1. Introduction
Bacteriocins, including lantibiotics, are ribosomally synthesized peptides that exhibit antimicrobial activity against a wide range of human and animal pathogens [
1]. Due to their potential as alternatives to currently used antibiotics in medical applications and as natural preservatives in food processing, bacteriocins have gained considerable interest [
2,
3]. Quantifying bacteriocins is crucial in their discovery, purification, and production optimization. Bioassays, based on inhibiting the growth of sensitive indicator microorganisms, are commonly used for the detection and quantification of bacteriocins. Despite the utility of these bioassays in the discovery of numerous bacteriocins over the years [
4,
5], they present several challenges. These challenges include failure to detect very low concentrations of bacteriocins, inability to differentiate among co-produced antimicrobial molecules, and the dependence of the assay’s outcome on the indicator’s growth phase and physiological status [
6,
7]. To overcome these shortcomings, immunoassays have been developed for detecting and quantifying a few bacteriocins, such as nisin, pediocin, lacticin RM, and propionicin PLG-1 [
8,
9,
10,
11].
In an enzyme-linked immunosorbent assay (ELISA), antibodies are used to detect and quantify a specific molecule (e.g., a bacteriocin) in a sample. Specific antibodies must be generated and then utilized to detect and quantify the target bacteriocin peptide within a complex biological sample [
8,
12]. Compared with traditional bioassays, immunoassays can distinguish, with high specificity, among different types of bacteriocins in a complex matrix, allowing for the detection and quantification of the targeted peptide only. Furthermore, immunoassays are also highly sensitive, enabling the detection and quantification of bacteriocins at concentrations that are too low to be detected by bioassays [
10]. In addition to specificity and sensitivity, ELISAs are also rapid and cost-effective. ELISA-based methods and associated polyclonal antibodies have been successfully developed for the quantitative determination of several bacteriocins, including nisin, propionicin PLG-1, C-terminal amino acid sequence of hiracin JM79, and enterocin P [
8,
10,
13,
14].
Although many bacteriocins have been discovered lately, including paenibacillin, very few of these have been implemented industrially. One of the major implementation hurdles is the lack of accurate and specific methods to quantify these bacteriocins in their ready-to-use preparations or after addition to food. In the current study, polyclonal antibodies were generated, and a competitive ELISA was subsequently developed for detecting and quantifying paenibacillin, a novel lantibiotic bacteriocin. This bacteriocin was discovered in
Paenibacillus polymyxa, a commensal bacterium isolated from the fermented vegetable Kimchee [
4]. The molecular mass and primary structure of paenibacillin (2983.54 Da; CH
3CO-A-Dha-I-I-K-Dhb-Dhb-I-K-V-A(1)-K-A-V-A(1)-K-Abu(2)-L-Abu(3)-A(2)-I-A(3)-Abu(4)-G-A(5)-A(4)-Dha-N-A(5)-K) were elucidated in previous studies [
4,
15]. This bacteriocin is potent against a wide array of foodborne pathogens and spoilage microorganisms, including
Bacillus spp.,
Staphylococcus aureus,
Listeria spp., and
Clostridium sporogenes [
4]. Thus, developing an ELISA for paenibacillin quantification will facilitate optimizing its production, provide a versatile method for the discovery of unexplored paenibacillin-like peptides, and potentially quantifying paenibacillin variants produced by natural microbiota.
2. Materials and Methods
2.1. Bacterial Strains and Paenibacillin
The microbial peptide paenibacillin was obtained as a purified powder (more than 90% purity as determined by high-performance liquid chromatography (HPLC) (
Supplementary File S1) from the Food Microbiology Laboratory at The Ohio State University, Columbus, OH. Paenibacillin was produced by the bacteriocinogenic strain
P. polymyxa OSY-DF ATCC PTA-7852 (
P. polymyxa OSY-DF, wild type) and purified as described in a previous study [
4]. For testing the usefulness of the developed competitive ELISA in paenibacillin quantification, the peptide producers
P. polymyxa OSY-EC (mutant) and
P. polymyxa OSY-DF (wild type) were selected for this analysis. The mutant strain (OSY-EC) was developed in a previous study [
16] to produce paenibacillin at higher levels than those produced by the wild type (OSY-DF). Both
P. polymyxa strains were transferred from the frozen stock cultures, streaked onto tryptic soy agar (TSA; Becton Dickinson and Company, Franklin Lakes, NJ, USA), and incubated for 24 h at 37 °C before few colonies were transferred into tryptic soy broth (TSB; Becton Dickinson, and Company) and incubated at 37 °C for 24 h with shaking at 120 rpm. Subsequently, both strains were grown in TSB at 30 °C or 37 °C with shaking at 120 rpm until their cultures reached 1.2 optical density at 600 nm (OD
600). Crude paenibacillin-containing supernatants from both cultures were prepared by centrifugation and microfiltration as described in previous studies [
16,
17] before being used for competitive ELISA experiments. A conventional bioassay also was used to determine paenibacillin activity, using
Listeria innocua ATCC 33090 as an indicator strain [
4,
16,
17]. For comparing the transcription of the paenibacillin biosynthetic gene cluster in
P. polymyxa OSY-EC and
P. polymyxa OSY-DF, both strains were grown in TSB at 30 °C or 37 °C, with shaking at 120 rpm to reach an OD
600 of 1.2.
2.2. Anti-Paenibacillin Antiserum Production
Anti-paenibacillin antiserum production and the succeeding purification of anti-paenibacillin polyclonal antibodies (pAbs) were performed by Creative Diagnostics company (Shirley, NY, USA). Paenibacillin was coupled to keyhole limpet hemocyanin (KLH) for immunizing female New Zealand white rabbits. Paenibacillin was also coupled to bovine serum albumin (BSA) for developing the ELISA. Paenibacillin–KLH conjugate (0.5 mg), suspended in phosphate-buffered saline (PBS; pH 7.5), was mixed with complete Freund’s adjuvant, in a ratio of 1:1, and injected subcutaneously at ten different sites of the dorsal region of each rabbit. The second injection of the paenibacillin–KLH conjugate (0.5 mg) mixed with incomplete Freund’s adjuvant was given 14 days after the first injection. Three additional booster injections were given consecutively at 14, 28, and 42 days from the second injection. Twelve days after the third booster, anti-paenibacillin antisera were collected and tested by an indirect ELISA. Subsequently, anti-paenibacillin pAbs were purified from the raw antisera by using affinity chromatography if the titer was more than 1:8000.
2.3. Affinity Chromatography Purification
Anti-paenibacillin pAbs were purified by using an affinity chromatography column (Creative Diagnostics) equilibrated to room temperature before use and immobilized with purified paenibacillin to capture anti-paenibacillin IgG. The anti-paenibacillin antisera were clarified by centrifugation at 20,000× g and 4 °C for 20 min, and the supernatant was collected. Supernatants were diluted with a binding buffer (0.1 M phosphate buffer and 0.15 M NaCl; pH 8). To remove the residues of the storage buffer, an agarose affinity column was washed with the binding buffer (30% ethanol in water or 0.05% thimerosal in water) before adjusting the pH to 8.0 and adding the anti-paenibacillin antisera to the column at a rate of 0.5–1 mL/min. Protein absorbance was monitored at 280 nm by using an in-line UV monitor. Subsequently, the column was washed with the binding buffer at a flow rate of 1 mL/min to remove unbound proteins. The anti-paenibacillin immunoglobulins were eluted from the affinity column with 0.1 M citric acid, and 1 mL fractions were collected in tubes containing 1 M neutralization buffer (PBS at pH 8). The pooled fractions containing the anti-paenibacillin immunoglobulins were dialyzed against PBS and concentrated by using a low-molecular-weight cutoff membrane, according to the manufacturer’s instructions.
2.4. Competitive ELISA Development
The competitive ELISA is based on the competition between immobilized paenibacillin on the surfaces of microplate wells and the free paenibacillin in the sample for recognition by the anti-paenibacillin pAbs. The signal obtained is inversely proportional to the concentration of paenibacillin in the sample. For developing a competitive ELISA method, several parameters were optimized; these included determining the most effective blocker to yield the lowest background signal, determining the anti-paenibacillin antibody titer, and the construction of a standard curve for enabling the quantification of paenibacillin concentrations in unknown samples.
2.4.1. Evaluation of Different Blockers
The wells of flat-bottomed clear polystyrene microtiter plates (Catalogue number #9018; Corning, Tewksbury, MA, USA) were coated with 100 µL of paenibacillin–BSA conjugate (200 ng) in a coating buffer solution (CBS; 1.59 g Na2CO3 + 2.93 g NaHCO3 per liter, pH 9.6; Boca Scientific Inc., Dedham, MA, USA) and incubated overnight at 4 °C in a humid chamber. Based on preliminary experiments, the use of higher doses of paenibacillin–BSA conjugate (400 ng) for coating did not yield satisfactory results compared with 200 ng of the conjugate. After coating, the plates were washed five times with 200 µL of the wash buffer, PBS with 0.1% Tween 20 (PBST; Sigma Aldrich, St. Louis, MO, USA) at pH 7.4. The coated–washed plates were blocked for 2 h at 37 °C with one of three blockers in PBS (pH 7.4): (i) 3% BSA + 0.1% Tween 20, (ii) 10% normal goat serum (NGS; Thermo Fisher Scientific, Waltham, MA, USA), and (iii) 3% Tween 20. Subsequently, the blocking solution was removed, and the plates were washed four times with PBST. Next, anti-paenibacillin pAbs-free negative control rabbit serum (Creative Diagnostics) was serially diluted in the appropriate blocking solution at the following dilutions (1:1 k, 1:4 k, 1:8 k, 1:16 k, 1:32 k, 1:64 k, 1:128 k, 1:256 k, 1:512 k, and 1:1024 k), and 100 µL of these diluted sera was added to each well of the microtiter plate, followed by 1 h incubation at 37 °C. Blank wells received the appropriate blocking solution alone. After incubation, the unbound negative serum was removed by washing five times with 200 µL of PBST. Then, 100 µL of diluted biotinylated goat anti-rabbit immunoglobulin (VECTASTAIN® ABC Standard kit; Vector Laboratories, Newark, CA, USA) was added to each well and incubated at 37 °C for 1 h. After incubation, the plates were washed five times with PBST; this was followed by the addition of 100 µL of a complex of avidin and biotinylate horseradish peroxidase (HRP) (VECTASTAIN® ABC Standard kit) to each well and incubation at 37 °C for 30 min. After incubation, the plates were washed five times with PBST, and the bound HRP was detected by adding 100 µL of tetramethylbenzidine (TMB) substrate (1-step™ Turbo TMB-ELISA; Thermo Fisher Scientific) to each well, incubating at 37 °C for 3 min, and stopping the reaction with 50 µL of 2 M H2SO4 (R&D™ Systems Stop solution, Fisher Scientific, Fair Lawn, NJ, USA). The spectrophotometric absorbance of the signals was measured at 450 nm by using a microplate reader (SpectraMax; Molecular Devices, San Jose, CA, USA).
2.4.2. Determination of Anti-Paenibacillin Antibody Titer
The wells of the microtiter plate were coated overnight at 4 °C with 100 µL of paenibacillin–BSA conjugate (200 ng) in CBS as previously described in
Section 2.4.1. Following coating, the plates were washed and blocked with 10% NGS, due to its lowest background signals compared with BSA and Tween 20. Serially diluted negative control sera or purified anti-paenibacillin pAbs (dilutions: 1:1 k, 1:4 k, 1:8 k, 1:16 k, 1:32 k, 1:64 k, 1:128 k, 1:256 k, 1:512 k, and 1:1024 k), both suspended in 10% NGS blocking solution, were added to the wells (100 µL each) and incubated for 1 h at 37 °C. After incubation, the microtiter plates were washed and treated with biotinylated goat anti-rabbit IgG and the avidin–biotin–HRP complex, as described in the previous section. Detection of HRP was indicative of bound anti-paenibacillin pAbs. The spectrophotometric absorbance of the signals were measured at 450 nm after adding the TMB substrate, either undiluted or 1:1-diluted with water. The titer of purified anti-paenibacillin pAbs was determined as the highest dilution that produced an absorbance value at least twice that of the corresponding negative control serum dilution.
2.4.3. Competitive ELISA Standard Curve
The microtiter plates were coated and blocked with 10% NGS, as previously described in
Section 2.4.2. Subsequently, 100 µL of pure paenibacillin in PBS at varying concentrations was mixed with 100 µL of 1:1000 anti-paenibacillin pAbs in 10% NGS and incubated for 1 h at 37 °C. This was followed by the addition of 100 µL of the mixture, achieving final paenibacillin concentrations of 1.56, 6.2, 12.5, 25, 50, 100, and 200 ng/well, to the coated and blocked microtiter plates wells and incubation for 1 h at 37 °C. After washing, the bound anti-paenibacillin pAbs to the paenibacillin–BSA conjugate on the plate were determined after adding biotinylated anti-rabbit IgG, avidin–biotin–HRP complex, and the chromogenic reagent; the resulting signals were measured at 450 nm as described in the previous sections. The OD
450 values were plotted against the pure paenibacillin concentrations per well, and data were fitted to different nonlinear models (GraphPad Prism 9.4.1.; San Diego, CA, USA). The following three-parameter nonlinear regression model provided the best representation of the data, and it was used to build the standard curve:
where
X is the concentration of paenibacillin;
Y is the OD
450 value;
A and
B are the minimum and maximum curve asymptotes, respectively; and
C is the concentration at which the response is half-way between
A and
B.
2.4.4. Determination of Paenibacillin in Unknown Samples
Non-diluted paenibacillin-containing crude supernatants from
P. polymyxa OSY-DF and
P. polymyxa OSY-EC were analyzed by using the competitive ELISA described in
Section 2.4.3, except that pure paenibacillin was replaced with crude supernatants. The standard curve equation was used to determine paenibacillin concentrations in the crude supernatants. Blank wells received the blocking solution, instead of anti-paenibacillin pAbs, and negative control wells included those with PBS or fresh TSB medium instead of standard paenibacillin solution or crude supernatants.
2.5. Recovery of Paenibacillin from Different Matrices by Using Competitive ELISA
Paenibacillin was dissolved in sterile PBS (pH 7.4), MRS, or diluted skim milk (prepared as 10% skim milk solution), yielding final concentrations of 2.5 and 10 µg/mL in each matrix. Paenibacillin recovery was assessed by using the competitive ELISA described in
Section 2.4.3. Each aliquot (100 µL) of these paenibacillin-containing preparations was mixed with 100 µL of anti-paenibacillin pAbs (1:1000 dilution) in 10% NGS blocking solution and incubated for 1 h at 37 °C. Then, 100 µL of the incubated mixture was added to the coated and blocked microtiter plate well. The bound anti-paenibacillin pAbs to the paenibacillin–BSA conjugate on the plate were determined as previously described. Paenibacillin recovery rates from MRS or 10% skim milk solution were compared to those from PBS as a control medium.
2.6. Conventional Bioassay for Paenibacillin Activity
Paenibacillin activity was determined according to a bacteriocin bioassay method [
18] with some modifications. Aliquots (10 µL) of pure paenibacillin, prepared at varying concentrations (0.15, 0.30, 0.6, 1.2, 2.5, 5, 10, and 20 µg per mL), were spotted onto TSA plates overlaid with soft TSA agar (0.75% agar), which was seeded with overnight culture of
L. innocua ATCC 33090. The spotted TSA plates were left undisturbed for 60 min to allow the agar medium to absorb the spotted dilution. The TSA plates were incubated for 24 h at 37 °C and observed for zones of inhibition to determine paenibacillin antimicrobial activity. The bioassay’s limit of detection (LOD) was determined as the lowest paenibacillin concentration that produced zones of inhibition on the spotted plates.
2.7. Gene Expression Analysis of Paenibacillin Biosynthetic Genes
P. polymyxa OSY-DF and OSY-EC strains were grown at 30 °C or 37 °C until both strains reached the same optical density as previously mentioned. Cells of both strains were harvested by centrifugation at 10,000×
g for 7 min, and the resulting cell pellets were used for RNA extraction as described in a previous study [
19]. Total RNA extraction, RNA cleanup, DNase treatment, cDNA synthesis, and cDNA utilization for relative expression of paenibacillin biosynthetic genes were performed as mentioned in the previous study by using reverse-transcription quantitative polymerase chain reaction (RT-qPCR; CFX96 Touch Real-Time PCR detection system; Bio-Rad, Hercules, CA, USA). The transcription levels of paenibacillin biosynthetic genes (
paeA,
paeP,
paeB,
paeC,
paeI,
paeT,
paeN,
agrB,
agrD,
agrA, and
agrC) and the reference gene,
16S rRNA, were measured in the mutant
P. polymyxa OSY-EC and compared with the wild type
P. polymyxa OSY-DF by using the 2
−ΔΔCt method [
20]. The primer sequences for paenibacillin biosynthetic genes were designed in the current study and used in the RT-qPCR (
Supplementary File S2).
2.8. Statistical Analysis
Each experiment was performed in duplicate and independently repeated twice, at least. Data are represented as the means ± SDs of the repeats. Analysis of variance (ANOVA) with Tukey’s/LSD pairwise comparisons was used to determine statistical significance among groups or pairwise comparisons, with a p-value of ≤0.05 being considered significant. Statistical analyses and standard curve construction were performed by using commercial statistical software (GraphPad Prism 9.4.1).
4. Discussion
Paenibacillin, a novel lantibiotic discovered in 2007 and purified from
P. polymyxa OSY-DF, exhibits potent bactericidal effects at low concentrations against a variety of Gram-positive pathogenic and spoilage bacteria [
4]. The potential use of paenibacillin in food preservation is promising, and it has been produced recently in an economical cheese acid whey-based medium [
17]. Given these advantages, paenibacillin could serve as an alternative to nisin, a natural food preservative that is currently facing challenges due to reduced antimicrobial efficacy caused by antimicrobial resistance [
21]. Developing a quantitative and sensitive method for detecting paenibacillin improves the feasibility of its application. The advantages of the paenibacillin competitive ELISA, developed in the current study, over the conventional bioassays are summarized in
Supplementary File S1.
In the current study, a reliable and efficient procedure for primary immunization of female New Zealand rabbits was developed to produce anti-paenibacillin pAbs. To ensure successful immunization and antibody production, paenibacillin was conjugated to KLH, which led to satisfactory levels of anti-paenibacillin pAbs in the collected raw antisera. Unconjugated paenibacillin was not used for rabbit immunization, since researchers have previously found that sole injection with some lantibiotics (e.g., nisin A, pediocin AcH, and pediocin RS2) without hapten conjugation did not yield satisfactory immunization outcomes in terms of antibody production [
9,
22,
23].
Subsequently, a competitive ELISA method for quantitative detection of paenibacillin in complex matrices was established. Optimizing the ELISA required the elimination of non-specific background signals by using a blocking agent, which non-specifically binds to assay well surfaces, thereby reducing the signal-to-noise ratio. It was concluded that NGS was the most effective blocker, compared with Tween 20 or BSA (
Figure 1). NGS contains a variety of proteins, including immunoglobulins and other serum proteins, which serve as blocking agents to prevent the non-specific binding of the detection antibodies to the assay well surfaces [
24,
25]. In contrast, BSA has only one protein as a blocking agent, which accounts for its lower blocking activity compared with NGS [
26]. Utilizing NGS as a blocking agent enabled the effective development of an ELISA in the current study.
A competitive ELISA was chosen for numerous advantages. This type of assay does not require the preprocessing of paenibacillin-containing samples (i.e., crude or impure extracts can be used directly) and is less sensitive to matrix inhibitory effects. Competitive ELISAs have been successfully developed for several antimicrobial peptides, such as polymyxin B and nisin [
12,
27,
28,
29,
30]. In the current study, a sigmoidal three-parameter nonlinear regression model was used to construct a competitive ELISA standard curve, which accurately correlated paenibacillin concentration with the OD
450 signals (
R2 = 0.95). The LOD of the competitive ELISA assay was significantly lower than that of the conventional bioassay; the latter is influenced by factors such as indicator growth status and the bacterial lawn’s thicknesses. The competitive ELISA detected concentrations as low as 15.6 ng/mL, which constitutes satisfactory sensitivity comparable to those reported in previous studies showing detection limits of 5 to 100 ng/mL for nisin A and 17.5 ng/mL for polymyxin B [
12,
27]. This low detection limit enables the paenibacillin-competitive ELISA assay to provide high sensitivity, detect small quantities of the peptide in natural food matrices, and potentially discover paenibacillin variants that are naturally produced in low quantities.
The use of conventional bioassays to determine the antimicrobial activity of paenibacillin-containing crude supernatants may not accurately differentiate between paenibacillin activity only and the synergistic activity resulting from paenibacillin, polymyxin and fusaricidin, which are naturally co-produced in the same microbiological medium [
16,
17]. This concern reduces the specificity of the paenibacillin bioassay, which can be addressed by using specific antibodies for paenibacillin. The competitive ELISA for paenibacillin, developed in the current study, displayed robust pAbs–paenibacillin interactions in crude supernatants from
P. polymyxa OSY-DF and
P. polymyxa OSY-EC (
Figure 5). Despite the complexity of these supernatants, the ELISA assay showed high specificity in capturing paenibacillin due to the strong affinity of these pAbs for the peptide. This observation is supported by previous studies reporting high specificity of competitive ELISA methods for detecting polymyxin B and nisin Z [
8,
12]. The current competitive ELISA had adequate repeatability. The percent coefficients of variation (%CV) for paenibacillin quantification in the crude supernatants of
P. polymyxa OSY-EC were 29.4% and 31.8% at 30 °C and 37 °C, respectively. For OSY-DF, the %CV values were 27.2% and 7.0% at the same temperatures, indicating the method’s reliability across varying conditions.
The specificity of immunoassays used for quantitative measurements of bacteriocins is advantageous [
31,
32,
33]. The developed competitive ELISA showed specificity as demonstrated by (i) the limited reactivity of the anti-paenibacillin pAbs with other antimicrobial peptides (nisin, vancomycin, and polymyxin), as shown in
Supplementary File S1, and (ii) its ability to distinguish paenibacillin levels produced by the mutant
P. polymyxa OSY-EC compared with the wild type
P. polymyxa OSY-DF strain. The mutant accumulated more paenibacillin than the wild type at both incubation temperatures (30 °C and 37 °C), consistent with a previous report that showed that the mutant produced a higher paenibacillin titer than the wild type as determined by a conventional bioassay [
16]. The competitive ELISA results for the mutant and wild type paenibacillin producers were further validated by the relative gene expression analysis of the paenibacillin biosynthetic gene cluster (
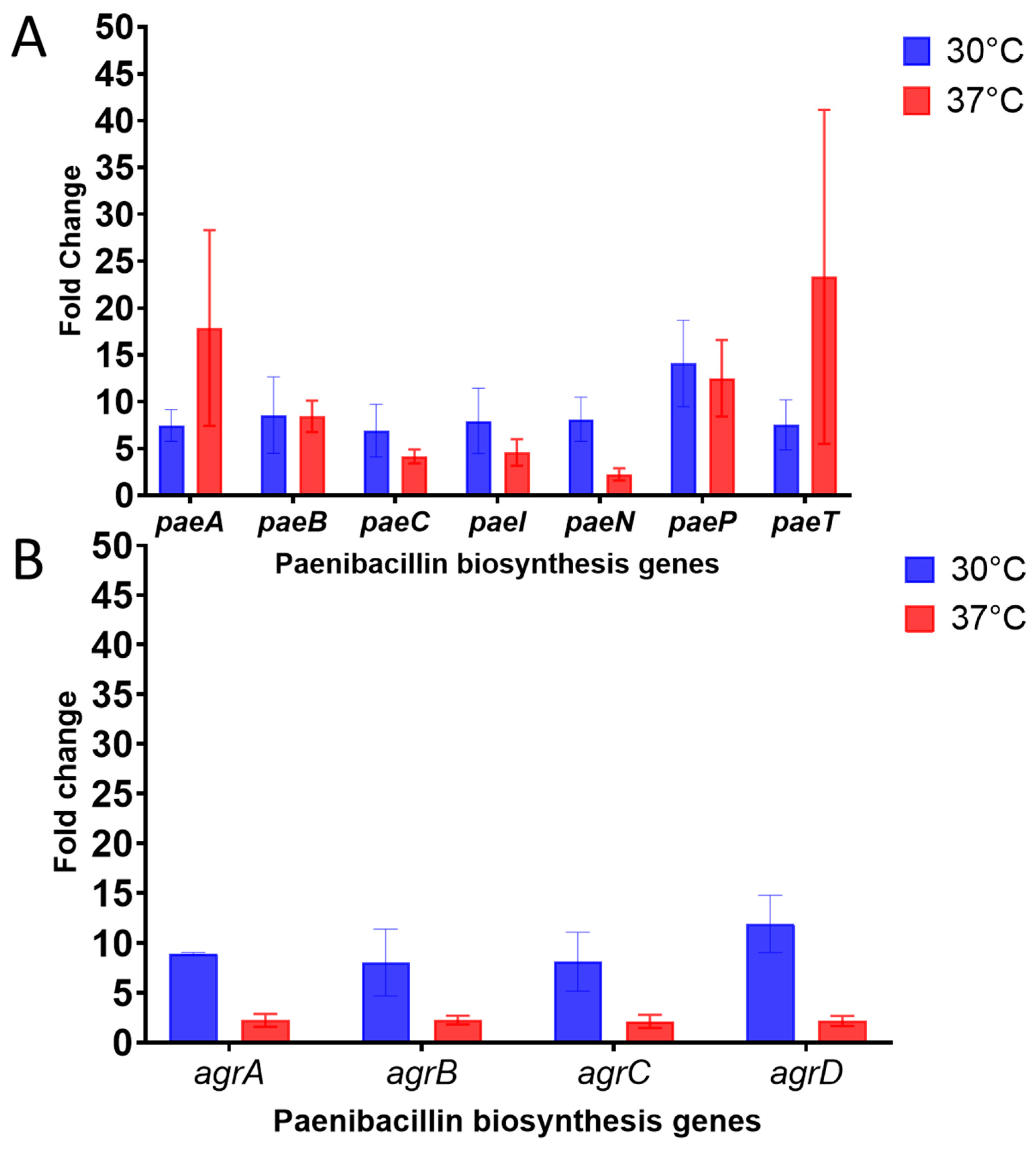
Figure 6). All the genes were upregulated in the mutant, with at least a 5-fold increase (at 30 °C) or a 2-fold increase (at 37 °C) compared with the wild type. It is important to note that the upregulated 11 genes are primarily responsible for driving paenibacillin production, modification by tailoring enzymes (peptidase, dehydratase, and cyclase), transportation, immunity, and the regulatory Agr system; the latter is presumed to play a crucial role in paenibacillin biosynthesis [
34].
The sensitivity of immunoassays can be impacted by the sample matrix [
35,
36]. The competitive ELISA for paenibacillin exhibited high sensitivity and accuracy, as demonstrated by its capacity to detect spiked paenibacillin in various matrices (
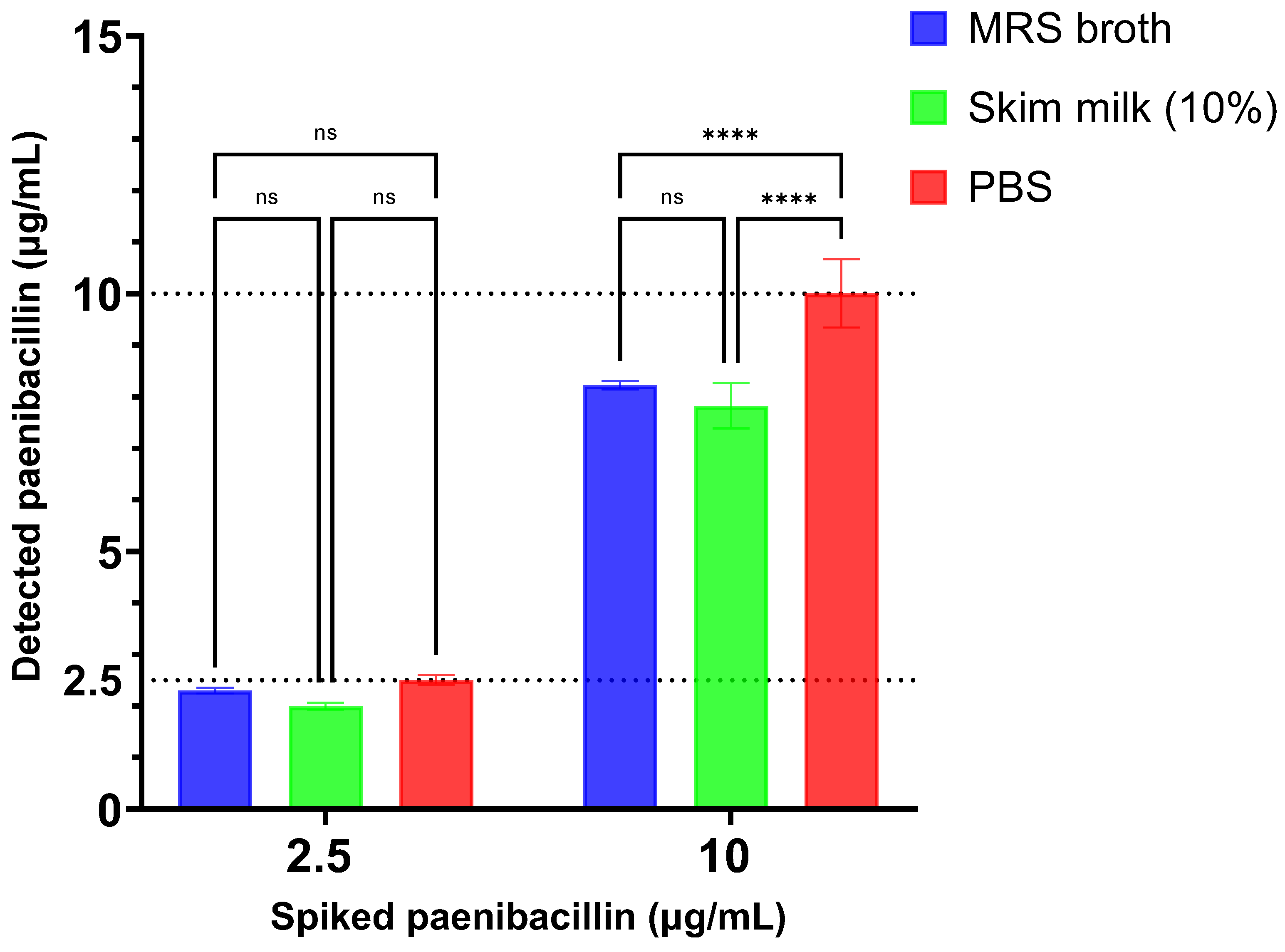
Figure 7) with high recover rates (≥78.2%). Paenibacillin was spiked at 2.5–10 µg/mL to potentially represent the approximate levels at which the peptide is produced by
P. polymyxa strains. However, paenibacillin recovery was slightly affected in MRS broth and skim milk solution when the antimicrobial peptide was spiked at 10 µg/mL. This could be attributed to the presence of proteins in these matrices that might compete with paenibacillin for binding to the anti-paenibacillin antibodies or to paenibacillin. Nonetheless, additional investigation is required to verify this assumption and to improve the recovery of the lantibiotic. Moreover, the competitive ELISA demonstrated sufficient repeatability in paenibacillin recovery from these matrices as evident by the low %CV. Specifically, for a spiked paenibacillin concentration of 2.5 µg/mL, the %CV values were 2.6% in MRS broth, 3.7% in 10% skim milk solution, and 3.8% in PBS. At a higher paenibacillin concentration of 10 µg/mL, the %CV values were 0.95% in MRS broth, 5.6% in 10% skim milk solution, and 6.6% in PBS. These findings indicate the assay’s limited variability across different matrices.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






