterone acetate (MPA) for up to 2 weeks for induction of decidualization [43,44]. Unstimulated cells were prepared as a control group.
2.3. Human EnSC Cell Line KC02-44D and Treatment
Because patient-derived EnSCs can vary greatly in their responsiveness to stimulation, owing to the influence of many confounding factors, including differences in the timing of collection during the menstrual cycle, the expression of cytokines that may regulate IL17A-producing cells was examined using KC02-44D cells (American Type Culture Collection, Manassas, VA, USA), an established human EnSCs cell line [45]. KC02-44D were cultured in DMEM medium without phenol red containing Glutamax (2 mmol/L, Thermo Fisher Scientific), 10% Fetal Bovine Serum (EU Origin), Charcoal Stripped (CS-FBS, Biowest, France), streptomycin (100 μg/mL), and penicillin (100 IU/mL). Cells were cultured until near confluence, changing the medium every 3 days. KC02-44D were treated with 10−8 mol/L E2, 10−7 mol/L MPA, and 0.5 mM 8-Bromo-cAMP (Sigma-Aldrich Co., LLC, St. Louis, CO, USA) (E2 + MPA + cAMP treatment) for up to 6 days for the induction of decidualization [46]. Unstimulated cells were prepared as a control group. To test the effects of IL17A on decidualization, in addition to the control and E2 + MPA + cAMP treatments, 10 ng/mL IL17A (Recombinant Human IL-17A, Fujifilm Corp., Tokyo, Japan) or E2 + MPA + cAMP + 10 ng/mL IL17A stimulations were conducted for up to 6 days.
2.4. Quantitative Polymerase Chain Reaction (qPCR)
The extraction of total RNA from EnSCs or KC02-44D cells with or without treatment was conducted using the Sepasol®-RNA I Super G (Nacalai Tesque Inc., Kyoto, Japan). Reverse transcription was performed using a ReverTraAce qPCR RT master mix with gDNA remover (Toyobo, Osaka, Japan). qPCR analysis was conducted on a LightCycler96 (Roche Diagnostics K.K., Tokyo, Japan) and Thunderbird Next qPCR Mix (Toyobo). The qPCR primers are presented in Table 2. Relative expression was calculated using the 2−ΔΔCt method [47]. Hypoxanthine-phospho-ribosyl-transferase 1 (HPRT1) was used as a housekeeping gene.
2.5. Western Blotting
To determine whether IL17A stimulation led to FOXO1 phosphorylation, we examined the downstream pathways of IL17A. IL17A binds to the IL17RA homodimer, IL17RC homodimer, and/or IL17RA-IL17RC heterodimer [48] and activates intracellular ACT1 by changing its steric structure, subsequently transmitting the signal to IkB kinase alpha (IKKA)/IKKB [49]. In contrast, the downstream regulation of Akt and FOXO1 phosphorylation by IKKA has been previously described [50].
The soluble fraction was prepared from KC02-44D cells cultured with or without stimulation using MPER reagent (Thermo Fisher Scientific) and protease inhibitors (Roche Diagnostics K.K.). Western blotting was conducted to quantify IKKA and IKKB protein levels. Proteins were separated on a 10% sodium dodecyl sulfate (SDS)–polyacrylamide gel and transferred onto a PVDF membrane, which was then blocked using Blocking One (Nacalai Tesque Inc.). The membrane was then incubated with 1/1000 IKKβ (D30C6) rabbit mAb (#8943, Cell Signaling Tech, Danvers, MA, USA, RRID: AB_11024092), 1/1000 IKKα (D3W6N) rabbit mAb (#61294, Cell Signaling Tech, RRID: AB_2799606), or 1/10,000 mouse β-actin antibody (Sigma-Aldrich, RRID: AB_476743, Cat# A5316) in TBS containing 5% Blocking One and 0.1% Tween-20 overnight at 4 °C. After washing, the membrane was incubated with 1:5000 Goat anti-rabbit-HRP (RRID: AB_2336198, VECTOR Laboratories, Burlingame, CA, USA, Cat# PI-1000) or 1:10,000 sheep anti-mouse-HRP (RRID: AB_772210, GE Healthcare Life Science, Chicago, IL, USA, Cat# NA931). The resulting complexes were visualized using Chemi-Lumi One L (Nacalai Tesque Inc.) and LAS 4000 (GE Healthcare Life Science). ImageJ 1.54g software (National Institutes of Health, Maryland, MD, USA) was used to determine band intensity as previously described [51]. ACTB was used as a loading control.
2.6. Immunocytochemistry
The sterilized cover glass was laid on a 6-well plate, and 0.3 × 10⁶ KC02-44D cells were seeded onto it. Three groups were studied: (I) control, (II) E2 + MPA + cAMP-treated, and (III) E2 + MPA + cAMP + 10 ng/mL IL17A-treated groups. After 6 days with medium change every 3 days, cultured KC02-44D were subjected to formaldehyde fixation for 5 min at 22 °C and were then permeabilized with methanol for 5 min at 4 °C. KC02-44D were then incubated with a primary antibody (Foxo1-rabbit-mAb (C29H4) #2880 diluted to 1/1000, Cell Signaling Tech, RRID: AB_2106495) overnight at 4 °C, followed by incubation with a goat anti-rabbit-Alexa488 (# A-11008, Molecular Probes, US-OR, RRID: AB_143165) diluted 1/3000 and 1 µg/mL propidium iodide. LSM700 (Carl Zeiss Co., Ltd., Tokyo, Japan) was used to observe the stained cells. Positive staining rates in the nucleus and cytoplasm were calculated and averaged by visually counting the cytoplasm-stained, nucleus-localized, and cytoplasm-localized FOXO1 in images obtained from two independent individuals. In total, 15 images (945 cells) were taken in group I, 15 images (552 cells) in group II, and 15 images (552 cells) in group III of an area covering 1 mm2. These images were used for analysis.
2.7. Statistical Analyses
The normality of data was confirmed using the Shapiro–Wilk test. Welch’s t-test was used to compare the means of the two groups with the Bonferroni correction. p-values of < 0.05 were considered as indicative of statistical significance. SPSS software was used for all statistical analyses (version 22.0; IBM Corp., Armonk, NY, USA).
3. Results
3.1. Elevations in IL1B, IL23A, and IL17D in Decidualized EnSCs
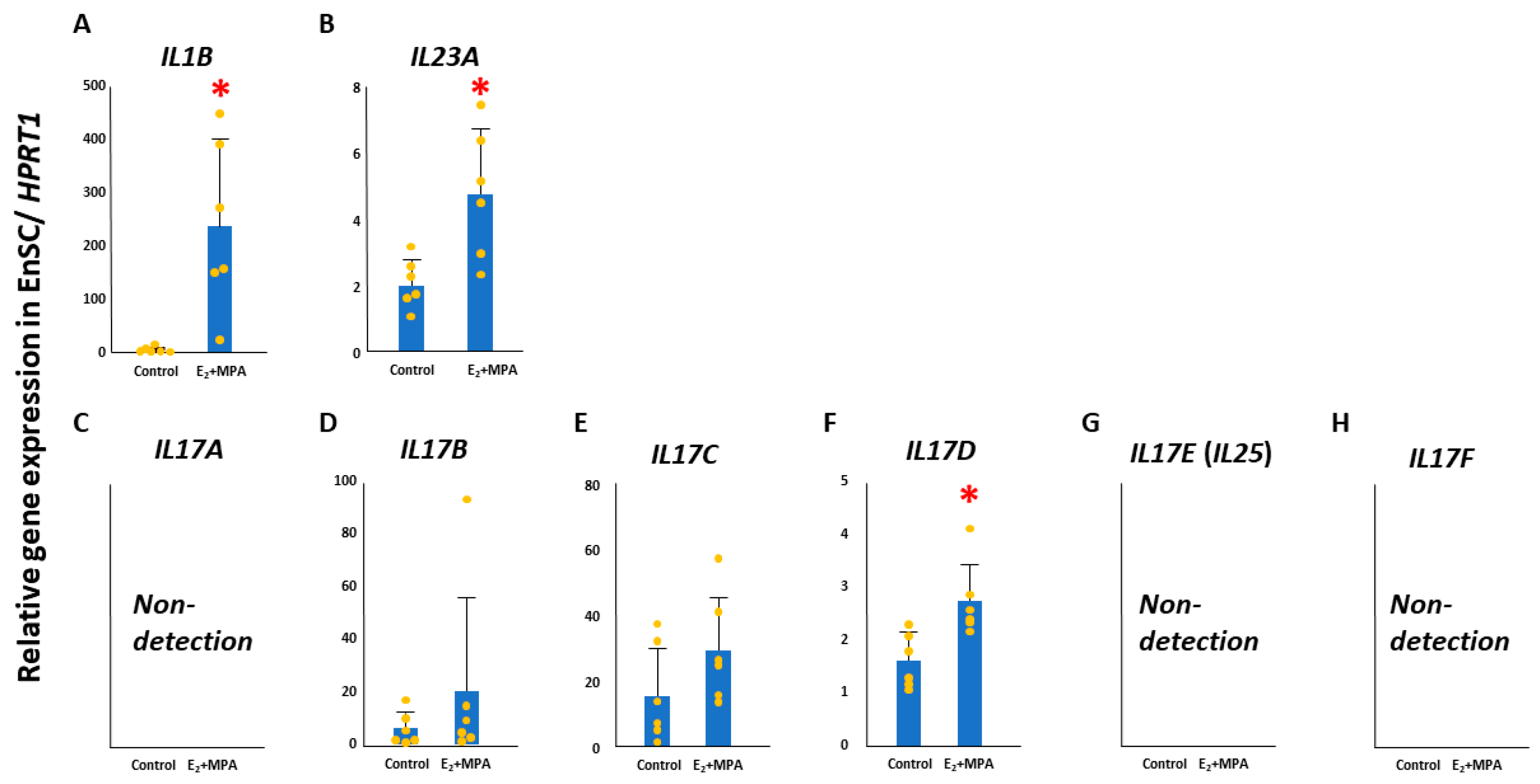
After 12 days of decidualization treatment of primary culture EnSCs, IL1B and IL23A were significantly upregulated compared with their levels in unstimulated cells (control) (p < 0.05) (Figure 1). Further examination of changes in the expression of IL17 gene family members revealed no expression of IL17A, IL17E (IL25), or IL17F in EnSCs. In contrast, IL17B, IL17C, and IL17D were detected, and a significant upregulation of IL17D was observed during decidualization (p < 0.05, Figure 1).
3.2. Cytokines in KC02-44D Cells
Significant upregulation of IL1B and IL23A was shown in decidualized KC02-44D compared with their levels in control cells (p < 0.05, Figure 2). Meanwhile, there was no change in IL17A, IL17B, IL17C, IL17E, or IL17F (Figure 2). IL17D, however, was significantly upregulated in KC02-44D cells during decidualization (p < 0.05; Figure 2).
3.3. Responsiveness of EnSCs to γδ17T and ILC3-Derived Cytokines
Activated γδ17T and ILC3 cells secrete IL17A and IL22. IL17A receptor-encoding genes IL17RA and IL17RC [48] were significantly upregulated in decidualized EnSCs (Figure 3, p < 0.05). Meanwhile, there was no expression of IL22RA1, which encodes an IL22 receptor, in the EnSCs (Figure 3). Receptor gene expression was confirmed in human endometrial tissue.
Subsequently, KC02-44D cells were subjected to a 6-day decidualization treatment to examine changes in interleukin receptor expression. qPCR analysis confirmed a significant upregulation of IL17RA and IL17RC in decidualized KC02-44D (p < 0.05). IL22RA1 expression was not detected (Figure 3).
3.4. IL17A Treatment of KC02-44D Cells
As IL17RA and IL17RC expression was detected in EnSCs, IL17A secreted from activated γδ17T and ILC3 cells might have some effect on decidualization. Therefore, we treated KC02-44D cells for 6 days with 0, 0.1, 1.0, 10, or 100 ng/mL IL17A, based on previous studies [37] to examine changes in PRL. A significant increase in PRL was observed in the decidualized group relative to the control group. However, no effect of IL17A was observed. Because there was no clear difference in the effect of IL17A concentration on PRL, we examined its effect on other genes using concentrations from previous studies (10 ng/mL) [52].
3.5. Effect of IL17A on Decidualization Markers in KC02-44D Cells
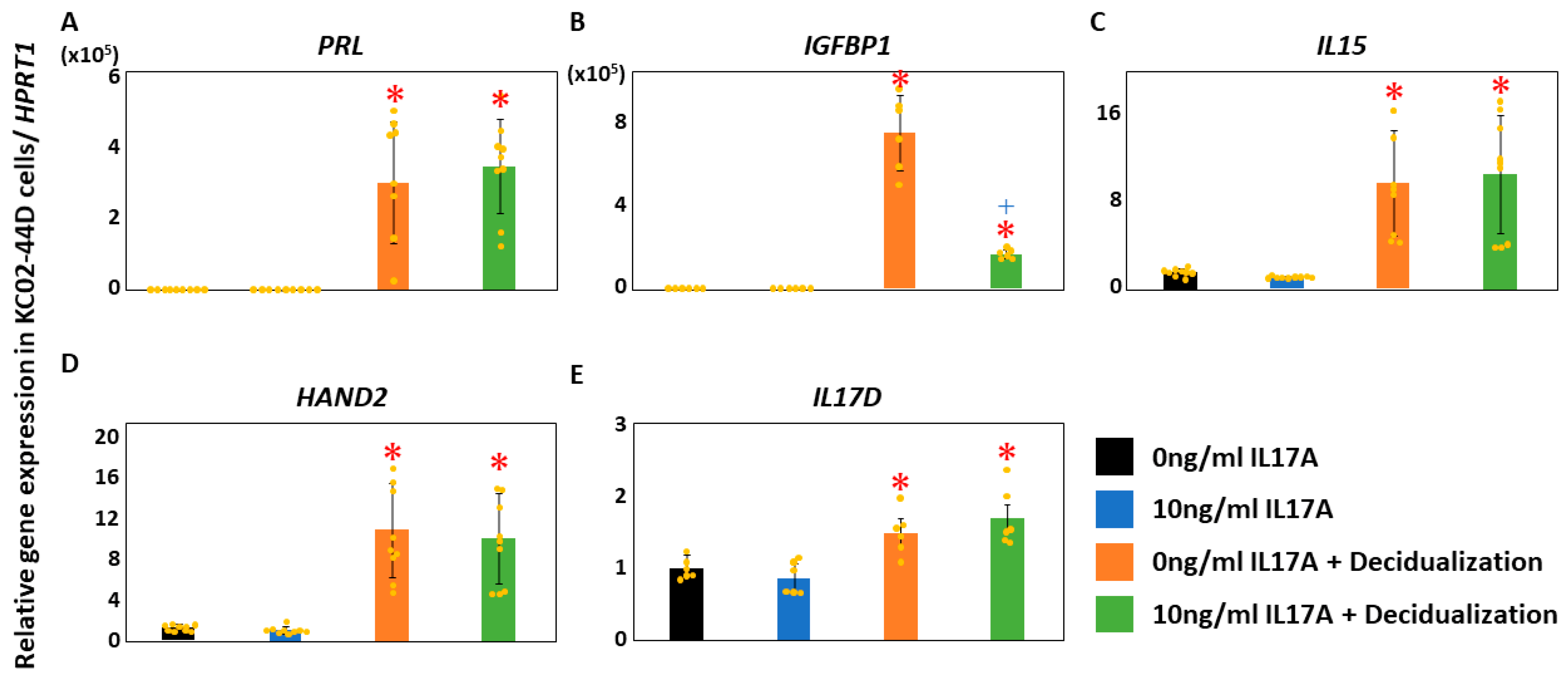
Next, we examined the decidualization markers [53] in decidualized KC02-44D cells treated with 10 ng/mL IL17A. Although IGFBP1 was significantly upregulated in decidualized KC02-44D (p < 0.05), significant suppression of IGFBP1 was observed in the IL17A-stimulated cells compared to levels in decidualized KC02-44D (p < 0.05, Figure 4). Further, significant upregulation of PRL, IL15, HAND2, and IL17D was noted in the decidualized KC02-44D (p < 0.05, Figure 4). There was no effect of additional IL17A stimulation in E2 + MPA+ cAMP-treated cells (Figure 4).
3.6. Effect of IL17A on FOXO1 Localization in KC02-44D Cells
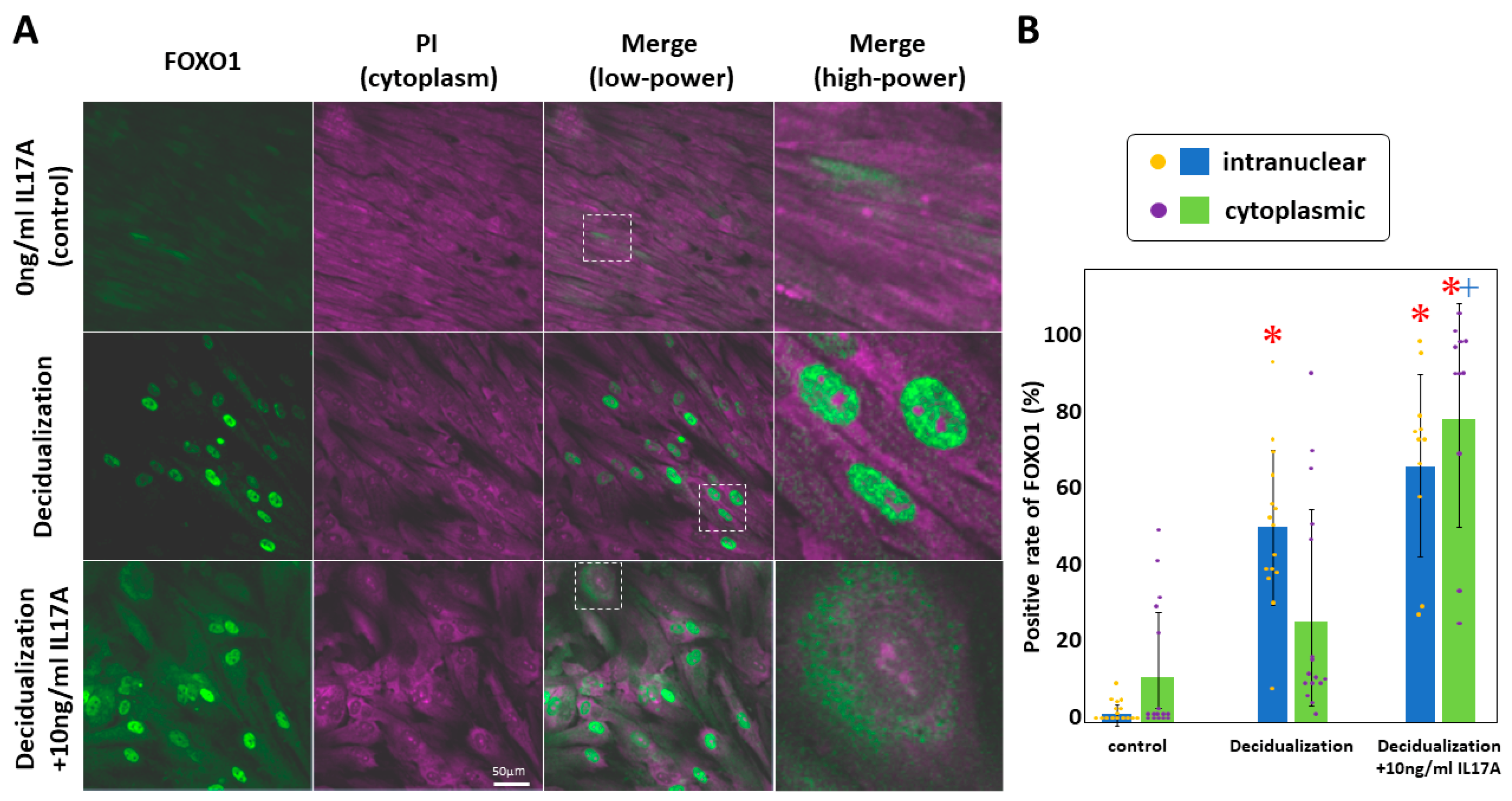
In KC02-44D cells, IL17A stimulation causes the phosphorylation of decidualization marker FOXO1 and its subsequent migration out of the nucleus, resulting in a downregulation of IGFBP1 [54,55]. After KC02-44D cells were stimulated with or without IL17A for 6 days, we examined the subcellular localization of FOXO1 [56]. We expected to stain the nucleus with propidium iodide, but the cytoplasm and nucleolus were also fluorescently labeled (Figure 5, purple). This localization suggested that the RNA molecules were labeled with propidium iodide. FOXO1 was fluorescently labeled with a FOXO1 antibody and Alexa488-secondary antibody (green). Increased localization of FOXO1 in the nucleus was found in the decidualized cells in comparison with that in the untreated cells, with changes in FOXO1 localization to the cytoplasm observed following additional IL17A stimulation (Figure 5A). FOXO1 localization in the nucleus or cytoplasm was determined, and the frequency of FOXO1 relative to the total cell number included in the entire image was calculated for each image, whereafter differences in frequency were assessed. A significant increase in the nuclear localization of FOXO1 was found in decidualized cells and IL17A additionally stimulated cells in comparison with that in untreated KC02-44D controls (p < 0.05). Furthermore, a significant increase in FOXO1 translocation from the nucleus to the cytoplasm was observed in the IL17A additionally stimulated cells in comparison to that in decidualized cells (p < 0.05) (Figure 5B).
3.7. Mechanism of IL17A-Dependent FOXO1 Migration
To assess changes in the downstream signaling of IL17A, we detected IKKA and IKKB levels in KC02-44D cells after additional stimulation with IL17A. Normalized IKKA band densities were 2.59 ± 0.03 (mean ± SD, control), 1.09 ± 0.46 (decidualization), and 0.75 ± 0.14 (decidualization +IL17A) (Figure 6A). IKKB molecular densities were 0.13 ± 0.03 (control), 0.16 ± 0.02 (decidualization), and 0.16 ± 0.04 (decidualization +IL17A) (Figure 6A). We observed a significant decrease in IKKA (p < 0.05) and no change in IKKB in the IL17A-stimulated cells compared to control levels (Figure 6B).
4. Discussion
In this study, decidualization of EnSCs isolated from the human endometrium and of the endometrial stromal cell line KC02-44D induced a significant upregulation of IL1B, IL23A, and IL17D expression [15,41], affecting the proliferation, differentiation, and functional regulation of γδ17T and ILC3 [30,33,38,39,40,57,58], which is involved in innate immunity and embryo implantation in the endometrium. This finding prompted us to explore the interactions between decidualized EnSCs and IL17A-producing immune cells. Therefore, we examined the expression of IL17RA/IL17RC and IL22RA1, which encode receptors for IL17A and IL22 [32] secreted from activated γδ17T and ILC3, observing the upregulation of IL17RA and IL17RC, but not IL22RA1, in both EnSCs and KC02-44D cells. Furthermore, IL17A stimulation during decidualization of KC02-44D cells suppressed IGFBP1 expression. As the involvement of FOXO1 in this repression has been suggested [59], the intracellular localization of FOXO1 was examined. FOXO1 migration into the cytoplasm was observed in decidualized cells with additional IL17A stimulation. IGFBP1 downregulation occurred following FOXO1 migration out of the nucleus. Furthermore, analysis of IL17A receptor downstream signaling revealed a reduction in the amount of IKKA, suggesting the involvement of IKKA in this localization change.
It has been suggested that γδT cells are the major source of IL17A in various non-lymphoid tissues, including the uterus [23,26,27,28]. The crucial role of IL17A in maintaining health in response to infection, physiological stress, and injury, as well as in autoimmune diseases, is well recognized [38,60,61,62]. γδT cells, which are thought to be a key source of IL17A in the uterus [23], may facilitate IL17A-induced trophoblast migration and thus play an important role in defense against infection, in addition to potential involvement in pregnancy in young women. Unlike γδT cells in other tissues, uterine immune cells need to both defend against infection and tolerate allogeneic fetuses during pregnancy. γδ17T cells in the endometrium do not express granzyme B and CD107a like cytotoxic γδT [23]. Thus, γδ17T cells are thought to tolerate the fetus, with cytotoxic γδT providing protection against infection. Comparison of data from allogenic and syngeneic reproductive pairs also indicates that γδT cells may play an important role in successful pregnancy [23]. Furthermore, γδT cells have been found to be abundant at the maternal–fetal interface during pregnancy as well as in the non-pregnant uterus [23], suggesting their involvement in endometrial decidualization. In the decidualized EnSCs in this study, IL17A was implicated in the regulation of IGFBP1 expression, a major decidualization marker. Overexpression of IGFBP1 in the endometrium has been associated with impaired fetal growth and placental insufficiency and is thought to cause placenta accreta [63]. Our results suggest that in decidualized EnSCs, IL17A decreases IKKA protein levels, regulates downstream Akt and FOXO1 phosphorylation, and restores IGFBP1 expression to a constant level. This suggests that IL17A secreted by γδ17T and ILC3 may contribute to the prevention of fetal growth defects caused by placental insufficiency and the development of placenta accreta by restricting the overexpression of IGFBP1 in decidualized EnSCs.
ILCs are immune cells involved in innate defense and tissue remodeling. NK cells, lymphoid tissue inducer cells, and ILC1-3 belong to the ILC family [64], with all subsets observed within human uterine tissues [29,30,31,33]. The ILC3 fraction identified via flow cytometry also contains ILC precursors that can give rise to ILC1-3 [65,66]. ILC3 in the human decidua interact with decidualized EnSCs, suggesting that they contribute to innate immunity as well as vascular and tissue architecture [33]. Indeed, ILC3 in the decidua likely act on extravillous trophoblasts and modulate invasion by secreting granulocyte–macrophage colony-stimulating factor, X-C motif chemokine ligand 1, and macrophage inflammation protein 1α and 1β [31,67,68]. The results of this study suggest that their secretion from ILC3 [68] is dependent on activation by IL1B, IL23A, and IL17D secreted from decidualized EnSCs.
IL17A, from intrauterine γδ17T cells, which is thought to play an important role in EVT invasion, is highly abundant [23]. γδ17T cells are recruited into the uterus in response to estrogen, which increases in the blood with follicle development and induces IL17A production in γδ17T cells [69]. Therefore, their involvement in uterine function from the proliferative to secretory phases is also possible. In this context, it is interesting to follow the changes in expression and secretion of IL1B, IL23, and IL17D in EnSCs under estrogen stimulation conditions. ILC3 was reduced in eutopic endometrium during endometriosis [70] when compared to levels in normal endometrium. Therefore, we suggest the involvement of ILC3 in normal decidualization and the menstrual cycle. Further, dysregulation of ILC3 might contribute to endometriosis, consequent implantation failure, and recurrent pregnancy loss.
Limitations
This study demonstrated that the levels of IL1B, IL23A, and IL17D, which regulate γδ17T and ILC3 proliferation and differentiation, are elevated in decidualized EnSCs. Further, IL17A secreted from these cells affects EnSCs decidualization. However, because γδ17T and ILC3 samples derived from the human endometrium are difficult to obtain and these cell lines have not been established outside of mice [71,72], examining the effect of supernatant from decidualized human EnSC culture, which is thought to contain IL17D and other factors, on the growth and differentiation of human cells has not been possible. Since IL17D is also expressed in CD19(+) B cells and quiescent CD4(+) T cells [73,74], determining the localization of IL17D expression in EnSCs using actual uterine tissue sections is necessary. Furthermore, a comparison of IL1B, IL23A, and IL17D expression in decidualized EnSCs and the presence of γδ17T or ILC3 in the vicinity of decidualized EnSCs using endometrial tissue sections from healthy subjects and patients with infertility may lead to the discovery of a new category of obstetric diseases caused by γδ17T and/or ILC3. It is also necessary to measure cytokines, including IL17A and others, in the blood of patients experiencing preeclampsia, preterm labor, and miscarriage.
5. Conclusions
We found that EnSCs may regulate γδ17T and ILC3 in the endometrium and that IGFBP1 is suppressed by IL17A secreted by the latter two cell types. This regulation occurs via the IL17A-IKKA-FOXO1 phosphorylation-IGFBP1 axis.
Author Contributions
Conceptualization, S.T.; methodology, S.T., H.M. and H.O.; validation, S.T., M.S., N.Y. and K.F.; formal analysis, S.T., M.S., N.Y. and K.F.; investigation, S.T., M.S., N.Y. and K.F.; resources, S.T., H.M. and H.O.; data curation, S.T. and M.S.; writing—original draft preparation, S.T.; writing—review and editing, S.T., H.M. and H.O.; visualization, S.T. and M.S.; supervision, H.M. and H.O.; project administration, S.T.; funding acquisition, S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Takeda Science Foundation 2018 to Susumu Tanaka; and JSPS KAKENHI to Hidetaka Okada, grant number 21K09480, and Hiromi Murata, grant number 21K09529.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Kansai Medical University (protocol code: 2006101, approval date: 10 July 2006).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Murata, H.; Tanaka, S.; Tsuzuki-Nakao, T.; Kido, T.; Kakita-Kobayashi, M.; Kida, N.; Hisamatsu, Y.; Tsubokura, H.; Hashimoto, Y.; Kitada, M.; et al. The transcription factor hand2 up-regulates transcription of the il15 gene in human endometrial stromal cells. J. Biol. Chem. 2020, 295, 9596–9605. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhao, W.; Huang, X.; Jiang, Z.; Liu, L.; Cui, L.; Li, X.; Li, D.; Du, M. Torc2/3-mediated dusp1 upregulation is essential for human decidualization. Reproduction 2021, 161, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Tanaka, S.; Okada, H. The regulators of human endometrial stromal cell decidualization. Biomolecules 2022, 12, 1275. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.W.; Lai, Z.Z.; Yang, H.L.; Zhou, W.J.; Zhao, X.Y.; Xie, F.; Liu, S.P.; Chen, W.D.; Zhang, T.; Ye, J.F.; et al. An igf1-expressing endometrial stromal cell population is associated with human decidualization. BMC Biol. 2022, 20, 276. [Google Scholar] [CrossRef] [PubMed]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.E.; Stephenson, E.; Polanski, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Chao, F.; Yi, L.; Yin, G.J.; Bao, S.H.; Qiu, L.H.; Lin, Q.D. Increased prevalence of t helper 17 (th17) cells in peripheral blood and decidua in unexplained recurrent spontaneous abortion patients. J. Reprod. Immunol. 2010, 84, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Santner-Nanan, B.; Peek, M.J.; Khanam, R.; Richarts, L.; Zhu, E.; de St Groth, B.F.; Nanan, R. Systemic increase in the ratio between foxp3+ and il-17-producing cd4+ t cells in healthy pregnancy but not in preeclampsia. J. Immunol. 2009, 183, 7023–7030. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Nakashima, A.; Hidaka, T.; Okabe, M.; Bac, N.D.; Ina, S.; Yoneda, S.; Shiozaki, A.; Sumi, S.; Tsuneyama, K.; et al. A role for il-17 in induction of an inflammation at the fetomaternal interface in preterm labour. J. Reprod. Immunol. 2010, 84, 75–85. [Google Scholar] [CrossRef]
- Chang, S.H.; Dong, C. Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal. 2011, 23, 1069–1075. [Google Scholar] [CrossRef]
- Iwakura, Y.; Ishigame, H.; Saijo, S.; Nakae, S. Functional specialization of interleukin-17 family members. Immunity 2011, 34, 149–162. [Google Scholar] [CrossRef]
- Bastid, J.; Dejou, C.; Docquier, A.; Bonnefoy, N. The emerging role of the il-17b/il-17rb pathway in cancer. Front. Immunol. 2020, 11, 718. [Google Scholar] [CrossRef]
- Chang, S.H.; Reynolds, J.M.; Pappu, B.P.; Chen, G.; Martinez, G.J.; Dong, C. Interleukin-17c promotes th17 cell responses and autoimmune disease via interleukin-17 receptore. Immunity 2011, 35, 611–621. [Google Scholar] [CrossRef]
- Reynolds, J.M.; Martinez, G.J.; Nallaparaju, K.C.; Chang, S.H.; Wang, Y.H.; Dong, C. Cutting edge: Regulation of intestinal inflammation and barrier function by il-17c. J. Immunol. 2012, 189, 4226–4230. [Google Scholar] [CrossRef]
- Brembilla, N.C.; Boehncke, W.H. Revisiting the interleukin 17 family of cytokines in psoriasis: Pathogenesis and potential targets for innovative therapies. Front. Immunol. 2023, 14, 1186455. [Google Scholar] [CrossRef]
- Huang, J.; Lee, H.Y.; Zhao, X.; Han, J.; Su, Y.; Sun, Q.; Shao, J.; Ge, J.; Zhao, Y.; Bai, X.; et al. Interleukin-17d regulates group 3 innate lymphoid cell function through its receptor cd93. Immunity 2021, 54, 673–686.e4. [Google Scholar] [CrossRef]
- Barlow, J.L.; McKenzie, A.N. Il-25: A key requirement for the regulation of type-2 immunity. Biofactors 2009, 35, 178–182. [Google Scholar] [CrossRef]
- Morita, H.; Moro, K.; Koyasu, S. Innate lymphoid cells in allergic and nonallergic inflammation. J. Allergy Clin. Immunol. 2016, 138, 1253–1264. [Google Scholar] [CrossRef]
- von Moltke, J.; Ji, M.; Liang, H.E.; Locksley, R.M. Tuft-cell-derived il-25 regulates an intestinal ilc2-epithelial response circuit. Nature 2016, 529, 221–225. [Google Scholar] [CrossRef]
- Wang, W.J.; Zhang, H.; Chen, Z.Q.; Zhang, W.; Liu, X.M.; Fang, J.Y.; Liu, F.J.; Kwak-Kim, J. Endometrial tgf-beta, il-10, il-17 and autophagy are dysregulated in women with recurrent implantation failure with chronic endometritis. Reprod. Biol. Endocrinol. 2019, 17, 2. [Google Scholar] [CrossRef]
- Crosby, D.A.; LGlover, E.; Brennan, E.P.; Kelly, P.; Cormican, P.; Moran, B.; Giangrazi, F.; Downey, P.; Mooney, E.E.; Loftus, B.J.; et al. Dysregulation of the interleukin-17a pathway in endometrial tissue from women with unexplained infertility affects pregnancy outcome following assisted reproductive treatment. Hum. Reprod. 2020, 35, 1875–1888. [Google Scholar] [CrossRef]
- Farshchi, M.; Abdollahi, E.; Saghafi, N.; Hosseini, A.; Fallahi, S.; Rostami, S.; Rostami, P.; Rafatpanah, H.; Habibagahi, M. Evaluation of th17 and treg cytokines in patients with unexplained recurrent pregnancy loss. J. Clin. Transl. Res. 2022, 8, 256–265. [Google Scholar] [PubMed]
- Moura, G.A.; Rocha, Y.M.; Moura, F.L.D.; Freitas, J.O.; Rodrigues, J.P.V.; Goncalves, V.P.; Nicolete, R. Immune system cells modulation in patients with reproductive issues: A systematic review approach. JBRA Assist. Reprod. 2024, 28, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Wu, Q.; Huang, J.; Yang, B.; Liang, C.; Chi, P.; Wu, C. Tissue resident memory gammadeltat cells in murine uterus expressed high levels of il-17 promoting the invasion of trophocytes. Front. Immunol. 2020, 11, 588227. [Google Scholar] [CrossRef]
- You, Y.; Stelzl, P.; Joseph, D.N.; Aldo, P.B.; Maxwell, A.J.; Dekel, N.; Liao, A.; Whirledge, S.; Mor, G. Tnf-alpha regulated endometrial stroma secretome promotes trophoblast invasion. Front. Immunol. 2021, 12, 737401. [Google Scholar] [CrossRef]
- Wu, H.X.; Jin, L.P.; Xu, B.; Liang, S.S.; Li, D.J. Decidual stromal cells recruit th17 cells into decidua to promote proliferation and invasion of human trophoblast cells by secreting il-17. Cell. Mol. Immunol. 2014, 11, 253–262. [Google Scholar] [CrossRef]
- Sumaria; Roediger, B.; Ng, L.G.; Qin, J.; Pinto, R.; Cavanagh, L.L.; Shklovskaya, E.; de St Groth, B.F.; Triccas, J.A.; Weninger, W. Cutaneous immunosurveillance by self-renewing dermal gammadelta t cells. J. Exp. Med. 2011, 208, 505–518. [Google Scholar]
- Pinget, G.V.; Corpuz, T.M.; Stolp, J.; Lousberg, E.L.; Diener, K.R.; Robertson, S.A.; Sprent, J.; Webster, K.E. The majority of murine gammadelta t cells at the maternal-fetal interface in pregnancy produce il-17. Immunol. Cell Biol. 2016, 94, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Kohlgruber, A.C.; Gal-Oz, S.T.; LaMarche, N.M.; Shimazaki, M.; Duquette, D.; Koay, H.F.; Nguyen, H.N.; Mina, A.I.; Paras, T.; Tavakkoli, A.; et al. Gammadelta t cells producing interleukin-17a regulate adipose regulatory t cell homeostasis and thermogenesis. Nat. Immunol. 2018, 19, 464–474. [Google Scholar] [CrossRef]
- Male, V.; Hughes, T.; McClory, S.; Colucci, F.; Caligiuri, M.A.; Moffett, A. Immature nk cells, capable of producing il-22, are present in human uterine mucosa. J. Immunol. 2010, 185, 3913–3918. [Google Scholar] [CrossRef]
- Doisne, J.M.; Balmas, E.; Boulenouar, S.; Gaynor, L.M.; Kieckbusch, J.; Gardner, L.; Hawkes, D.A.; Barbara, C.F.; Sharkey, A.M.; Brady, H.J.; et al. Composition, development, and function of uterine innate lymphoid cells. J. Immunol. 2015, 195, 3937–3945. [Google Scholar] [CrossRef]
- Montaldo, E.; Vacca, P.; Chiossone, L.; Croxatto, D.; Loiacono, F.; Martini, S.; Ferrero, S.; Walzer, T.; Moretta, L.; Mingari, M.C. Unique eomes(+) nk cell subsets are present in uterus and decidua during early pregnancy. Front. Immunol. 2015, 6, 646. [Google Scholar] [CrossRef] [PubMed]
- Huhn, O.; Zhao, X.; Esposito, L.; Moffett, A.; Colucci, F.; Sharkey, A.M. How do uterine natural killer and innate lymphoid cells contribute to successful pregnancy? Front. Immunol. 2021, 12, 607669. [Google Scholar] [CrossRef] [PubMed]
- Vacca, P.; EMontaldo; Croxatto, D.; Loiacono, F.; Canegallo, F.; Venturini, P.L.; Moretta, L.; Mingari, M.C. Identification of diverse innate lymphoid cells in human decidua. Mucosal. Immunol. 2015, 8, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Romero, R.; Miller, D.; Silva, P.; Panaitescu, B.; Theis, K.R.; Arif, A.; Hassan, S.S.; Gomez-Lopez, N. Innate lymphoid cells at the human maternal-fetal interface in spontaneous preterm labor. Am. J. Reprod. Immunol. 2018, 79, e12820. [Google Scholar] [CrossRef] [PubMed]
- Vacca, P.; Vitale, C.; Munari, E.; Cassatella, M.A.; Mingari, M.C.; Moretta, L. Human innate lymphoid cells: Their functional and cellular interactions in decidua. Front. Immunol. 2018, 9, 1897. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Gu, A.; He, H.; Zhao, Q.; Yu, Y.; Chen, J.; Cheng, Z.; Zhou, P.; Zhou, Q.; Jin, M. Autoimmune thyroid disease disrupts immune homeostasis in the endometrium of unexplained infertility women-a single-cell rna transcriptome study during the implantation window. Front. Endocrinol. 2023, 14, 1185147. [Google Scholar] [CrossRef] [PubMed]
- Vacca, P.; Chiossone, L.; Mingari, M.C.; Moretta, L. Heterogeneity of nk cells and other innate lymphoid cells in human and murine decidua. Front. Immunol. 2019, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Sutton, C.E.; Lalor, S.J.; Sweeney, C.M.; Brereton, C.F.; Lavelle, E.C.; Mills, K.H. Interleukin-1 and il-23 induce innate il-17 production from gammadelta t cells, amplifying th17 responses and autoimmunity. Immunity 2009, 31, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Papotto, P.H.; Goncalves-Sousa, N.; Schmolka, N.; Iseppon, A.; Mensurado, S.; Stockinger, B.; Ribot, J.C.; Silva-Santos, B. Il-23 drives differentiation of peripheral gammadelta17 t cells from adult bone marrow-derived precursors. EMBO Rep. 2017, 18, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Papotto, P.H.; Reinhardt, A.; Prinz, I.; Silva-Santos, B. Innately versatile: Gammadelta17 t cells in inflammatory and autoimmune diseases. J. Autoimmun. 2018, 87, 26–37. [Google Scholar] [CrossRef]
- Bernink, J.H.; Krabbendam, L.; Germar, K.; de Jong, E.; Gronke, K.; Kofoed-Nielsen, M.; Munneke, J.M.; Hazenberg, M.D.; Villaudy, J.; Buskens, C.J.; et al. Interleukin-12 and -23 control plasticity of cd127(+) group 1 and group 3 innate lymphoid cells in the intestinal lamina propria. Immunity 2015, 43, 146–160. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Nakajima, T.; Yoshimura, T.; Yasuda, K.; Kanzaki, H. The inhibitory effect of dienogest, a synthetic steroid, on the growth of human endometrial stromal cells in vitro. Mol. Hum. Reprod. 2001, 7, 341–347. [Google Scholar] [CrossRef]
- Murata, H.; Tsuzuki, T.; Kido, T.; Kakita-Kobayashi, M.; Kida, N.; Hisamatsu, Y.; Okada, H. Progestin-induced heart and neural crest derivatives-expressed transcript 2 inhibits angiopoietin 2 via fibroblast growth factor 9 in human endometrial stromal cells. Reprod. Biol. 2019, 19, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.H.; Huang, J.R.; Mazella, J.; Rosenberg, M.; Tseng, L. Differential effects of progestin and relaxin on the synthesis and secretion of immunoreactive prolactin in long term culture of human endometrial stromal cells. J. Clin. Endocrinol. Metab. 1990, 71, 889–899. [Google Scholar] [CrossRef]
- Yuhki, M.; Kajitani, T.; Mizuno, T.; Aoki, Y.; Maruyama, T. Establishment of an immortalized human endometrial stromal cell line with functional responses to ovarian stimuli. Reprod. Biol. Endocrinol. 2011, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Kakita-Kobayashi, M.; Murata, H.; Nishigaki, A.; Hashimoto, Y.; Komiya, S.; Tsubokura, H.; Kido, T.; Kida, N.; Tsuzuki-Nakao, T.; Matsuo, Y.; et al. Thyroid hormone facilitates in vitro decidualization of human endometrial stromal cells via thyroid hormone receptors. Endocrinology 2020, 161, bqaa049. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta ct) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Wright, J.F.; Bennett, F.; Li, B.; Brooks, J.; Luxenberg, D.P.; Whitters, M.J.; Tomkinson, K.N.; Fitz, L.J.; Wolfman, N.M.; Collins, M.; et al. The human il-17f/il-17a heterodimeric cytokine signals through the il-17ra/il-17rc receptor complex. J. Immunol. 2008, 181, 2799–2805. [Google Scholar] [CrossRef]
- Wu, L.; Zepp, J.; Li, X. Function of act1 in il-17 family signaling and autoimmunity. Adv. Exp. Med. Biol. 2012, 946, 223–235. [Google Scholar]
- Weng, H.Y.; Hsu, M.J.; Wang, C.C.; Chen, B.C.; Hong, C.Y.; Chen, M.C.; Chiu, W.T.; Lin, C.H. Zerumbone suppresses ikkalpha, akt, and foxo1 activation, resulting in apoptosis of gbm 8401 cells. J. Biomed. Sci. 2012, 19, 86. [Google Scholar] [CrossRef]
- Tanaka, S.; Honda, Y.; Sawachika, M.; Futani, K.; Yoshida, N.; Kodama, T. Degradation of stk16 via kctd17 with ubiquitin–proteasome system in relation to sleep–wake cycle. Kinases Phosphatases 2023, 1, 14–22. [Google Scholar] [CrossRef]
- Guilloteau, K.; Paris, I.; Pedretti, N.; Boniface, K.; Juchaux, F.; Huguier, V.; Guillet, G.; Bernard, F.X.; Lecron, J.C.; Morel, F. Skin inflammation induced by the synergistic action of il-17a, il-22, oncostatin m, il-1alpha, and tnf-alpha recapitulates some features of psoriasis. J. Immunol. 2010, 184, 5263–5270. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Tanaka, S.; Okada, H. Immune tolerance of the human decidua. J. Clin. Med. 2021, 10, 351. [Google Scholar] [CrossRef] [PubMed]
- Obsil, T.; Obsilova, V. Structural basis for DNA recognition by foxo proteins. Biochim. Biophys. Acta 2011, 1813, 1946–1953. [Google Scholar] [CrossRef] [PubMed]
- Tamura, I.; Jozaki, K.; Sato, S.; Shirafuta, Y.; Shinagawa, M.; Maekawa, R.; Taketani, T.; Asada, H.; Tamura, H.; Sugino, N. The distal upstream region of insulin-like growth factor-binding protein-1 enhances its expression in endometrial stromal cells during decidualization. J. Biol. Chem. 2018, 293, 5270–5280. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Tanaka, S.; Hisamatsu, Y.; Tsubokura, H.; Hashimoto, Y.; Kitada, M.; Okada, H. Transcriptional regulation of lgals9 by hand2 and foxo1 in human endometrial stromal cells in women with regular cycles. Mol. Hum. Reprod. 2021, 27, gaab063. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; van de Pavert, S.A.; Cooper, M.D.; Belz, G.T. The evolution of innate lymphoid cells. Nat. Immunol. 2016, 17, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Einenkel, R.; Ehrhardt, J.; Zygmunt, M.; Muzzio, D.O. Oxygen regulates ilc3 antigen presentation potential and pregnancy-related hormone actions. Reprod. Biol. Endocrinol. 2022, 20, 109. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Takakura, M.; Fujii, R.; Maida, Y.; Bono, Y.; Mizumoto, Y.; Zhang, X.; Kiyono, T.; Kyo, S. The prb-dependent foxo1/igfbp-1 axis is essential for progestin to inhibit endometrial epithelial growth. Cancer Lett. 2013, 336, 68–75. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The il-17 family of cytokines in health and disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Amarilyo, G.; Lourenco, E.V.; Shi, F.D.; La Cava, A. Il-17 promotes murine lupus. J. Immunol. 2014, 193, 540–543. [Google Scholar] [CrossRef]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulen, M.F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K.; et al. Interleukin-23-independent il-17 production regulates intestinal epithelial permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef]
- Crossey, P.A.; Pillai, C.C.; Miell, J.P. Altered placental development and intrauterine growth restriction in igf binding protein-1 transgenic mice. J. Clin. Investig. 2002, 110, 411–418. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells: 10 years on. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef]
- Lim, A.I.; Li, Y.; Lopez-Lastra, S.; Stadhouders, R.; Paul, F.; Casrouge, A.; Serafini, N.; Puel, A.; Bustamante, J.; Surace, L.; et al. Systemic human ilc precursors provide a substrate for tissue ilc differentiation. Cell 2017, 168, 1086–1100.e10. [Google Scholar] [CrossRef]
- Haliloglu, Y.; Ozcan, A.; Erdem, S.; Azizoglu, Z.B.; Bicer, A.; Ozarslan, O.Y.; Kilic, O.; Okus, F.Z.; Demir, F.; Canatan, H.; et al. Characterization of cord blood cd3(+) tcrvalpha7.2(+) cd161(high) t and innate lymphoid cells in the pregnancies with gestational diabetes, morbidly adherent placenta, and pregnancy hypertension diseases. Am. J. Reprod. Immunol. 2022, 88, e13555. [Google Scholar] [CrossRef]
- Pearson, C.; Thornton, E.E.; McKenzie, B.; Schaupp, A.L.; Huskens, N.; Griseri, T.; West, N.; Tung, S.; Seddon, B.P.; Uhlig, H.H.; et al. Ilc3 gm-csf production and mobilisation orchestrate acute intestinal inflammation. eLife 2016, 5, e10066. [Google Scholar] [CrossRef] [PubMed]
- Huhn, O.; Ivarsson, M.A.; Gardner, L.; Hollinshead, M.; Stinchcombe, J.C.; Chen, P.; Shreeve, N.; Chazara, O.; Farrell, L.E.; Theorell, J.; et al. Distinctive phenotypes and functions of innate lymphoid cells in human decidua during early pregnancy. Nat. Commun. 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Wu, Q.; Yang, B.; Wu, C. Estrogen enhanced the expression of il-17 by tissue-resident memory gammadeltat cells from uterus via interferon regulatory factor 4. FASEB J. 2022, 36, e22166. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, T.; Tanaka, Y.; Hamaguchi, M.; Fujii, M.; Shimura, K.; Ogawa, K.; Mori, T.; Kusuki, I.; Fukui, M.; Kitawaki, J. Reduced innate lymphoid cells in the endometrium of women with endometriosis. Am. J. Reprod. Immunol. 2022, 87, e13502. [Google Scholar] [CrossRef] [PubMed]
- Allan, D.S.; Kirkham, C.L.; Aguilar, O.A.; Qu, L.C.; Chen, P.; Fine, J.H.; Serra, P.; Awong, G.; Gommerman, J.L.; Zuniga-Pflucker, J.C.; et al. An in vitro model of innate lymphoid cell function and differentiation. Mucosal. Immunol. 2015, 8, 340–351. [Google Scholar] [CrossRef]
- Zhou, J.; Yue, J.; Yao, Y.; Hou, P.; Zhang, T.; Zhang, Q.; Yi, L.; Mi, M. Dihydromyricetin protects intestinal barrier integrity by promoting il-22 expression in ilc3s through the ampk/sirt3/stat3 signaling pathway. Nutrients 2023, 15, 355. [Google Scholar] [CrossRef]
- Liu, X.; Sun, S.; Liu, D. Il-17d: A less studied cytokine of il-17 family. Int. Arch. Allergy Immunol. 2020, 181, 618–623. [Google Scholar] [CrossRef]
- Starnes, T.; Broxmeyer, H.E.; Robertson, M.J.; Hromas, R. Cutting edge: Il-17d, a novel member of the il-17 family, stimulates cytokine production and inhibits hemopoiesis. J. Immunol. 2002, 169, 642–646. [Google Scholar] [CrossRef]
Figure 1.
Changes in IL1B, IL23A, and IL17D expression in decidualized endometrial stromal cells (EnSCs). The significant increases in IL1B ((A), p < 0.05), IL23A ((B), p < 0.05), and IL17D ((F), p < 0.05) were observed in decidualized EnSCs. No significant differences in IL17A (C), IL17B (D), IL17C (E), IL17E (G), and IL17F (H). * significantly different from the control (p < 0.05). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL, interleukin; IL1B, interleukin 1 beta; IL23A, interleukin 23 subunit alpha.
Figure 1.
Changes in IL1B, IL23A, and IL17D expression in decidualized endometrial stromal cells (EnSCs). The significant increases in IL1B ((A), p < 0.05), IL23A ((B), p < 0.05), and IL17D ((F), p < 0.05) were observed in decidualized EnSCs. No significant differences in IL17A (C), IL17B (D), IL17C (E), IL17E (G), and IL17F (H). * significantly different from the control (p < 0.05). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL, interleukin; IL1B, interleukin 1 beta; IL23A, interleukin 23 subunit alpha.

Figure 2.
Cytokine expression in KC02-44D cells. Significant increases in IL1B ((A), p < 0.05), IL23A ((B), p < 0.05), and IL17D ((F), p < 0.05) were observed in decidualized KC02-44D cells. No significant differences were noted in IL17A (C), IL17B (D), IL17C (E), IL17E (G), and IL17F (H). * significantly different from control (p < 0.05). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL, interleukin; IL1B, interleukin 1 beta; IL23A, interleukin 23 subunit alpha.
Figure 2.
Cytokine expression in KC02-44D cells. Significant increases in IL1B ((A), p < 0.05), IL23A ((B), p < 0.05), and IL17D ((F), p < 0.05) were observed in decidualized KC02-44D cells. No significant differences were noted in IL17A (C), IL17B (D), IL17C (E), IL17E (G), and IL17F (H). * significantly different from control (p < 0.05). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL, interleukin; IL1B, interleukin 1 beta; IL23A, interleukin 23 subunit alpha.

Figure 3.
Responsiveness of decidualized endometrial stromal cells (EnSCs) and KC02-44D cells to ILC3-derived cytokines. Significant increases in IL17RA [(A,D), p < 0.05] and IL17RC [(B,E), p < 0.05] and no significant differences in IL22RA1C (C,F) were observed in decidualized EnSCs and KC02-44D cells. * p < 0.05 vs. control using Welch’s t-test. PCR and agarose gel electrophoresis were performed using distilled water as a negative control to confirm their presence or absence, and IL17RA and IL17RC signals were con-firmed to have specific sample-dependent amplifications (G). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL17RA, interleukin 17 receptor A; IL17RC, interleukin 17 receptor C; IL22RA1, interleukin 22 receptor subunit alpha 1; N.D., no detection.
Figure 3.
Responsiveness of decidualized endometrial stromal cells (EnSCs) and KC02-44D cells to ILC3-derived cytokines. Significant increases in IL17RA [(A,D), p < 0.05] and IL17RC [(B,E), p < 0.05] and no significant differences in IL22RA1C (C,F) were observed in decidualized EnSCs and KC02-44D cells. * p < 0.05 vs. control using Welch’s t-test. PCR and agarose gel electrophoresis were performed using distilled water as a negative control to confirm their presence or absence, and IL17RA and IL17RC signals were con-firmed to have specific sample-dependent amplifications (G). HPRT1, hypoxanthine phosphoribosyltransferase 1; E2, estradiol; MPA, medroxyprogesterone acetate; IL17RA, interleukin 17 receptor A; IL17RC, interleukin 17 receptor C; IL22RA1, interleukin 22 receptor subunit alpha 1; N.D., no detection.

Figure 4.
Effect of IL17A on decidualization markers in KC02-44D cells. Significant upregulation of PRL [(A), p < 0.05], IGFBP1 [(B), p < 0.05], IL15 [(C), p < 0.05], HAND2 [(D), p < 0.05], and IL17D [(E), p < 0.05] was found in the decidualized KC02-44D cells. Additional IL17A stimulation only affected IGFBP1 expression ((B), p < 0.05) in E2 + MPA+ cAMP-treated cells. * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. +: p < 0.05 vs. 0 ng/mL IL17A + decidualization using Welch’s t-test with Bonferroni correction. HPRT1, hypoxanthine phosphoribosyltransferase 1; PRL, prolactin; IGFBP1, insulin-like growth factor-binding protein 1; IL15, interleukin 15; HAND2, heart and neural crest derivatives expressed 2; IL17D, interleukin 17 D; IL17A, interleukin 17 A.
Figure 4.
Effect of IL17A on decidualization markers in KC02-44D cells. Significant upregulation of PRL [(A), p < 0.05], IGFBP1 [(B), p < 0.05], IL15 [(C), p < 0.05], HAND2 [(D), p < 0.05], and IL17D [(E), p < 0.05] was found in the decidualized KC02-44D cells. Additional IL17A stimulation only affected IGFBP1 expression ((B), p < 0.05) in E2 + MPA+ cAMP-treated cells. * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. +: p < 0.05 vs. 0 ng/mL IL17A + decidualization using Welch’s t-test with Bonferroni correction. HPRT1, hypoxanthine phosphoribosyltransferase 1; PRL, prolactin; IGFBP1, insulin-like growth factor-binding protein 1; IL15, interleukin 15; HAND2, heart and neural crest derivatives expressed 2; IL17D, interleukin 17 D; IL17A, interleukin 17 A.

Figure 5.
Effect of IL17A on FOXO1 in KC02-44D. The cytoplasm and nucleolus were stained with PI (purple). FOXO1 was fluorescently labeled using a FOXO1 antibody and an Alexa488-secondary antibody (green). The high-power images are magnified versions of the dashed squares in the low-power images (A). Localization in the nucleus or cytoplasm was determined, whereafter the frequency was calculated relative to the number of cells in the entire image covering an area of 1 mm2 (B). * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. + p < 0.05, vs. decidualization using Welch’s t-test with Bonferroni correction. IL17A, interleukin 17 A; FOXO1, Forkhead box protein O1; PI, propidium iodide.
Figure 5.
Effect of IL17A on FOXO1 in KC02-44D. The cytoplasm and nucleolus were stained with PI (purple). FOXO1 was fluorescently labeled using a FOXO1 antibody and an Alexa488-secondary antibody (green). The high-power images are magnified versions of the dashed squares in the low-power images (A). Localization in the nucleus or cytoplasm was determined, whereafter the frequency was calculated relative to the number of cells in the entire image covering an area of 1 mm2 (B). * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. + p < 0.05, vs. decidualization using Welch’s t-test with Bonferroni correction. IL17A, interleukin 17 A; FOXO1, Forkhead box protein O1; PI, propidium iodide.

Figure 6.
Mechanism of IL17A-dependent FOXO1 migration. After KC02-44D cells were stimulated with or without 10 ng/mL IL17A for 6 days, IKKA, IKKB, and ACTB levels were quantified (A). The obtained signals were analyzed using ImageJ and normalized to ACTB band intensity. Significant suppression of IKKA was noted with IL17A stimulation ((B), p < 0.05). There was no significant difference in IKKB among the three groups (C). * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. IL17A, interleukin 17 A; IKKA (CHUK), component of inhibitor of nuclear factor kappa B kinase complex; IKKB (IKBKB), inhibitor of nuclear factor kappa B kinase subunit beta; ACTB, actin beta.
Figure 6.
Mechanism of IL17A-dependent FOXO1 migration. After KC02-44D cells were stimulated with or without 10 ng/mL IL17A for 6 days, IKKA, IKKB, and ACTB levels were quantified (A). The obtained signals were analyzed using ImageJ and normalized to ACTB band intensity. Significant suppression of IKKA was noted with IL17A stimulation ((B), p < 0.05). There was no significant difference in IKKB among the three groups (C). * p < 0.05 vs. control using Welch’s t-test with Bonferroni correction. IL17A, interleukin 17 A; IKKA (CHUK), component of inhibitor of nuclear factor kappa B kinase complex; IKKB (IKBKB), inhibitor of nuclear factor kappa B kinase subunit beta; ACTB, actin beta.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Patient information.
| Sample No. | Materials | Methods | Age, Years | Menstrual Cycle Phase at the Time of Collection | |
|---|---|---|---|---|---|
| 1 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 50 | Proliferative |
| 2 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 45 | Mid-secretory |
| 3 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 48 | Late secretory |
| 4 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 50 | Mid-secretory |
| 5 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 44 | Late secretory |
| 6 | Primary culture EnSCs | Treated with E2 + MPA for 12 days | RT-qPCR | 49 | Proliferative |
RT-qPCR, reverse transcription–quantitative polymerase chain reaction; E2, estradiol; MPA, medroxyprogesterone acetate; EnSC, endometrial stromal cells.
Table 2.
Primers for qPCR.
| Gene Symbol | Definition | Primer Name | Sequence (5′-3′) |
|---|---|---|---|
| HPRT1 | Hypoxanthine Phosphoribosyltransferase 1 | 895F | CTAGTTCTGTGGCCATCTGCTTAG |
| 1034R | GGGAACTGATAGTCTATAGGCTCATAGTG | ||
| PRL | Prolactin | 374F | ATTCGATAAACGGTATACCCATGGC |
| 623R | TTGCTCCTCAATCTCTACAGCTTTG | ||
| IGFBP1 | Insulin-like Growth Factor Binding Protein 1 | 636F | CTATGATGGCTCGAAGGCTC |
| 791R | TTCTTGTTGCAGTTTGGCAG | ||
| HAND2 | Heart and Neural Crest Derivatives expressed 2 | 1479F | AGAGGAAGAAGGAGCTGAACGA |
| 1552R | CGTCCGGCCTTTGGTTTT | ||
| IL15 | Interleukin 15 | 165F | GTTCACCCCAGTTGCAAAGT |
| 351R | CCTCCAGTTCCTCACATTC | ||
| IL1B | Interleukin 1 beta | 162F | AGCTGATGGCCCTAAACAGATG |
| 305R | TTGTCCATGGCCACAACAAC | ||
| IL23A | Interleukin23, alpha subunit p19 | 71F | ATCAGGCTCAAAGCAAGTGG |
| 196R | AGCAACAGCAGCATTACAGC | ||
| IL17A | Interleukin 17A | 1685F | TCTCTTCCTCAAGCAACACTCC |
| 1777R | AAAGTTCGTTCTGCCCCATC | ||
| IL17B | Interleukin 17B | 594F | GCACCTGCATCTTCTGAATCAC |
| 667R | ACAAAGGTGCAAGGAGGATG | ||
| IL17C | Interleukin 17C | 883F | TGCAGAAAAGGTGTCACACG |
| 1012R | AAACAGGGGTACTTCCAAGGAG | ||
| IL17D | Interleukin 17D | 1791F | TGGAACGTGACATCTTTGCC |
| 1925R | AAGCCTCCAGATTGATCTCTGC | ||
| IL17E (IL25) | Interleukin 17E | 736F | AGGCTGTACCGTGTTTCCTTAG |
| 862R | CCTTCATGGCAAGTGGTTGTAC | ||
| IL17F | Interleukin 17F | 259F | ATGAAAACCAGCGCGTTTCC |
| 398R | ATTGATGCAGCCCAAGTTCC | ||
| IL17RA | Interleukin 17 Receptor A | 793F | TGACCAGTTTTCCGCACATG |
| 923R | ACAGCACCCTTTAAGGTTGC | ||
| IL17RC | Interleukin 17 Receptor C | 877F | TGCAGTTTGGTCAGTCTGTG |
| 1000R | TGCTGTGTGTGGTTGAGTTC | ||
| IL22RA1 | Interleukin 22 Receptor Subunit Alpha 1 | 652F | TGGCACCATCATGATTTGCG |
| 780R | AAGCCCATGGAGAACAGGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tanaka, S.; Sawachika, M.; Yoshida, N.; Futani, K.; Murata, H.; Okada, H. IL17A Suppresses IGFBP1 in Human Endometrial Stromal Cells. Reprod. Med. 2024, 5, 43-56. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed5020006
AMA Style
Tanaka S, Sawachika M, Yoshida N, Futani K, Murata H, Okada H. IL17A Suppresses IGFBP1 in Human Endometrial Stromal Cells. Reproductive Medicine. 2024; 5(2):43-56. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed5020006
Chicago/Turabian StyleTanaka, Susumu, Misa Sawachika, Namika Yoshida, Kensuke Futani, Hiromi Murata, and Hidetaka Okada. 2024. "IL17A Suppresses IGFBP1 in Human Endometrial Stromal Cells" Reproductive Medicine 5, no. 2: 43-56. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed5020006