

Assessing Prospects of Integrating Asian Carp Polyculture in Europe: A Nature-Based Solution under Climate Change?

 , , ,
, , ,
Abstract
:1. Introduction
- Identifying global carp distribution centers: Utilizing species distribution modeling techniques, we seek to pinpoint and compare the primary centers of carp distribution worldwide.
- Analyzing carp spread in Europe: We delve into the likelihood and prospects for the expansion of Asian carp populations in Europe, particularly in northeastern regions, and explore the factors influencing this spread.
- Assessing polyculture potential: We examine the viability of incorporating Asian carp species into polyculture systems by comparing their natural biotopes (ecological niches, niche clustering) to those found in their new distribution centers in Europe and America.
2. Materials and Methods
2.1. Occurrence Data Collection
2.2. Environmental Data
2.3. Model Building
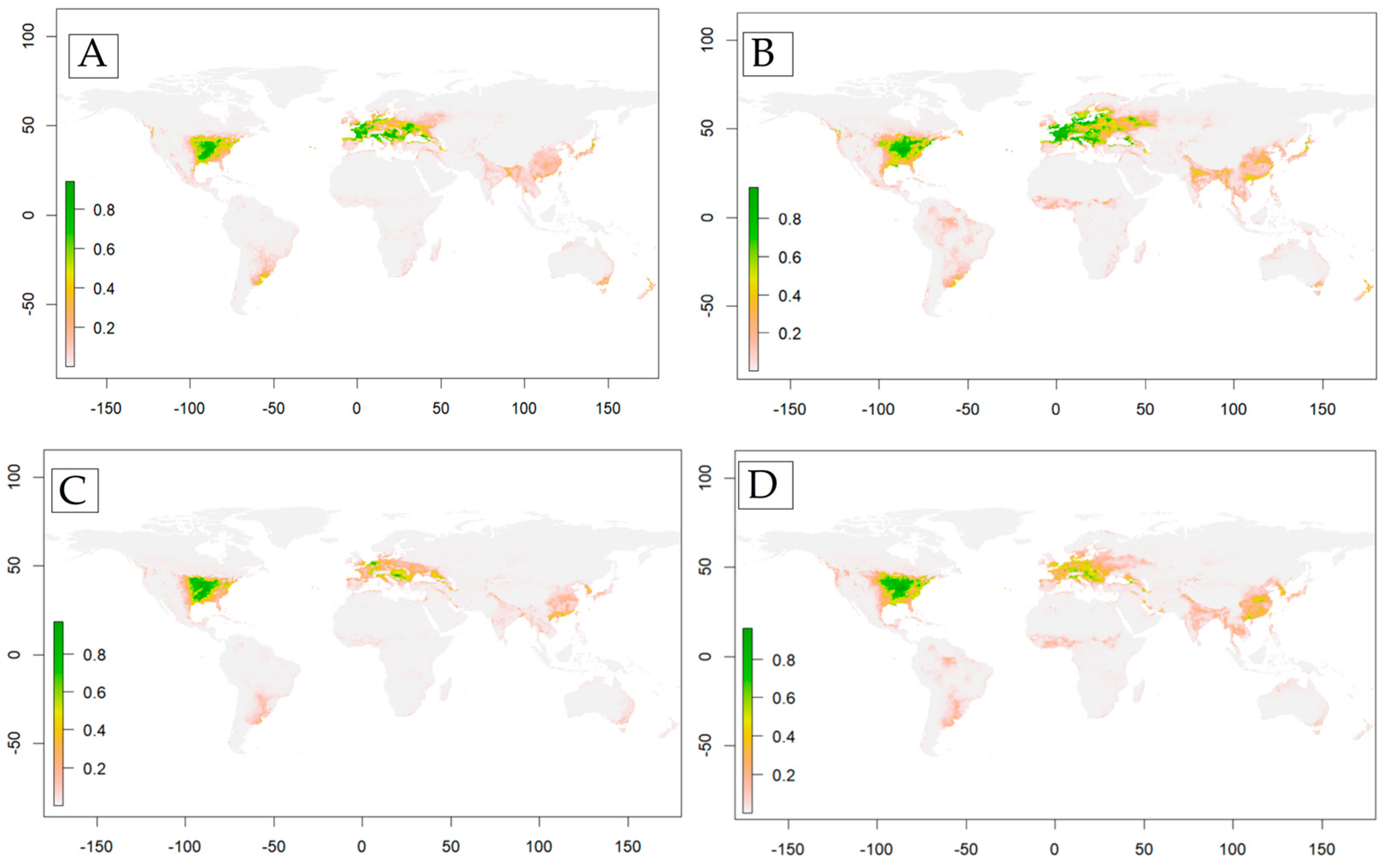
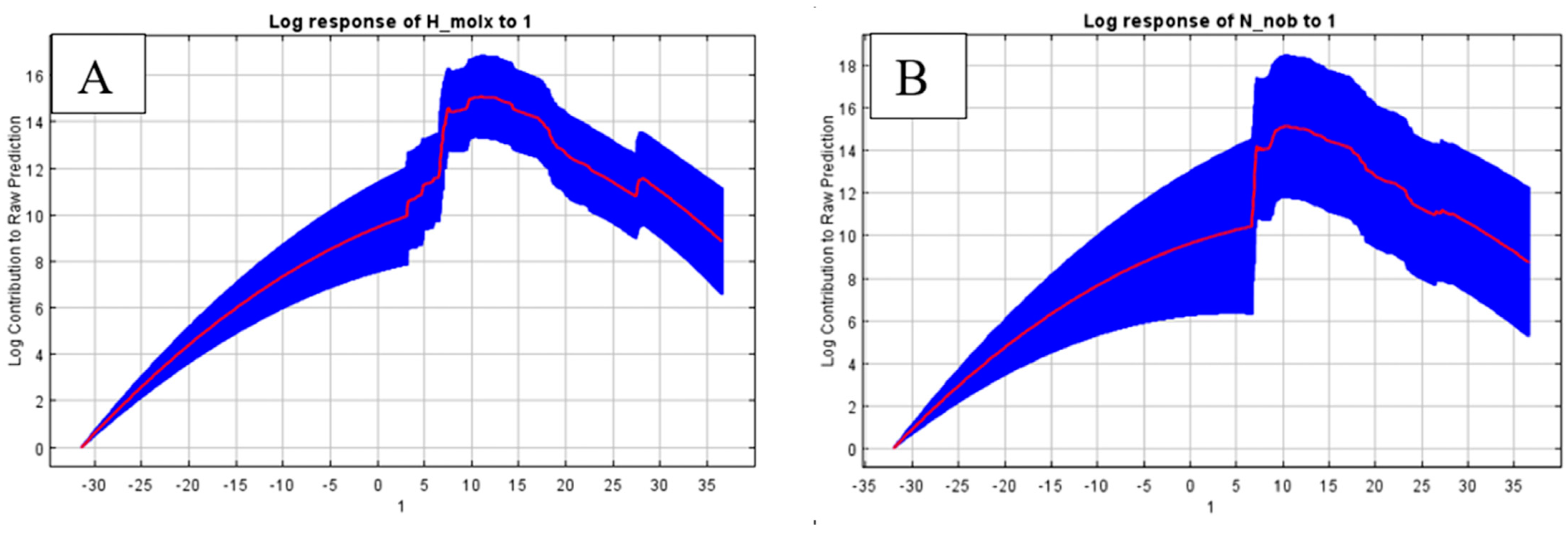
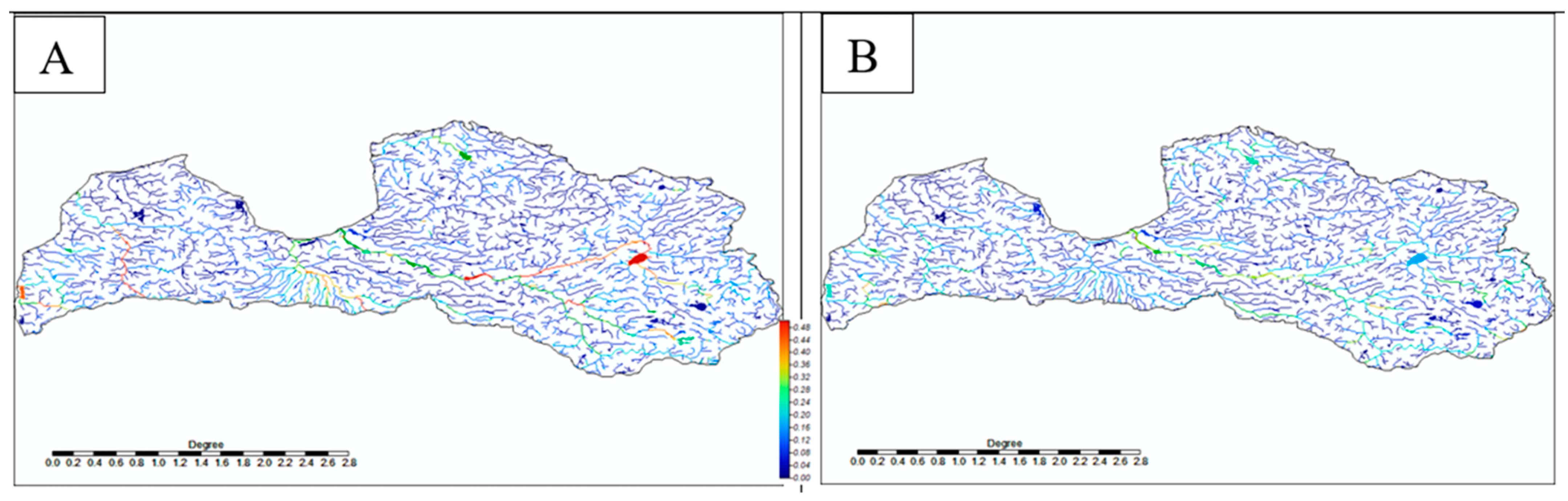
3. Results
4. Discussion
5. Conclusions
- Thorough impact assessments: Comprehensive evaluations of the potential ecological consequences of Asian carp species occurrence in Northern European ecosystems are essential (including the introduction of health-threatening infections and parasites).
- Monitoring and management strategies: Effective monitoring and management strategies are critical for mitigating any negative impacts on native fauna and ensuring the sustainable development of aquaculture practices.
- Collaborative approach: Collaboration among scientists, policymakers, and stakeholders is paramount to developing sustainable aquaculture practices that balance food production with the preservation of native biodiversity.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liao, I.C.; Chao, N.H. Aquaculture and food crisis: Opportunities and constraints. Asia Pac. J. Clin. Nutr. 2009, 18, 564–569. [Google Scholar] [PubMed]
- Neori, A.; Shpige, M.; Guttman, L.; Israel, A. Development of polyculture and integrated multi-trophic aquaculture (IMTA) in Israel: A review. Isr. J. Aquac.-Bamidgeh 2017, 69, 1–19. [Google Scholar] [CrossRef]
- Rakocy, J.E. Aquaponics—Integrating fish and plant culture. In Aquaculture Production Systems; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 344–386. [Google Scholar] [CrossRef]
- Billard, R.; Berni, P. Trends in cyprinid polyculture. Cybium 2004, 28, 255–261. [Google Scholar]
- Jeney, Z.; Jian, Z. Use and exchange of aquatic resources relevant for food and aquaculture: Common carp (Cyprinus carpio L.). Rev. Aquac. 2009, 1, 163–173. [Google Scholar]
- Woynarovich, A.; Bueno, P.B.; Altan, O.; Jeney, Z.; Reantaso, M.; Xinhua, Y.; Van Anrooy, R. Better management practices for carp production in Central and Eastern Europe, the Caucasus and Central Asia; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Szücs, I.; Stündl, L.; Váradi, L. Carp farming in Central and Eastern Europe and a case study in multifunctional aquaculture. In Species and System Selection for Sustainable Aquaculture; Springer: Berlin/Heidelberg, Germany, 2007; pp. 389–414. [Google Scholar]
- Kuybida, V.V.; Nekrasova, O.D.; Kutsokon, Y.K.; Lopatynska, V.V. Summer fish kills in the Kaniv Reservoir. Hydrobiol. J. 2019, 55, 103–106. [Google Scholar] [CrossRef]
- Nekrasova, O.D.; Tytar, V.M.; Kuybida, V.V. GIS Modeling of Climate Change Vulnerability of Amphibians and Reptiles in Ukraine; NAS of Ukraine, Shmalgausen Institute of Zoology NAS: Kyiv, Ukraine, 2019; pp. 1–204. ISBN 978-966-02-8956-7. (In Ukrainian)
- Nekrasova, O.; Tytar, V.; Pupins, M.; Čeirāns, A.; Marushchak, O.; Skute, A. GIS Modeling Study of the Distribution of Viviparous Invasive Alien Fish Species in Eastern Europe in Terms of Global Climate Change, as Exemplified by Poecilia reticulata Peters, 1859 and Gambusia holbrooki Girarg, 1859. Diversity 2021, 13, 385. [Google Scholar] [CrossRef]
- Tytar, V.; Nekrasova, O.; Pupins, M.; Čeirāns, A.; Skute, A. Modelling the range expansion of pumpkinseed Lepomis gibbosus across Europe, with a special focus on Ukraine and Latvia. North-West. J. Zool. 2022, 18, 143–150. [Google Scholar]
- Georges, J.-Y.; Pupins, M.; Nekrasova, O.; Marushchak, O.; Tytar, V.; Skute, A.; Ceirans, A.; Petrov, I.; Garkajs, A.; Theissinger, K. Distribution trends of invasive Asian carps (Hypophthalmichthys molitrix and Aristichthys nobilis) in Europe in the context of climate change. In Proceedings of the 36th Congress of the International Society of Limnology, Berlin, Germany, 7–10 August 2022; p. 335. [Google Scholar]
- Wang, J.; Lamer, J.T.; Gaughan, S.; Wachholtz, M.; Wang, C.; Lu, G. Transcriptomic comparison of invasive bigheaded carps (Hypophthalmichthys nobilis and Hypophthalmichthys molitrix) and their hybrids. Ecol. Evol. 2016, 6, 8452–8459. [Google Scholar] [CrossRef]
- Karpova, E.P.; Boltachev, A.R. Fish of the Inner Water Reservoirs in the Crimean Peninsula; Business-Inform: Simferopol, Crimea, 2012; p. 200. [Google Scholar]
- Ayles, G.B.; Campbell, K.; Gillis, D.; Saunders, L.; Scott, K.J.; Tallman, R.; Traverse, N. Technical assessment of the status, health and sustainable harvest levels of the lake Winnipeg fisheries resource. In Lake Winnipeg Quota Review Task Force Report, 2nd ed.; Conover, G., Simmonds, R., Whalen, M., Eds.; Asian Carp Working Group, Aquatic Nuisance Species Task Force: Washington, DC, USA, 2011; Volume 3, pp. 154–196. [Google Scholar]
- Zambrano, L.; Martínez-Meyer, E.; Menezes, N.; Peterson, A.T. Invasive potential of common carp (Cyprinus carpio) and Nile tilapia (Oreochromis niloticus) in American freshwater systems. Can. J. Fish. Aquat. Sci. 2006, 63, 1903–1910. [Google Scholar] [CrossRef]
- Milardi, M.; Chapman, D.; Lanzoni, M.; Long, J.M.; Castaldelli, G. First evidence of bighead carp wild recruitment in Western Europe, and its relation to hydrology and temperature. PLoS ONE 2017, 12, e0189517. [Google Scholar] [CrossRef]
- Jawdhari, A.; Mihăilescu, D.F.; Fendrihan, S.; Jujea, V.; Stoilov-Linu, V.; Negrea, B.M. Silver Carp (Hypophthalmichthys molitrix) (Asian Silver Carp) Presence in Danube Delta and Romania—A Review with Data on Natural Reproduction. Life 2022, 12, 1582. [Google Scholar] [CrossRef]
- Conover, G.; Simmonds, R.; Whalen, M. Management and control plan for bighead, black, grass, and silver carps in the United States. In Asian Carp Working Group, Aquatic Nuisance Species Task Force, Eds.; Asian Carp Working Group, Aquatic Nuisance Species Task Force: Washington, DC, USA, 2007; Volume 3, pp. 154–196. [Google Scholar]
- Novitskiy, R.A. Scale, Direction and Consequences of Alien Fish Species Invasions in the Dnieper Reservoirs: Avt. Ph.D. Thesis, Dnipro State Agrarian and Economic University, Kyiv, Ukraine, 2019; p. 41. [Google Scholar]
- Ciolac, A. Migration of fishes in Romanian Danube River. Appl. Ecol. Environ. Res. 2004, 2, 143–163. [Google Scholar] [CrossRef]
- Molnár, T.; Lehoczky, I.; Edviné Meleg, E.; Boros, G.; Specziár, A.; Mozsár, A.; Vitál, Z.; Józsa, V.; Allele, W.; Urbányi, B.; et al. Comparison of the Genetic Structure of Invasive Bigheaded Carp (Hypophthalmichthys spp.) Populations in Central-European Lacustrine and Riverine Habitats. Animals 2021, 11, 2018. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.C.E.; Knowler, D.; Kyobe, D.; de la Cueva Bueno, P. Preliminary Damage Estimates for Selected Invasive Fauna in B.C.; Report Prepared for Ecosystems Branch; B.C. Ministry of Environment, Victoria, B.C. by ESSA Technologies Ltd.: Vancouver, BC, USA, 2013; p. 62.
- Pupins, M.; Nekrasova, O.; Marushchak, O.; Tytar, V.; Theissinger, K.; Čeirāns, A.; Skute, A.; Georges, J.-Y. Potential Threat of an Invasive Fish Species for Two Native Newts Inhabiting Wetlands of Europe Vulnerable to Climate Change. Diversity 2023, 15, 201. [Google Scholar] [CrossRef]
- Soes, D.M.; Leuven, R.S.E.W.; Matthews, J.; Broeckx, P.B.; Haenen, O.L.M.; Engelsma, M.Y. A risk analysis of bigheaded carp (Hypophthalmichthys sp.) in the Netherlands. 2011. 118p. Available online: https://repository.ubn.ru.nl/handle/2066/123477 (accessed on 20 April 2024).
- DeVaney, S.C.; McNyset, K.M.; Williams, J.B.; Peterson, A.T.; Wiley, E.O. A tale of four “carp”: Invasion potential and ecological niche modeling. PLoS ONE 2009, 4, e5451. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wiley, E.O.; Mcnyset, K.M. Ecological niche modeling as a predictive tool: Silver and bighead carps in North America. Biol. Invasions 2007, 9, 43–51. [Google Scholar] [CrossRef]
- Kulhanek, S.A.; Leung, B.; Ricciardi, A. Using ecological niche models to predict the abundance and impact of invasive species: Application to the common carp. Ecol. Appl. 2011, 21, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Lübcker, N.; Zengeya, T.A.; Dabrowski, J.; Robertson, M.P. Predicting the potential distribution of invasive silver carp Hypophthalmichthys molitrix in South Africa. Afr. J. Aquat. Sci. 2014, 39, 157–165. [Google Scholar] [CrossRef]
- Smith, A. Bigheaded carps (Hypophthalmichthys molitrix and H. nobilis) An Annotated Bibliography on Literature Composed from 1970 to 2014. Publication 2890. Mississippi State University Extension, 2015. 43 p. Available online: https://extension.msstate.edu/sites/default/files/publications/publications/p2890.pdf (accessed on 4 September 2023).
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; p. 35. [Google Scholar]
- Historical Aquaculture in Northern Europe. Historical Aquaculture in Northern Europe, Eds. Madeleine Bonow, Håkan Olsén and Ingvar Svanberg; Elanders: Stockholm, Sweden, 2016; p. 204.
- Aristichthys nobilis (Richardson, 1846) (=Hypophthalmichthys nobilis) GBIF.org. Available online: https://www.gbif.org/occurrence/download/0097149-210914110416597 (accessed on 7 January 2022).
- Hypophthalmichthys molitrix (Valenciennes, 1844) GBIF.org. Available online: https://www.gbif.org/occurrence/download/0097129-210914110416597 (accessed on 7 January 2022).
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Peterson, A.T.; Falconi, M.; Contreras-Díaz, R.G. ntbox: An r package with graphical user interface for modelling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Jarošik, V.; Ota, N. Extending the suite of Bioclim variables: A proposed registry system and case study using principal components analysis. Methods Ecol. Evol. 2014, 5, 956–960. [Google Scholar] [CrossRef]
- Domisch, S.; Amatulli, G.; Jetz, W. Near-global freshwater-specific environmental variables for biodiversity analyses in 1 km resolution. Sci. Data 2015, 2, 150073. [Google Scholar] [CrossRef]
- Trabucco, A.; Zomer, R. Global Aridity Index and Potential Evapotranspiration (ET0) Climate Database v2. Figshare. Fileset. Available online: https://figshare.com/articles/dataset/Global_Aridity_Index_and_Potential_Evapotranspiration_ET0_Climate_Database_v2/7504448 (accessed on 4 September 2023).
- Phillips, S.J. A brief tutorial on Maxent. ATT Res. 2005, 190, 231–259. [Google Scholar]
- Peterson, A.T.; Papes, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating predictive models of species’ distributions: Criteria for selecting optimal models. Ecol. Model. 2003, 162, 211–232. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 29 February 2024).
- Rubenstein, M.A.; Weiskopf, S.R.; Bertrand, R.; Carter, S.L.; Comte, L.; Eaton, M.J.; Thompson, L.M. Climate change and the global redistribution of biodiversity: Substantial variation in empirical support for expected range shifts. Environ. Evid. 2023, 12, 1–21. [Google Scholar] [CrossRef]
- Colwell, R.K.; Thiago, F. Rangel. Hutchinson’s duality: The once and future niche. Proc. Natl. Acad. Sci. USA 2009, 106, 19651–19658. [Google Scholar] [CrossRef]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thiuller, W.; Peterson, A.T.; Giusan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Contribution Percent | Permutation Importance |
|---|---|---|
| H. molitrix | ||
| CliMond (Table S1) | ||
| bio1—Annual mean temperature (°C) | 22.0 | 26.6 |
| bio17—Precipitation of driest quarter (mm) | 19.2 | 1.2 |
| bio3—Isothermality | 14.1 | 6.3 |
| EarthEnv NGEI (Table S2) | ||
| lc_avg_09—Urban/built-up | 34.4 | 10.0 |
| lc_avg_04—Mixed/other trees | 12.1 | 18.1 |
| A. nobilis | ||
| CliMond (Table S1) | ||
| bio1—Annual mean temperature (°C) | 24.0 | 44.0 |
| bio3—Isothermality | 18.3 | 8.8 |
| bio30—Lowest weekly moisture index | 9.5 | 0.4 |
| EarthEnv NGEI (Table S2) | ||
| lc_avg_09—Urban/built-up | 29.1 | 11.8 |
| lc_avg_07—Cultivated and managed vegetation | 16.4 | 14.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekrasova, O.; Pupins, M.; Tytar, V.; Fedorenko, L.; Potrokhov, O.; Škute, A.; Čeirāns, A.; Theissinger, K.; Georges, J.-Y. Assessing Prospects of Integrating Asian Carp Polyculture in Europe: A Nature-Based Solution under Climate Change? Fishes 2024, 9, 148. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes9040148
Nekrasova O, Pupins M, Tytar V, Fedorenko L, Potrokhov O, Škute A, Čeirāns A, Theissinger K, Georges J-Y. Assessing Prospects of Integrating Asian Carp Polyculture in Europe: A Nature-Based Solution under Climate Change? Fishes. 2024; 9(4):148. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes9040148
Chicago/Turabian StyleNekrasova, Oksana, Mihails Pupins, Volodymyr Tytar, Leonid Fedorenko, Oleksandr Potrokhov, Arturs Škute, Andris Čeirāns, Kathrin Theissinger, and Jean-Yves Georges. 2024. "Assessing Prospects of Integrating Asian Carp Polyculture in Europe: A Nature-Based Solution under Climate Change?" Fishes 9, no. 4: 148. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes9040148