
Grafting Genotype Combination Effect of Vitis ssp. on Roots Phylloxeration Degree and Vigor of Grapevine
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Site
2.2. Experiment Set Up
2.3. Sampling and Measurements
2.4. Grape Phylloxera Populations
2.5. Statistical Analysis
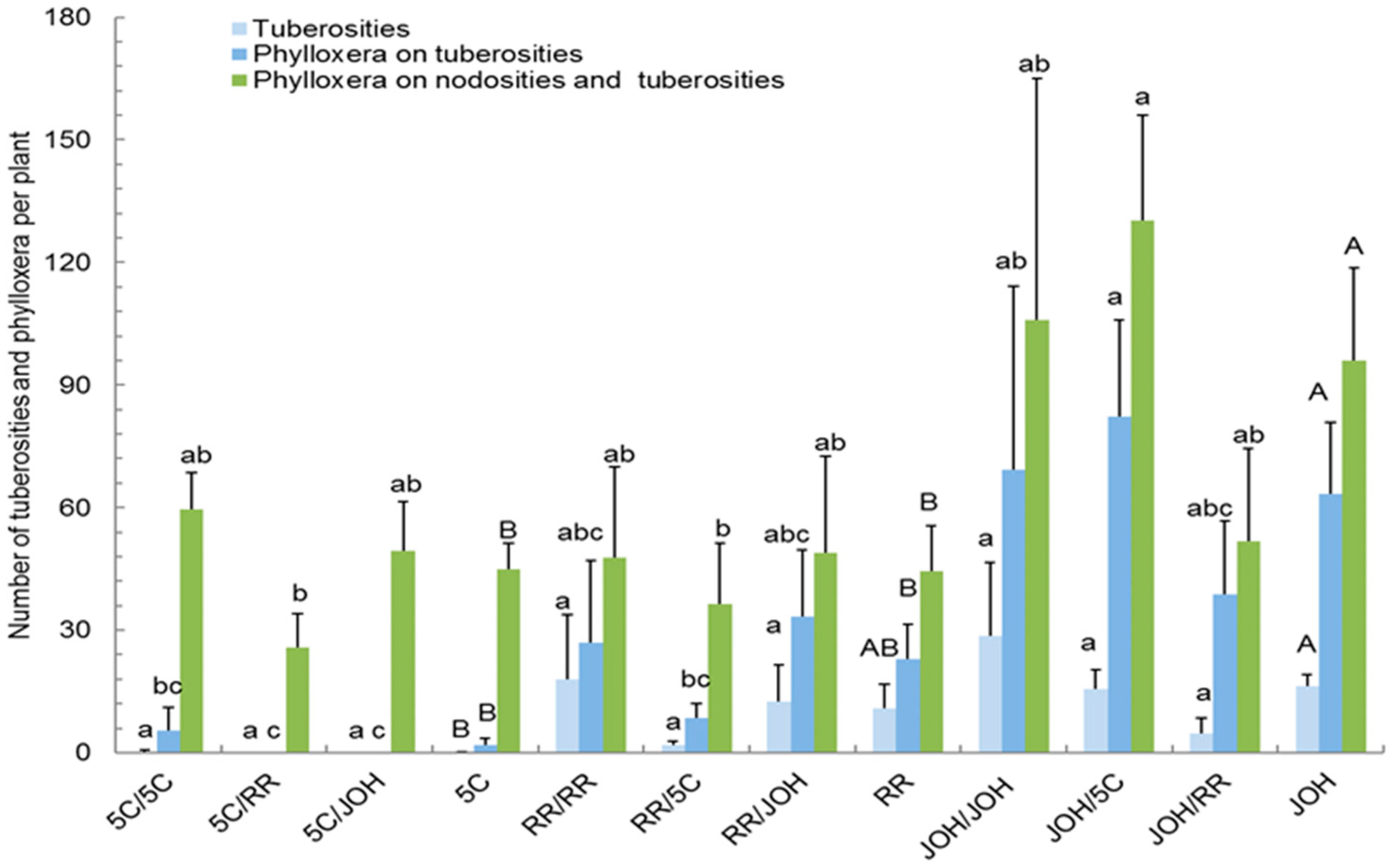
3. Results
3.1. Grape Phylloxera Infestation Level
3.2. Vigor of Scion-Graft Combinations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Granett, J.; Timper, P.; Lider, L.A. Grape phylloxera (Daktulosphaira vitifoliae) (Homoptera: Phylloxeridae) biotypes in California. J. Econom. Entomol. 1985, 78, 1463–1467. [Google Scholar] [CrossRef]
- Benheim, D.; Rochfort, S.; Robertson, E.; Potter, I.D.; Powell, K.S. Grape phylloxera (Daktulosphaira vitifoliae)—A review of potential detection and alternative management options. Ann. Appl. Biol. 2012, 161, 91–115. [Google Scholar] [CrossRef]
- Hofmann, E. Die Histologie der Nodositäten verschiedener Rebensorten bei Reblausbefall. Vitis 1957, 1, 66–81. [Google Scholar]
- Niklowitz, W. Histologische Studien an Reblausgallen und Reblausabwehrnekrosen (Viteus vulpinae CB. Auf Vitis vinifera und Vitis riparia). Phytopath. Z. 1955, 24, 299–340. [Google Scholar]
- Griesser, M.; Lawo, N.C.; Crespo-Martinez, S.; Schoedl-Hummel, K.; Wieczorek, K.; Gorecka, M.; Liebner, F.; Zweckmair, T.; Pavese, N.S.; Kreil, D.; et al. Phylloxera (Daktulosphaira vitifoliae Fitch) alters the carbohydrate metabolism in root galls to allowing the compatible interaction with grapevine (Vitis ssp.) roots. Plant. Sci. 2015, 234, 38–49. [Google Scholar] [CrossRef]
- Kellow, A.V.; Sedgley, M.; Van Heeswijck, R. Interaction between Vitis vinifera and grape phylloxera: Changes in root tissue during nodosity formation. Ann. Bot. 2004, 93, 581–590. [Google Scholar] [CrossRef]
- Savi, T.; García González, A.; Carlos Herrera, J.; Forneck, A. Gas exchange, biomass and non-structural carbohydrates dynamics in vines under combined drought and biotic stress. BMC Plant Biol. 2019, 19, 408. [Google Scholar] [CrossRef]
- Body, M.J.A.; Appel, H.M.; Edger, P.P.; Schultz, J.C. A gall-forming insect manipulates hostplant phytohormone synthesis, concentrations, and signaling. bioRxiv 2019. [Google Scholar] [CrossRef]
- Nabity, P.D.; Haus, M.J.; Berenbaum, M.R.; DeLucia, E.H. Leaf-galling phylloxera on grapes reprograms host metabolism and morphology. Proc. Natl. Acad. Sci. USA 2013, 110, 16663–16668. [Google Scholar] [CrossRef]
- Du, Y.; Zheng, Q.; Zhai, H.; Jiang, E.; Wang, Z. Selectivity of Phylloxera viticola Fitch (Homoptera: Phylloxeridae) to grape with different resistance and the identification of grape root volatiles. Acta Entomol. Sin. 2009, 52, 537–543. [Google Scholar]
- Schultz, J.C.; Edger, P.P.; Body, M.J.A.; Appel, H.M. A galling insect activates plant reproductive programs during gall development. Sci. Rep. 2019, 9, 1833. [Google Scholar] [CrossRef]
- Idris, I.; Arabi, M.I.E. The relationship between grape phylloxera and Fusarium root infection. Adv. Hortic. Sci. 2014, 28, 14–19. [Google Scholar]
- Walker, M.A.; Lund, K.; Agüero, C.; Riaz, S.; Fort, K.; Heinitz, C.; Romero, N. Breeding grape rootstocks for resistance to phylloxera and nematodes—It’s not always easy. Acta Hortic. 2014, 1045, 89–97. [Google Scholar] [CrossRef]
- Schmid, J.; Manty, F.; Rühl, E.H. Utilizing the complete phylloxera resistance of Vitis cinerea Arnold in rootstock breeding. Acta Hortic. 2003, 603, 393–400. [Google Scholar] [CrossRef]
- Zhang, J.; Hausmann, L.; Eibach, R.; Welter, L.J.; Töpfer, T.; Zyprian, E.M. A framework map from grapevine V3125 (Vitis vinifera ‘Schiava grossa’ x ‘Riesling’) x rootstock cultivar ‘Börner’ (Vitis riparia × Vitis cinerea) to localize genetic determinants of phylloxera root resistance. Theor. Appl. Genet. 2009, 119, 1039–1051. [Google Scholar] [CrossRef]
- Smith, H.M.; Clarke, C.W.; Smith, B.P.; Carmody, B.M.; Thomas, M.R.; Clingeleffer, P.R.; Powell, K.S. Genetic identification of SNP markers linked to a new grape phylloxera resistant locus in Vitis cinerea for marker-assisted selection. BMC Plant Biol. 2018, 18, 360. [Google Scholar] [CrossRef]
- Rubio, B.; Lalanne-Tisné, G.; Voisin, R.; Tandonnet, J.P.; Portier, U.; Van Ghelder, C.; Lafargue, M.; Petit, J.P.; Donnart, M.; Joubard, B.; et al. Characterization of genetic determinants of the resistance to phylloxera, Daktulosphaira vitifoliae, and the dagger nematode Xiphinema index from muscadine background. BMC Plant Biol. 2020, 20, 213. [Google Scholar] [CrossRef]
- Dietrich, A.; Wolf, T.; Eimert, K.; Schröder, M.B. Activation of gene expression during hypersensitive response (HR) induced by auxin in the grapevine rootstock cultivar ‘Börner’. Vitis 2010, 49, 15–21. [Google Scholar]
- Pavloušek, P. Screening of rootstock hybrids with Vitis cinerea Arnold for phylloxera resistance. Cent. Eur. J. Biol. 2012, 7, 708–719. [Google Scholar] [CrossRef]
- Qing, Z.; Yuanpeng, D.; Yan, D.; Heng, H. Changes of secondary metabolites content and phenolics metabolism-associated enzymes in phylloxera infested nodosities. Acta Hortic. 2011, 904, 67–76. [Google Scholar] [CrossRef]
- Clarke, C.W.; Powell, K.S.; Norng, S.; Carmody, B.M.; Walpole, M.; Cunningham, J.P. Performance of a Leaf-Galling Phylloxera (Daktulosphaira vitifoliae) on Roots of Diverse Vitis spp. Rootstocks in North East Victoria, Australia. Aust. J. Grape Wine Res. 2023, 2023, 1328258. [Google Scholar] [CrossRef]
- Rispe, C.; Legeai, F.; Nabity, P.D.; Fernández, R.; Arora, A.K.; Baa-Puyoulet, P.; Banfill, C.R.; Bao, L.; Barberà, M.; Bouallègue, M.; et al. The genome sequence of the grape phylloxera provides insights into the evolution, adaptation, and invasion routes of an iconic pest. BMC Plant Biol. 2020, 18, 90. [Google Scholar] [CrossRef]
- Forneck, A.; Mammerler, R.; Tello, J.; Breuer, M.; Müller, J.; Fahrentrapp, J. First European leaf-feeding grape phylloxera (Daktulosphaira vitifoliae Fitch) survey in Swiss and German commercial vineyards. Eur. J. Plant Pathol. 2019, 154, 1029–1039. [Google Scholar] [CrossRef]
- Tandonnet, J.P.; Cookson, S.J.; Vivin, P.; Ollat, N. Scion genotype controls biomass allocation and root development in grafted grapevine. Aust. J. Grape Wine Res. 2010, 16, 290–300. [Google Scholar] [CrossRef]
- Granett, J.; Walker, M.A.; Fossen, M.A. Association between grape phylloxera and strongly resistant rootstocks in California: Bioassays. Acta Hortic. 2007, 733, 25–31. [Google Scholar] [CrossRef]
- Clarke, C.W.; Henneken, J.; Carmody, B.M.; Cunningham, J.P. Performance of Six Genetically Diverse Phylloxera Strains on 5C Teleki (V. berlandieri × V. riparia) Rootstock. Aust. J. Grape Wine Res. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- Morlat, R.; Jacquet, A. The soil effects on the grapevine root system in several vineyards of the Loire valley (France). Vitis 1993, 32, 35–42. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Hypobiont | Epibiont | ||
|---|---|---|---|
| Teleki 5C | Riesling | Johanniter | |
| Teleki 5C | 5C × 5C | 5C × RR | 5C × JOH |
| Johanniter | JOH × 5C | JOH × RR | JOH × JOH |
| Riesling | RR × 5C | RR × RR | RR × JOH |
| Combination Hypobiont/Epibiont | Nr. of Nodosities (Average ± SE) | Nr. of Individuals (Average ± SE) | ||||||
|---|---|---|---|---|---|---|---|---|
| VEM | GF | VEM | GF | |||||
| 5C/5C | 273.5 ± 78.10 | a | 161.50 ± 26.35 | a | 60.7 ± 21.24 | a | 54 ± 7.39 | a |
| 5C/RR | 195.7 ± 34.85 | ab | 35.50 ± 16.46 | bc | 31.2 ± 13.30 | ab | 25.7 ± 8.39 | abc |
| 5C/JOH | 107.5 ± 36.36 | bc | 52.83 ± 8.02 | b | 30.3 ± 10.24 | ab | 49.3 ± 12.13 | ab |
| 5C | 192.2 ± 33.42 | A | 83.28 ± 16.85 | A | 40.7 ± 9.14 | A | 43.0 ± 5.98 | A |
| RR/RR | 7.5 ± 2.10 | c | 44.50 ± 12.29 | bc | 0.75± 0.48 | b | 20.7 ± 4.42 | bc |
| RR/5C | 2.0 ± 2.00 | c | 27.83 ± 13.89 | bc | 0.25 ± 0.25 | b | 28.0 ± 11.94 | abc |
| RR/JOH | 52.0 ± 14.59 | c | 6.33 ± 4.22 | c | 19.0 ± 6.76 | ab | 15.7 ± 7.75 | c |
| RR | 25.0 ± 8.83 | B | 26.22 ± 7.06 | B | 8.4 ± 3.74 | B | 21.4 ± 4.82 | B |
| JOH/JOH | 53.8 ± 49.19 | c | 22.17 ± 9.75 | bc | 21.40 ± 19.70 | ab | 8.5 ± 7.58 | c |
| JOH/5C | 1.0 ± 0.58 | c | 40.00 ± 4.56 | bc | 0.25 ± 0.25 | b | 40.0 ± 4.35 | ab |
| JOH/RR | 15.2 ± 7.50 | c | 8.50 ± 5.98 | c | 6.25 ± 2.90 | b | 22.2 ± 14.64 | abc |
| JOH | 25.69 ± 18.93 | B | 23.56 ± 4.96 | B | 10.23 ± 7.57 | B | 32.5 ± 6.30 | AB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vršič, S.; Gumzej, M.; Pulko, B.; Kocsis, L.; Eitle, M.W.; Forneck, A. Grafting Genotype Combination Effect of Vitis ssp. on Roots Phylloxeration Degree and Vigor of Grapevine. Horticulturae 2024, 10, 445. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050445
Vršič S, Gumzej M, Pulko B, Kocsis L, Eitle MW, Forneck A. Grafting Genotype Combination Effect of Vitis ssp. on Roots Phylloxeration Degree and Vigor of Grapevine. Horticulturae. 2024; 10(5):445. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050445
Chicago/Turabian StyleVršič, Stanko, Mojca Gumzej, Borut Pulko, László Kocsis, Markus W. Eitle, and Astrid Forneck. 2024. "Grafting Genotype Combination Effect of Vitis ssp. on Roots Phylloxeration Degree and Vigor of Grapevine" Horticulturae 10, no. 5: 445. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae10050445