
An Overview of the Mechanisms of Action and Administration Technologies of the Essential Oils Used as Green Insecticides


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
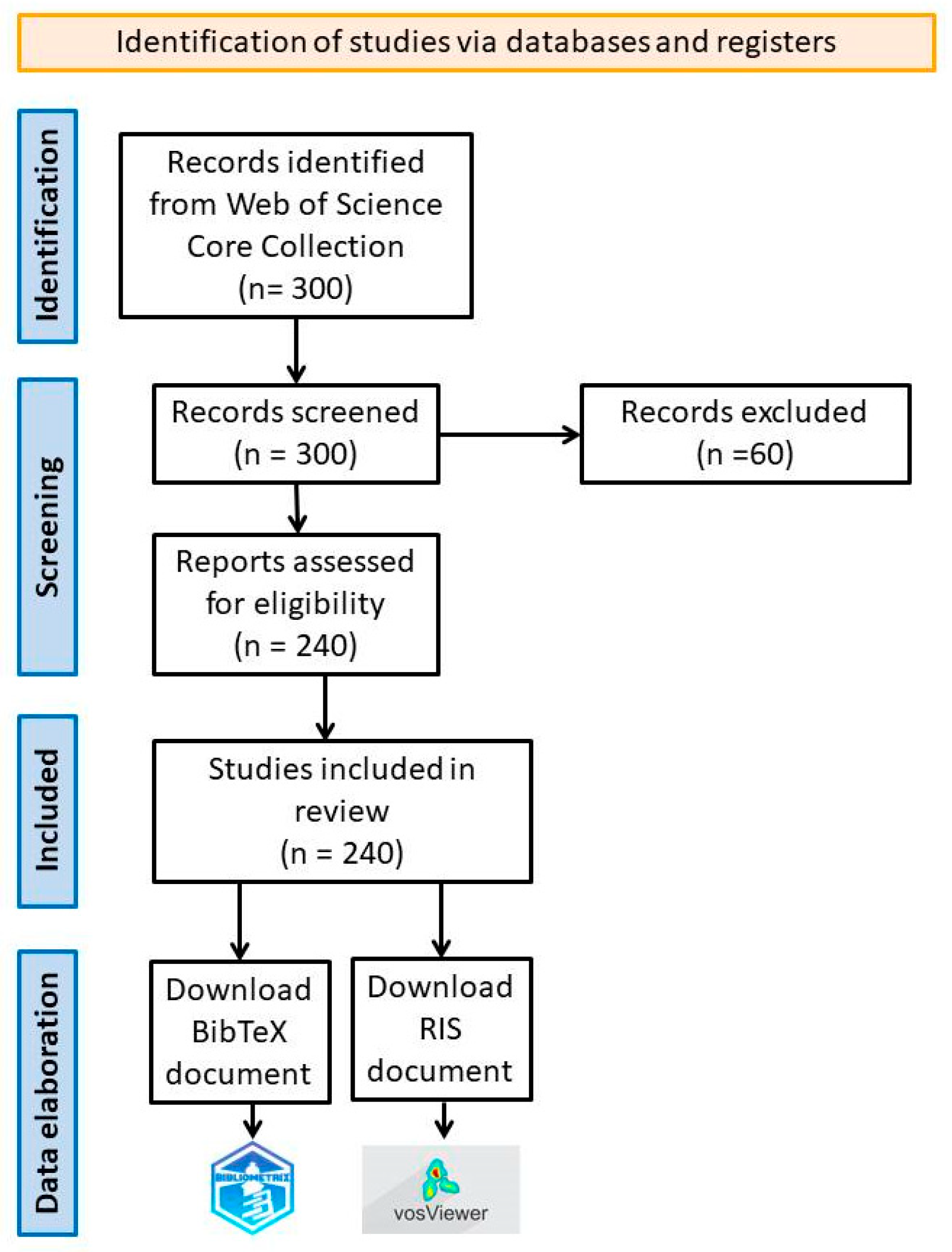
2. Methodology
3. Results and Discussions
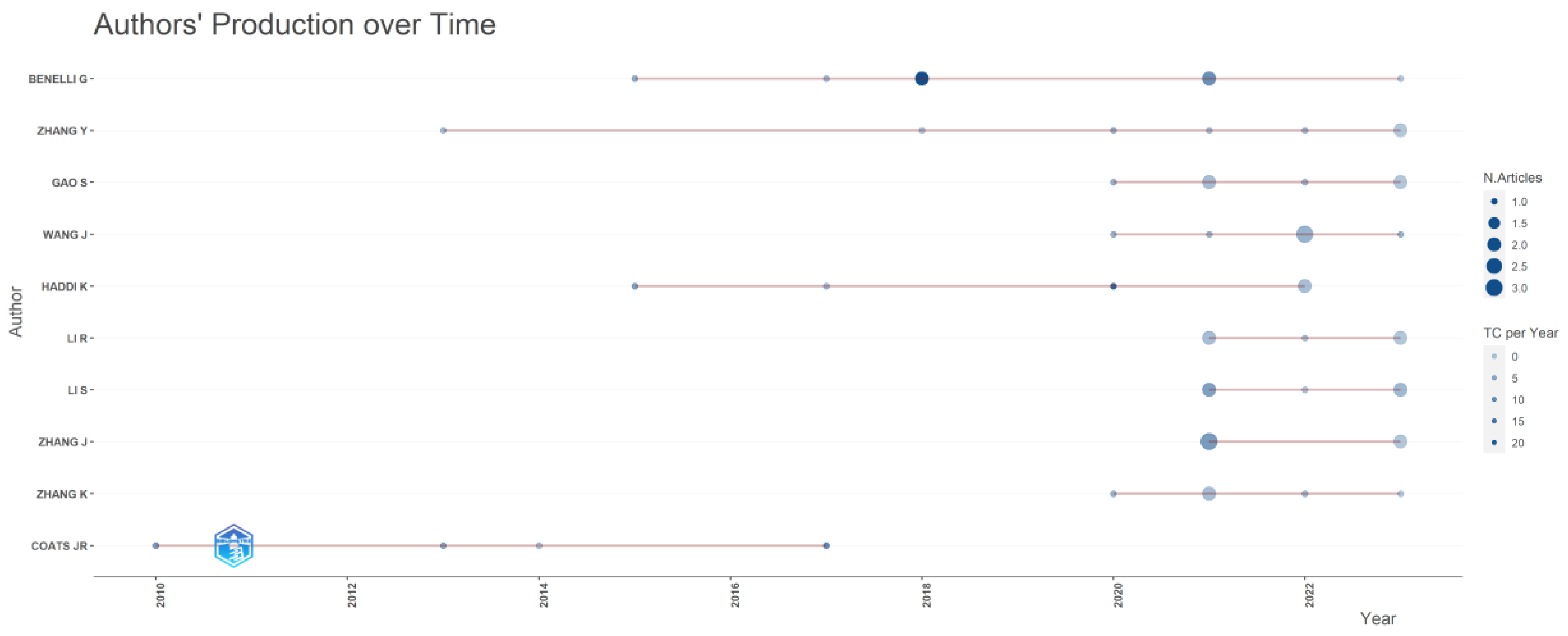
3.1. Bibliometric Analysis Regarding the Mechanisms of Action of the Essential Oils on Insects
3.2. Application Technologies of the Essential Oils: From Laboratory Experiments to Field Results
- −
- −
- −
3.2.1. Nanostructured Formulations of the Essential Oil—Advantages and Disadvantages
3.2.2. Mechanisms of Action of Essential Oils in Insect Species
- Changes in the activity of acetylcholinesterases (AChEs)
- Inhibition of octopamine and tyramine receptors (OARs/TARs)
- Inhibition of GABA (Gamma-amminobutyric acid) receptors
- Glutamate-gated chloride channels (GluCls)
- Antioxidant defense enzymatic systems
- Dysregulation of insect development in different stages of the life cycle
- Cuticular permeability and its role in sensitivity/resistance of the insects to the essential oils
- Repellent and deterrent effects on insects
3.2.3. The Synergistic Action of Essential Oils and Their Components on Insects
4. Conclusions
- First, research is required to investigate the effects of essential oils that have shown insecticidal properties in laboratory tests when applied in open-field conditions.
- The impact of the essential oils on non-target species must be studied in parallel with their effect against harmful insects; although they are considered green insecticides, they are not without action on other species, besides those they directly target. Insects, having similar anatomical structures, are assumed to be susceptible to the effects of the same bioactive substances.
- It is necessary to carry out studies on the economic efficiency of the use of nanoformulations in the distribution of essential oils in the open field. Although many studies show their increased effectiveness compared to bulk oils, the cost/efficiency ratio is important to ascertain.
- The mechanisms of action of essential oils on insects, although they have experienced a significant increase in recent years, are still insufficiently known. Chemoinformatics can be used to predict the effects of essential oil compounds on the molecular targets of insecticides.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Pavela, R. History, Presence and Perspective of Using Plant Extracts as Commercial Botanical Insecticides and Farm Products for Protection against Insects—A Review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar] [CrossRef]
- Bava, R.; Castagna, F.; Palma, E.; Marrelli, M.; Conforti, F.; Musolino, V.; Carresi, C.; Lupia, C.; Ceniti, C.; Tilocca, B.; et al. Essential Oils for a Sustainable Control of Honeybee Varroosis. Vet. Sci. 2023, 10, 308. [Google Scholar] [CrossRef]
- Müller, G.C.; Junnila, A.; Butler, J.; Kravchenko, V.D.; Revay, E.E.; Weiss, R.W.; Schlein, Y. Efficacy of the Botanical Repellents Geraniol, Linalool, and Citronella against Mosquitoes. J. Vector Ecol. 2009, 34, 2–8. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, H.; Hou, R.; Wu, J.; Wang, Y.; Huang, S.; Cheng, D.; Xu, H.; Zhang, Z. Effects of Sublethal Azadirachtin on the Immune Response and Midgut Microbiome of Apis cerana cerana (Hymenoptera: Apidae). Ecotoxicol. Environ. Saf. 2022, 229, 113089. [Google Scholar] [CrossRef]
- Ruiz, M.J.; Juárez, M.L.; Alzogaray, R.A.; Arrighi, F.; Arroyo, L.; Gastaminza, G.; Willink, E.; Bardón, A.; Vera, M.T. Oviposition Behaviour and Larval Development of Anastrepha fraterculus from Argentina in Citrus. Entomol. Exp Appl. 2015, 157, 198–213. [Google Scholar] [CrossRef]
- Da Silva, I.M.; Zanuncio, J.C.; Brügger, B.P.; Soares, M.A.; Zanuncio, A.J.V.; Wilcken, C.F.; Tavares, W.D.S.; Serrão, J.E.; Sediyama, C.S. Selectivity of the Botanical Compounds to the Pollinators Apis mellifera and Trigona hyalinata (Hymenoptera: Apidae). Sci. Rep. 2020, 10, 4820. [Google Scholar] [CrossRef]
- Andrade, L.H.D.; Oliveira, J.V.D.; Breda, M.O.; Marques, E.J.; Lima, I.M.D.M. Effects of Botanical Insecticides on the Instantaneous Population Growth Rate of Aphis gossypii Glover (Hemiptera: Aphididae) in Cotton. Acta Sci. Agron. 2012, 34, 119–214. [Google Scholar] [CrossRef]
- Gupta, I.; Singh, R.; Muthusamy, S.; Sharma, M.; Grewal, K.; Singh, H.P.; Batish, D.R. Plant Essential Oils as Biopesticides: Applications, Mechanisms, Innovations, and Constraints. Plants 2023, 12, 2916. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-Tool for Comprehensive Science Mapping Analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Xu, X.; Chen, Q.; Zhu, Z. Evolutionary Overview of Land Consolidation Based on Bibliometric Analysis in Web of Science from 2000 to 2020. Int. J. Environ. Res. Public Health 2022, 19, 3218. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Gostin, I.N.; Popescu, I.E. Evaluation of the Essential Oils Used in the Production of Biopesticides: Assessing Their Toxicity toward Both Arthropod Target Species and Beneficial Pollinators. Agriculture 2023, 14, 81. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, E.J.A.; Carvalho, F.C.; De Castro Oliveira, J.A.; Bertolucci, S.K.V.; Scotti, M.T.; Silveira, C.H.; Guedes, F.C.; Melo, J.O.F.; De Melo-Minardi, R.C.; De Lima, L.H.F. Elucidating the Molecular Mechanisms of Essential Oils’ Insecticidal Action Using a Novel Cheminformatics Protocol. Sci. Rep. 2023, 13, 4598. [Google Scholar] [CrossRef]
- Ikawati, S.; Himawan, T.; Abadi, A.L.; Tarno, H. Toxicity Nanoinsecticide Based on Clove Essential Oil against Tribolium castaneum (Herbst). J. Pestic. Sci. 2021, 46, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Miresmailli, S.; Isman, M.B. Botanical Insecticides Inspired by Plant–Herbivore Chemical Interactions. Trends Plant Sci. 2014, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-E. Biochemical Mechanisms Conferring Cross-Resistance to Fumigant Toxicities of Essential Oils in a Chlorpyrifos-Methyl Resistant Strain of Oryzaephilus surinamensis L. (Coleoptera: Silvanidae). J. Stored Prod. Res. 2002, 38, 157–166. [Google Scholar] [CrossRef]
- Tia, V.E.; Gueu, S.; Cissé, M.; Tuo, Y.; Gnago, A.J.; Konan, E. Bio-insecticidal effects of essential oil nano-emulsion of Lippia multiflora Mold. on major cabbage pests. J. Plant Prot. Res. 2023, 61, 103–109. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Salem, N.Y.; Abd ElNaby, S.S.; Adel, M.M. Characterization of Nanoparticles Loaded with Garlic Essential Oil and Their Insecticidal Activity Against Phthorimaea operculella (Zeller) (PTM) (Lepidoptera: Gelechiidae). Int. J. Nanosci. Nanotechnol. 2021, 17, 147–160. [Google Scholar]
- Ma, S.; Jia, R.; Guo, M.; Qin, K.; Zhang, L. Insecticidal Activity of Essential Oil from Cephalotaxus sinensis and Its Main Components against Various Agricultural Pests. Ind. Crops Prod. 2020, 150, 112403. [Google Scholar] [CrossRef]
- Ngongang, M.D.T.; Eke, P.; Sameza, M.L.; Mback, M.N.L.N.; Lordon, C.D.; Boyom, F.F. Chemical Constituents of Essential Oils from Thymus vulgaris and Cymbopogon citratus and Their Insecticidal Potential against the Tomato Borer, Tuta absoluta (Lepidoptera: Gelechiidae). Int. J. Trop. Insect Sci. 2022, 42, 31–43. [Google Scholar] [CrossRef]
- Krzyżowski, M.; Baran, B.; Łozowski, B.; Francikowski, J. The Effect of Rosmarinus officinalis Essential Oil Fumigation on Biochemical, Behavioral, and Physiological Parameters of Callosobruchus maculatus. Insects 2020, 11, 344. [Google Scholar] [CrossRef]
- De Souza, M.T.; De Souza, M.T.; Bernardi, D.; De Melo, D.J.; Zarbin, P.H.G.; Zawadneak, M.A.C. Insecticidal and Oviposition Deterrent Effects of Essential Oils of Baccharis spp. and Histological Assessment against Drosophila suzukii (Diptera: Drosophilidae). Sci. Rep. 2021, 11, 3944. [Google Scholar] [CrossRef]
- Xavier, V.M.; Message, D.; Picanço, M.C.; Chediak, M.; Júnior, P.A.S.; Ramos, R.S.; Martins, J.C. Acute Toxicity and Sublethal Effects of Botanical Insecticides to Honey Bees. J. Insect Sci. 2015, 15, 137. [Google Scholar] [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F.; Zappalà, L.; Palmeri, V. Bioactivity of Essential Oil-Based Nano-Biopesticides toward Rhyzopertha dominica (Coleoptera: Bostrichidae). Ind. Crops Prod. 2021, 162, 113257. [Google Scholar] [CrossRef]
- El-Helaly, A.A.; EL-Masarawy, M.S.; El-Bendary, H.M. Using Citronella to Protect Bees (Honeybee Apis mellifera L.) from Certain Insecticides and Their Nano Formulations. Braz. J. Biol. 2021, 81, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Neupane, A.C.; Sapakuka, S.; Tao, P.; Kafle, L. Repellancy and Contact Toxicity of Clove Bud Oil and Its Constituents against German Cockroaches, Blatella germanica (Dictyoptera: Blattellidae), under Laboratory Conditions. Int. J. Pest Manag. 2020, 66, 289–297. [Google Scholar] [CrossRef]
- Yammine, J.; Chihib, N.-E.; Gharsallaoui, A.; Ismail, A.; Karam, L. Advances in Essential Oils Encapsulation: Development, Characterization and Release Mechanisms. Polym. Bull. 2023, 81, 3837–3882. [Google Scholar] [CrossRef]
- Mossa, A.-T.H. Green Pesticides: Essential Oils as Biopesticides in Insect-pest Management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef]
- Lucia, A.; Guzmán, E. Emulsions Containing Essential Oils, Their Components or Volatile Semiochemicals as Promising Tools for Insect Pest and Pathogen Management. Adv. Colloid Interface Sci. 2021, 287, 102330. [Google Scholar] [CrossRef] [PubMed]
- Antunes Filho, S.; Dos Santos, M.S.; Dos Santos, O.A.L.; Backx, B.P.; Soran, M.-L.; Opriş, O.; Lung, I.; Stegarescu, A.; Bououdina, M. Biosynthesis of Nanoparticles Using Plant Extracts and Essential Oils. Molecules 2023, 28, 3060. [Google Scholar] [CrossRef] [PubMed]
- Dunan, L.; Malanga, T.; Benhamou, S.; Papaiconomou, N.; Desneux, N.; Lavoir, A.-V.; Michel, T. Effects of Essential Oil-Based Formulation on Biopesticide Activity. Ind. Crops Prod. 2023, 202, 117006. [Google Scholar] [CrossRef]
- Laudani, F.; Campolo, O.; Caridi, R.; Latella, I.; Modafferi, A.; Palmeri, V.; Sorgonà, A.; Zoccali, P.; Giunti, G. Aphicidal Activity and Phytotoxicity of Citrus sinensis Essential-Oil-Based Nano-Insecticide. Insects 2022, 13, 1150. [Google Scholar] [CrossRef] [PubMed]
- Manjesh, K.; Kundu, A.; Dutta, A.; Saha, S.; Neelakanthaiah, B.S. Bio-Insecticidal Nanoemulsions of Essential Oil and Lipid-Soluble Fractions of Pogostemon cablin. Front. Plant Sci. 2022, 13, 874221. [Google Scholar] [CrossRef] [PubMed]
- Sediawan, W.B.; Hartati, I.; Sulistyo, H.; Azis, M.M. A Review and Bibliometric Analysis on Essential Oil Nanoencapsulation. In Proceedings of the 4th Borobudur International Symposium on Science and Technology 2022 (BIS-STE 2022); Setiyo, M., Pambuko, Z.B., Praja, C.B.E., Setiawan, A., Yuliastuti, F., Muliawanti, L., Dewi, V.S., Eds.; Advances in Engineering Research; Atlantis Press International BV: Dordrecht, The Netherlands, 2023; Volume 225, pp. 79–90. ISBN 978-94-6463-283-5. [Google Scholar]
- Pascual-Villalobos, M.J.; López, M.D.; Castañé, C.; Soler, A.; Riudavets, J. Encapsulated Essential Oils as an Alternative to Insecticides in Funnel Traps. J. Econ. Entomol. 2015, 108, 2117–2120. [Google Scholar] [CrossRef] [PubMed]
- Dolma, S.K.; Suresh, P.S.; Singh, P.P.; Sharma, U.; Reddy, S.G.E. Insecticidal Activity of the Extract, Fractions, and Pure Steroidal Saponins of Trillium govanianum W All. Ex D. D on for the Control of Diamondback Moth (Plutella xylostella L.) and Aphid (Aphis craccivora K Och). Pest Manag. Sci. 2021, 77, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Palermo, T.B.; Cappellari, L.D.R.; Chiappero, J.; Meneguzzi, R.D.V.; Gil, S.; Giordano, W.; Banchio, E. How Do Plants Protect Themselves from Insects? A Practical Laboratory Exercise to Illustrate the Defence Mechanisms of the Plant through Secondary Metabolites. J. Biol. Educ. 2022, 1–17. [Google Scholar] [CrossRef]
- Mahran, H.A. Using Nanoemulsions of the Essential Oils of a Selection of Medicinal Plants from Jazan, Saudi Arabia, as a Green Larvicidal against Culex pipiens. PLoS ONE 2022, 17, e0267150. [Google Scholar] [CrossRef]
- Somala, N.; Laosinwattana, C.; Teerarak, M. Formulation Process, Physical Stability and Herbicidal Activities of Cymbopogon nardus Essential Oil-Based Nanoemulsion. Sci. Rep. 2022, 12, 10280. [Google Scholar] [CrossRef]
- Khalil, M.S.; Halawa, S.M.; Azab, M.M.; Morsy, A.R. Toxicity and Biochemical Effects of Citronella, Mustard and Sage Essential Oils and Their Nanoemulsions against Spodoptera littoralis (Boisd.)(Lepidoptera: Noctuidae). Benha J. Appl. Sci. 2023, 8, 79–88. [Google Scholar] [CrossRef]
- Garrido-Miranda, K.A.; Giraldo, J.D.; Schoebitz, M. Essential Oils and Their Formulations for the Control of Curculionidae Pests. Front. Agron. 2022, 4, 876687. [Google Scholar] [CrossRef]
- Ibrahim, S.S. Essential Oil Nanoformulations as a Novel Method for Insect Pest Control in Horticulture. In Horticultural Crops; Kossi Baimey, H., Hamamouch, N., Adjiguita Kolombia, Y., Eds.; IntechOpen: London, UK, 2020; ISBN 978-1-83880-421-3. [Google Scholar]
- Antofie, M.-M.; Sava Sand, C. Drought Stress Study on Nicotiana tabacum L., “Baladi”, an In Vitro Experimental Model. Agriculture 2021, 11, 845. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Nassrallah, A.A.; Abdel-Raheem, M.A.; Elbehery, H.H. Lemon Peel Essential Oil and Its Nano-Formulation to Control Agrotis ipsilon (Lepidoptera: Noctuidae). Sci. Rep. 2023, 13, 17922. [Google Scholar] [CrossRef]
- Ziaee, M.; Sheikhzadeh Takabi, A.; Ebadollahi, A. Fabrication of Carum copticum Essential Oil–Loaded Chitosan Nanoparticles and Evaluation Its Insecticidal Activity for Controlling Rhyzopertha dominica and Tribolium confusum. Front. Plant Sci. 2023, 14, 1187616. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.S.; Abou-Elseoud, W.S.; Elbehery, H.H.; Hassan, M.L. Chitosan-Cellulose Nanoencapsulation Systems for Enhancing the Insecticidal Activity of Citronella Essential Oil against the Cotton Leafworm Spodoptera littoralis. Ind. Crops Prod. 2022, 184, 115089. [Google Scholar] [CrossRef]
- Yang, F.-L.; Li, X.-G.; Zhu, F.; Lei, C.-L. Structural Characterization of Nanoparticles Loaded with Garlic Essential Oil and Their Insecticidal Activity against Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Agric. Food Chem. 2009, 57, 10156–10162. [Google Scholar] [CrossRef] [PubMed]
- De Matos, S.P.; Lucca, L.G.; Koester, L.S. Essential Oils in Nanostructured Systems: Challenges in Preparation and Analytical Methods. Talanta 2019, 195, 204–214. [Google Scholar] [CrossRef]
- Paula, H.C.B.; Sombra, F.M.; Abreu, F.O.M.S.; Paul, R.C.M.D. Lippia Sidoides Essential Oil Encapsulation by Angico Gum/Chitosan Nanoparticles. J. Braz. Chem. Soc. 2010, 21, 2359–2366. [Google Scholar] [CrossRef]
- Aguiar, M.C.S.; Das Graças Fernandes Da Silva, M.F.; Fernandes, J.B.; Forim, M.R. Evaluation of the Microencapsulation of Orange Essential Oil in Biopolymers by Using a Spray-Drying Process. Sci. Rep. 2020, 10, 11799. [Google Scholar] [CrossRef]
- Choi, Y.H.; Van Spronsen, J.; Dai, Y.; Verberne, M.; Hollmann, F.; Arends, I.W.C.E.; Witkamp, G.-J.; Verpoorte, R. Are Natural Deep Eutectic Solvents the Missing Link in Understanding Cellular Metabolism and Physiology? Plant Physiol. 2011, 156, 1701–1705. [Google Scholar] [CrossRef]
- Bajkacz, S.; Adamek, J. Development of a Method Based on Natural Deep Eutectic Solvents for Extraction of Flavonoids from Food Samples. Food Anal. Methods 2018, 11, 1330–1344. [Google Scholar] [CrossRef]
- Mišan, A.; Nađpal, J.; Stupar, A.; Pojić, M.; Mandić, A.; Verpoorte, R.; Choi, Y.H. The Perspectives of Natural Deep Eutectic Solvents in Agri-Food Sector. Crit. Rev. Food Sci. Nutr. 2020, 60, 2564–2592. [Google Scholar] [CrossRef] [PubMed]
- Barradas, T.N.; de Holanda e Silva, K.G. Nanoemulsions of essential oils to improve solubility, stability and permeability: A review. Environ. Chem. Lett. 2021, 19, 1153–1171. [Google Scholar] [CrossRef]
- Maes, C.; Bouquillon, S.; Fauconnier, M.-L. Encapsulation of Essential Oils for the Development of Biosourced Pesticides with Controlled Release: A Review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Hernández, E.; Peña-Chora, G.; Hernández-Velázquez, V.M.; Lormendez, C.C.; Toribio-Jiménez, J.; Romero-Ramírez, Y.; León-Rodríguez, R. The Stingless Bees (Hymenoptera: Apidae: Meliponini): A Review of the Current Threats to Their Survival. Apidologie 2022, 53, 8. [Google Scholar] [CrossRef]
- Liu, X.; Cao, A.; Yan, D.; Ouyang, C.; Wang, Q.; Li, Y. Overview of Mechanisms and Uses of Biopesticides. Int. J. Pest Manag. 2021, 67, 65–72. [Google Scholar] [CrossRef]
- Santana, K.; Do Nascimento, L.D.; Lima E Lima, A.; Damasceno, V.; Nahum, C.; Braga, R.C.; Lameira, J. Applications of Virtual Screening in Bioprospecting: Facts, Shifts, and Perspectives to Explore the Chemo-Structural Diversity of Natural Products. Front. Chem. 2021, 9, 662688. [Google Scholar] [CrossRef] [PubMed]
- Maia, E.H.B.; Assis, L.C.; De Oliveira, T.A.; Da Silva, A.M.; Taranto, A.G. Structure-Based Virtual Screening: From Classical to Artificial Intelligence. Front. Chem. 2020, 8, 343. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, S.H. Which Acetylcholinesterase Functions as the Main Catalytic Enzyme in the Class Insecta? Insect Biochem. Mol. Biol. 2013, 43, 47–53. [Google Scholar] [CrossRef]
- López, M.D.; Pascual-Villalobos, M.J. Mode of Inhibition of Acetylcholinesterase by Monoterpenoids and Implications for Pest Control. Ind. Crops Prod. 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [PubMed]
- Sakthivel, S.; Mohideen, H.S.; Raman, C.; Mohamad, S.B. Potential Acetylcholinesterase Inhibitor Acting on the Pesticide Resistant and Susceptible Cotton Pests. ACS Omega 2022, 7, 20515–20527. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C. Classification and Uses of Organophosphates and Carbamates. In Toxicology of Organophosphate & Carbamate Compounds; Elsevier: Amsterdam, The Netherlands, 2006; pp. 5–24. ISBN 978-0-12-088523-7. [Google Scholar]
- Tong, F.; Islam, R.M.; Carlier, P.R.; Ma, M.; Ekström, F.; Bloomquist, J.R. Effects of Anticholinesterases on Catalysis and Induced Conformational Change of the Peripheral Anionic Site of Murine Acetylcholinesterase. Pestic. Biochem. Physiol. 2013, 106, 79–84. [Google Scholar] [CrossRef]
- Almadiy, A.A.; Nenaah, G.E. Essential Oil of Origanum vulgare, Its Nanoemulsion and Bioactive Monoterpenes as Eco-Friendly Novel Green Pesticides for Controlling Aedes aegypti, the Common Vector of Dengue Virus. J. Essent. Oil Res. 2022, 34, 424–438. [Google Scholar] [CrossRef]
- Liu, J.; Hua, J.; Qu, B.; Guo, X.; Wang, Y.; Shao, M.; Luo, S. Insecticidal Terpenes from the Essential Oils of Artemisia nakaii and Their Inhibitory Effects on Acetylcholinesterase. Front. Plant Sci. 2021, 12, 720816. [Google Scholar] [CrossRef]
- Yeom, H.-J.; Kang, J.S.; Kim, G.-H.; Park, I.-K. Insecticidal and Acetylcholine Esterase Inhibition Activity of Apiaceae Plant Essential Oils and Their Constituents against Adults of German Cockroach (Blattella germanica). J. Agric. Food Chem. 2012, 60, 7194–7203. [Google Scholar] [CrossRef]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Glavan, G.; Novak, S.; Božič, J.; Jemec Kokalj, A. Comparison of Sublethal Effects of Natural Acaricides Carvacrol and Thymol on Honeybees. Pestic. Biochem. Physiol. 2020, 166, 104567. [Google Scholar] [CrossRef]
- Alves, M.D.S.; Campos, I.M.; Brito, D.D.M.C.D.; Cardoso, C.M.; Pontes, E.G.; Souza, M.A.A.D. Efficacy of Lemongrass Essential Oil and Citral in Controlling Callosobruchus maculatus (Coleoptera: Chrysomelidae), a Post-Harvest Cowpea Insect Pest. Crop Prot. 2019, 119, 191–196. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Olumuyiwa, T.A.; Olasehinde, T.A.; Ademiluyi, A.O.; Adeyemo, A.C. Insecticidal Activity of Essential Oil from Orange Peels (Citrus sinensis) against Tribolium confusum, Callosobruchus maculatus and Sitophilus oryzae and Its Inhibitory Effects on Acetylcholinesterase and Na+/K+-ATPase Activities. Phytoparasitica 2017, 45, 501–508. [Google Scholar] [CrossRef]
- Beyaoui, A.; Jlizi, S.; Ascrizzi, R.; Flamini, G.; Harrath, A.H.; Ben Jannet, H. Chemical Profiling and Biological Assessment of Trunk Bark Essential Oil from Eucalyptus camaldulensis: In Vitro Study Coupled with Chemoinformatics Calculations. J. Mol. Struct. 2024, 1300, 137120. [Google Scholar] [CrossRef]
- Lokeshwari, D.; Krishna Kumar, N.K.; Manjunatha, H. Multiple Mutations on the Second Acetylcholinesterase Gene Associated with Dimethoate Resistance in the Melon Aphid, Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2016, 109, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Prasannakumar, N.R.; Jyothi, N.; Saroja, S.; Lokesha, A.N. Insecticidal Properties of Ocimum basilicum and Mentha piperita Essential Oils against South American Tomato Moth, Phthorimaea absoluta (Meyrick) (Lepidoptera: Gelichiidae). Pestic. Biochem. Physiol. 2023, 190, 105329. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.D.; Maqueira, B. Insect Octopamine Receptors: A New Classification Scheme Based on Studies of Cloned Drosophila G-Protein Coupled Receptors. Invert. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Roeder, T. Tyramine and Octopamine: Ruling Behavior and Metabolism. Annu. Rev. Entomol. 2005, 50, 447–477. [Google Scholar] [CrossRef] [PubMed]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of Octopaminergic Receptors by Essential Oil Constituents Isolated from Aromatic Plants: Possible Mode of Action against Insect Pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Finetti, L.; Roeder, T.; Calò, G.; Bernacchia, G. The Insect Type 1 Tyramine Receptors: From Structure to Behavior. Insects 2021, 12, 315. [Google Scholar] [CrossRef]
- Orchard, I. Octopamine in Insects: Neurotransmitter, Neurohormone, and Neuromodulator. Can. J. Zool. 1982, 60, 659–669. [Google Scholar] [CrossRef]
- Pauls, D.; Blechschmidt, C.; Frantzmann, F.; El Jundi, B.; Selcho, M. A Comprehensive Anatomical Map of the Peripheral Octopaminergic/Tyraminergic System of Drosophila melanogaster. Sci. Rep. 2018, 8, 15314. [Google Scholar] [CrossRef]
- Roeder, T.; Seifert, M.; Kähler, C.; Gewecke, M. Tyramine and Octopamine: Antagonistic Modulators of Behavior and Metabolism: Octopamine and Tyramine in Insects. Arch. Insect Biochem. Physiol. 2003, 54, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Lapied, B.; Jankowski, W.; Stankiewicz, M. The Unusual Action of Essential Oil Component, Menthol, in Potentiating the Effect of the Carbamate Insecticide, Bendiocarb. Pestic. Biochem. Physiol. 2019, 158, 101–111. [Google Scholar] [CrossRef]
- Enan, E.E. Molecular and Pharmacological Analysis of an Octopamine Receptor from American Cockroach and Fruit Fly in Response to Plant Essential Oils. Arch. Insect. Biochem. Physiol. 2005, 59, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Lin, J.; Li, X.; Huang, J.; Liang, X.; Li, Y.; Bai, M.; He, H.; Lin, F.; Xu, H.; et al. Changes in Dopamine and Octopamine Levels Caused Disordered Behaviour in Red Imported Fire Ants Exposed to Cinnamon Essential Oils. Ind. Crops Prod. 2023, 199, 116801. [Google Scholar] [CrossRef]
- Enan, E. Insecticidal Activity of Essential Oils: Octopaminergic Sites of Action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, A.B.; Braza, M.K.E.; Nellas, R.B. The Interaction and Mechanism of Monoterpenes with Tyramine Receptor (SoTyrR) of Rice Weevil (Sitophilus oryzae). SN Appl. Sci. 2020, 2, 1592. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Singh, V.K.; Kedia, A.; Das, S.; Dubey, N.K. Essential Oils and Their Bioactive Compounds as Eco-Friendly Novel Green Pesticides for Management of Storage Insect Pests: Prospects and Retrospects. Env. Sci. Pollut. Res. 2021, 28, 18918–18940. [Google Scholar] [CrossRef]
- Devrnja, N.; Milutinović, M.; Savić, J. When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability 2022, 14, 6847. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of Monoterpenoid Insecticides on [3H]-TBOB Binding in House Fly GABA Receptor and 36Cl− Uptake in American Cockroach Ventral Nerve Cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Ozoe, Y. γ-Aminobutyrate- and Glutamate-Gated Chloride Channels as Targets of Insecticides. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 44, pp. 211–286. ISBN 978-0-12-394389-7. [Google Scholar]
- Guo, L.; Qiao, X.; Haji, D.; Zhou, T.; Liu, Z.; Whiteman, N.K.; Huang, J. Convergent Resistance to GABA Receptor Neurotoxins through Plant–Insect Coevolution. Nat. Ecol. Evol. 2023, 7, 1444–1456. [Google Scholar] [CrossRef]
- Qian, K.; Jiang, C.; Guan, D.; Zhuang, A.; Meng, X.; Wang, J. Characterization of Glutamate-Gated Chloride Channel in Tribolium castaneum. Insects 2023, 14, 580. [Google Scholar] [CrossRef]
- El Hassani, A.K.; Schuster, S.; Dyck, Y.; Demares, F.; Leboulle, G.; Armengaud, C. Identification, Localization and Function of Glutamate-gated Chloride Channel Receptors in the Honeybee Brain. Eur. J. Neurosci. 2012, 36, 2409–2420. [Google Scholar] [CrossRef]
- Abdel Rahman, K.M. Effects of Abamectin and Fipronil Insecticides on the Brain and Compound Eyes of the Embryo of Heteracris littoralis (Rambur) (Orthoptera: Acrididae). Int. J. Trop. Insect. Sci. 2023, 43, 1237–1241. [Google Scholar] [CrossRef]
- Lynagh, T.; Cromer, B.A.; Dufour, V.; Laube, B. Comparative Pharmacology of Flatworm and Roundworm Glutamate-Gated Chloride Channels: Implications for Potential Anthelmintics. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 244–255. [Google Scholar] [CrossRef]
- Yang, L.-H.; Huang, H.; Wang, J.-J. Antioxidant Responses of Citrus Red Mite, Panonychus citri (McGregor) (Acari: Tetranychidae), Exposed to Thermal Stress. J. Insect Physiol. 2010, 56, 1871–1876. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fu, Z.-X.; Kang, Z.-W.; Li, H.; Liu, T.-X.; Wang, D. Identification and Characterization of Antioxidant Enzyme Genes in Parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) and Expression Profiling Analysis under Temperature Stress. Insects 2022, 13, 447. [Google Scholar] [CrossRef]
- Ercan, F.S.; Baş, H.; Azarkan, S.Y. In Silico Detection of Cucurbitacin-E on Antioxidant Enzymes of Model Organism Galleria mellonella L. (Lepidoptera: Pyralidae) and Variation of Antioxidant Enzyme Activities and Lipid Peroxidation in Treated Larvae. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 130. [Google Scholar] [CrossRef]
- Isman, M.B. Commercial Development of Plant Essential Oils and Their Constituents as Active Ingredients in Bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Moustafa, M.A.M.; Hassan, N.N.; Alfuhaid, N.A.; Amer, A.; Awad, M. Insights into the Toxicity, Biochemical Activity, and Molecular Docking of Cymbopogon citratus Essential Oils and Citral on Spodoptera littoralis (Lepidoptera: Noctuidae). J. Econ. Entomol. 2023, 116, 1185–1195. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyamoto, M.; Murakami, A.; Ohigashi, H.; Osawa, T.; Uchida, K. A Phase II Detoxification Enzyme Inducer from Lemongrass: Identification of Citral and Involvement of Electrophilic Reaction in the Enzyme Induction. Biochem. Biophys. Res. Commun. 2003, 302, 593–600. [Google Scholar] [CrossRef]
- Omotoso, S.E.; Akinpelu, B.A.; Soyelu, O.J. Insecticidal Effect of Lemongrass Oil on Behavioural Responses and Biochemical Changes in Cowpea Weevil, Callosobruchus maculatus (Fabricius). J. Phytopathol. Pest Manag. 2020, 7, 14–30. [Google Scholar]
- Rajkumar, V.; Gunasekaran, C.; Christy, I.K.; Dharmaraj, J.; Chinnaraj, P.; Paul, C.A. Toxicity, Antifeedant and Biochemical Efficacy of Mentha piperita L. Essential Oil and Their Major Constituents against Stored Grain Pest. Pestic. Biochem. Physiol. 2019, 156, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, A.O.; Olajuyigbe, F.M.; Ajele, J.O.; Adedire, C.O. Activity of the Antioxidant Defense System in a Typical Bioinsecticide-and Synthetic Insecticide-Treated Cowpea Storage Beetle Callosobrochus maculatus F. (Coleoptera: Chrysomelidae). Int. J. Insect. Sci. 2014, 6, IJIS.S19434. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.O.; Nouzova, M.; Fruttero, L.L.; Leyria, J.; Ligabue-Braun, R.; Noriega, F.G.; Canavoso, L.E. Role of Methoprene-Tolerant in the Regulation of Oogenesis in Dipetalogaster maxima. Sci. Rep. 2022, 12, 14195. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Drenaggi, E.; Desneux, N.; Maggi, F. Phytol, (E)-Nerolidol and Spathulenol from Stevia rebaudiana Leaf Essential Oil as Effective and Eco-Friendly Botanical Insecticides against Metopolophium dirhodum. Ind. Crops Prod. 2020, 155, 112844. [Google Scholar] [CrossRef]
- Ghoneim, K.; Hamadah, K.; Selim, S.; Waheeb, H. Biopesticidal Potential of Nerolidol, a Sesquiterpene Compound, and Its Drastic Impact on Growth and Metamorphosis of the Cotton Leafworm Spodoptera littoralis (Lepidoptera: Noctuidae). Sch. Acad. J. Biosci. 2021, 9, 36–57. [Google Scholar] [CrossRef]
- Raikhel, A.S.; Brown, M.R.; Belles, X. Hormonal Control of Reproductive Processes. In Comprehensive Molecular Insect Science; Elsevier: Amsterdam, The Netherlands, 2005; pp. 433–491. ISBN 978-0-444-51924-5. [Google Scholar]
- Pan, X.; Connacher, R.P.; O’Connor, M.B. Control of the Insect Metamorphic Transition by Ecdysteroid Production and Secretion. Curr. Opin. Insect Sci. 2021, 43, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.A.A.; De Souza, N.A.; Feder, M.D.; Da Silva, C.E.; Garcia, E.D.S.; Azambuja, P.; Gonzalez, M.S.; Rangel, E.F. Effects of Azadirachtin on the Development and Mortality of Lutzomyia longipalpis Larvae (Diptera: Psychodidae: Phlebotominae). J. Med. Entomol. 2006, 43, 262–266. [Google Scholar] [CrossRef]
- Rembold, H. The Azadirachtins—Their Potential for Insect Control. Econ. Med. Plant Res. 1989, 3, 57–72. [Google Scholar]
- Bass, C.; Jones, C.M. Mosquitoes Boost Body Armor to Resist Insecticide Attack. Proc. Natl. Acad. Sci. USA 2016, 113, 9145–9147. [Google Scholar] [CrossRef]
- Gao, L.; Qiao, H.; Wei, P.; Moussian, B.; Wang, Y. Xenobiotic Responses in Insects. Arch. Insect Biochem. Physiol. 2022, 109, e21869. [Google Scholar] [CrossRef]
- Kim, S.; Yoon, J.; Tak, J.-H. Synergistic Mechanism of Insecticidal Activity in Basil and Mandarin Essential Oils against the Tobacco Cutworm. J. Pest Sci. 2021, 94, 1119–1131. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, J.; Lin, S.; He, J.; Deng, Y.; He, J.; Cheng, D. The Synergistic Effects of Rosehip Oil and Matrine against Icerya aegyptiaca (Douglas) (Hemiptera: Coccoidea) and the Underlying Mechanisms. Pest Manag. Sci. 2022, 78, 3424–3432. [Google Scholar] [CrossRef]
- Yoon, J.; Tak, J.-H. Cuticular Property Affects the Insecticidal Synergy of Major Constituents in Thyme Oil against Houseflies, Musca domestica. Sci. Rep. 2023, 13, 12654. [Google Scholar] [CrossRef]
- Ren, Y.; Li, Y.; Ju, Y.; Zhang, W.; Wang, Y. Insect Cuticle and Insecticide Development. Arch. Insect Biochem. Physiol. 2023, 114, e22057. [Google Scholar] [CrossRef]
- Karabörklü, S.; Ayvaz, A. A Comprehensive Review of Effective Essential Oil Components in Stored-Product Pest Management. J. Plant Dis. Prot. 2023, 130, 449–481. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, K.; You, C.; Wang, C.; Geng, Z.; Su, Y.; Wang, Y.; Du, S.; Deng, Z. Contact Toxicity and Repellency of the Essential Oil from Mentha haplocalyx BRIQ. against Lasioderma serricorne. Chem. Biodivers. 2015, 12, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Njuguna, M.J.; Hutchins, M.M.; Karenga, S. Efficacy of Essential Oils from Ocimum kenyense as a Biopesticide against Aphis gosypii, Thrips tabaci and Bemisia tabaci. Int. J. Adv. Res. 2021, 4, 14–26. [Google Scholar] [CrossRef]
- Sayed, S.; Soliman, M.M.; Al-Otaibi, S.; Hassan, M.M.; Elarrnaouty, S.-A.; Abozeid, S.M.; El-Shehawi, A.M. Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae). Plants 2022, 11, 463. [Google Scholar] [CrossRef] [PubMed]
- Gvozdenac, S.; Kiprovski, B.; Aćimović, M.; Jeremić, J.S.; Cvetković, M.; Bursić, V.; Ovuka, J. Repellent Activity of Cymbopogon citratus Essential Oil Against Four Major Stored Product Pests: Plodia interpunctella, Sitophilus oryzae, Acanthoscelides obtectus and Tribolium castaneum. Contemp. Agric. 2021, 70, 140–148. [Google Scholar] [CrossRef]
- Parreira, D.S.; Alcántara-de La Cruz, R.; Rodrigues Dimaté, F.A.; Batista, L.D.; Ribeiro, R.C.; Rigueira Ferreira, G.A.; Zanuncio, J.C. Bioactivity of Ten Essential Oils on the Biological Parameters of Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) Adults. Ind. Crops Prod. 2019, 127, 11–15. [Google Scholar] [CrossRef]
- Santana, A.D.S.; Baldin, E.L.L.; Santos, T.L.B.D.; Baptista, Y.A.; Santos, M.C.D.; Lima, A.P.S.; Tanajura, L.S.; Vieira, T.M.; Crotti, A.E.M. Synergism between Essential Oils: A Promising Alternative to Control Sitophilus zeamais (Coleoptera: Curculionidae). Crop Prot. 2022, 153, 105882. [Google Scholar] [CrossRef]
- Akers, R.P.; Getz, W.M. A Test of Identified Response Classes among Olfactory Receptor Neurons in the Honey-Bee Worker. Chem. Senses 1992, 17, 191–209. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popescu, I.E.; Gostin, I.N.; Blidar, C.F. An Overview of the Mechanisms of Action and Administration Technologies of the Essential Oils Used as Green Insecticides. AgriEngineering 2024, 6, 1195-1217. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering6020068
Popescu IE, Gostin IN, Blidar CF. An Overview of the Mechanisms of Action and Administration Technologies of the Essential Oils Used as Green Insecticides. AgriEngineering. 2024; 6(2):1195-1217. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering6020068
Chicago/Turabian StylePopescu, Irinel Eugen, Irina Neta Gostin, and Cristian Felix Blidar. 2024. "An Overview of the Mechanisms of Action and Administration Technologies of the Essential Oils Used as Green Insecticides" AgriEngineering 6, no. 2: 1195-1217. https://0-doi-org.brum.beds.ac.uk/10.3390/agriengineering6020068