
A Lateral Flow-Recombinase Polymerase Amplification Method for Colletotrichum gloeosporioides Detection
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Comparative Genomic Analysis
2.3. Fungal Cultivation and Genomic DNA Extraction
2.4. Primer Design and Specificity Identification
2.5. Sensitivity Testing of Primers
2.6. Preparing DNA Extraction of Yam Anthracnose Samples
2.7. Development of a Rapid RPA Detection Kit against C. gloeosporioides
3. Results
3.1. Comparative Genomic Analsysis of 60 Genomes Revealed C. gloeosporioides-Specific Orthogroups
3.2. Screening of Specific Primers for C. gloeosporioides Detection
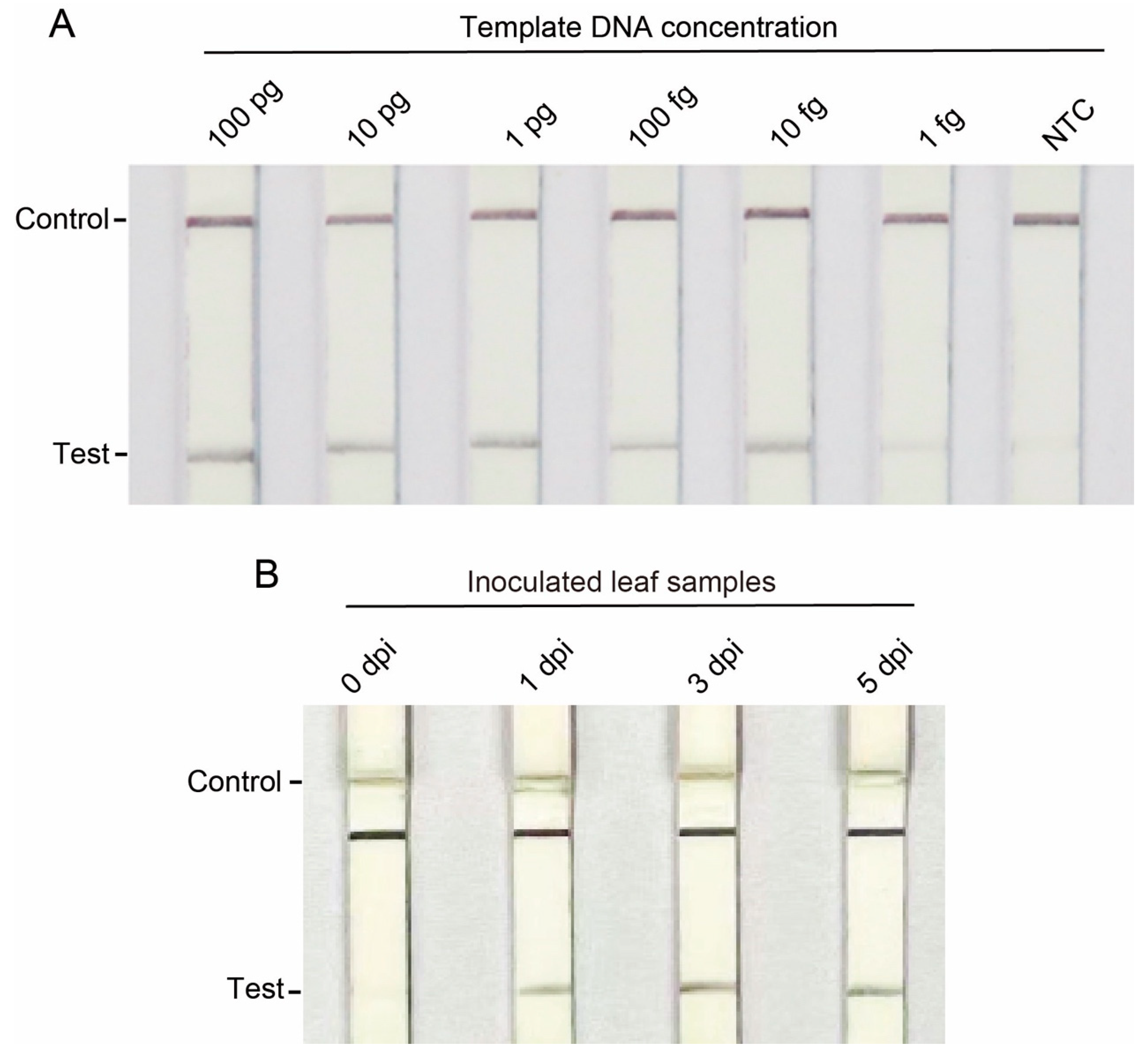
3.3. Sensitivity Testing of Primers for C. gloeosporioides Detection
3.4. Evaluation of Specific Primers for C. gloeosporioides Detection in Infected Plant Tissues
3.5. Development of the LF-RPA Method for Rapid Detection of C. gloeosporioides
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, P.; Shan, N.; Ali, A.; Sun, J.; Luo, S.; Xiao, Y.; Wang, S.; Hu, R.; Huang, Y.; Zhou, Q. Comprehensive Evaluation of Functional Components, Biological Activities, and Minerals of Yam Species (Dioscorea polystachya and D. alata) from China. LWT 2022, 168, 113964. [Google Scholar] [CrossRef]
- Lebot, V.; Lawac, F.; Legendre, L. The Greater Yam (Dioscorea alata L.): A Review of Its Phytochemical Content and Potential for Processed Products and Biofortification. J. Food Compos. Anal. 2023, 115, 104987. [Google Scholar] [CrossRef]
- More, S.J.; Ravi, V.; Raju, S.; de Freitas, S.T.; Pareek, S. Tropical Tuber Crops; CRC Press: Boca Raton, FL, USA, 2019; Volume 1, pp. 719–758. [Google Scholar]
- Ntui, V.O.; Uyoh, E.A.; Ita, E.E.; Markson, A.A.A.; Tripathi, J.N.; Okon, N.I.; Akpan, M.O.; Phillip, J.O.; Brisibe, E.A.; Ene-Obong, E.O.E. Strategies to Combat the Problem of Yam Anthracnose Disease: Status and Prospects. Mol. Plant Pathol. 2021, 22, 1302–1314. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.A.; Darkwa, K.; Olasanmi, B.; Kolade, O.; Mournet, P.; Bhattacharjee, R.; Lopez-Montes, A.; De Koeyer, D.; Adebola, P.; Kumar, L.; et al. Identification of QTLs Controlling Resistance to Anthracnose Disease in Water Yam (Dioscorea alata). Genes 2022, 13, 347. [Google Scholar] [CrossRef] [PubMed]
- Pornprapa, K.; Suriyasit, S.; Mongkutkarn, U.; Panisa, P.; Tanimnun, J. Cross-Resistance to Benzimidazole Group and Mancozeb Fungicides in Colletotrichum spp. Causing Anthracnose Disease. Sci. Technol. Asia 2022, 27, 400–408. [Google Scholar]
- Liu, F.; Ma, Z.; Hou, L.; Diao, Y.; Wu, W.; Damm, U.; Song, S.; Cai, L. Updating Species Diversity of Colletotrichum, with a Phylogenomic Overview. Stud. Mycol. 2022, 101, 1–56. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Talhinhas, P. Hosts of Colletotrichum. Mycosphere 2023, 14, 158–261. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life Styles of Colletotrichum Species and Implications for Plant Biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Luo, A.; Ling, C.; Ho, S.Y.W.; Zhu, C.D. Comparison of Methods for Molecular Species Delimitation Across a Range of Speciation Scenarios. Syst. Biol. 2018, 67, 830–846. [Google Scholar] [CrossRef]
- Attaluri, S.; Dharavath, R. Novel plant disease detection techniques-a brief review. Mol. Biol. Rep. 2023, 50, 9677–9690. [Google Scholar] [CrossRef]
- Kim, E.; Yang, S.M.; Kim, I.S.; Kim, H.Y. Identification of novel molecular targets for Weissella species-specific real-time PCR based on pangenome analysis. Appl. Microbiol. Biotechnol. 2022, 106, 4157–4168. [Google Scholar] [CrossRef] [PubMed]
- Chelkha, N.; Jardot, P.; Moussaoui, I.; Levasseur, A.; La Scola, B.; Colson, P. Core gene-based molecular detection and identification of Acanthamoeba species. Sci. Rep. 2020, 10, 1583. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.R.; Sreenivasaprasad, S.; Brown, A.E. Detection and differentiation of Colletotrichum gloeosporioides isolates using PCR. FEMS Microbiol. Lett. 1992, 77, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Vieira, W.A.d.S.; Bezerra, P.A.; Silva, A.C.d.; Veloso, J.S.; Câmara, M.P.S.; Doyle, V.P. Optimal Markers for the Identification of Colletotrichum Species. Mol. Phylogenetics Evol. 2020, 143, 106694. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.K.; Pritchard, L.; Birch, P.R. Comparative Genomics Reveals what Makes an Enterobacterial Plant Pathogen. Annu. Rev. Phytopathol. 2006, 44, 305–336. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust Demarcation of 17 Distinct Bacillus Species Clades, Proposed as Novel Bacillaceae Genera, by Phylogenomics and Comparative Genomic Analyses: Description of Robertmurraya kyonggiensis sp. nov. and Proposal for an Emended Genus Bacillus Limiting It Only to the Members of the Subtilis and Cereus Clades of Species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar] [PubMed]
- Miyauchi, S.; Kiss, E.; Kuo, A.; Drula, E.; Kohler, A.; Sánchez-García, M.; Morin, E.; Andreopoulos, B.; Barry, K.W.; Bonito, G. Large-Scale Genome Sequencing of Mycorrhizal Fungi Provides Insights into the Early Evolution of Symbiotic Traits. Nat. Commun. 2020, 11, 5125. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Duan, K.; Liu, Y.; Song, L.; Gao, Q.H. Method to Detect and Quantify Colonization of Anthracnose Causal Agent Colletotrichum gloeosporioides Species Complex in Strawberry by Real-time PCR. J. Phytopathol. 2022, 170, 326–336. [Google Scholar] [CrossRef]
- Syafitri, L.M.; Wibowo, A.; Widiastuti, A.; Subandiyah, S.; Harper, S. Molecular Identification of Colletotrichum gloeosporioides Causing Anthracnose on Shallot in Bantul, Yogyakarta, Indonesia. J. Biol. Divers. 2023, 24. [Google Scholar] [CrossRef]
- Babu, B.; Ochoa-Corona, F.M.; Paret, M.L. Recombinase polymerase amplification applied to plant virus detection and potential implications. Anal. Biochem. 2018, 546, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Macdonald, J.; Von Stetten, F. A Comprehensive Summary of a Decade Development of the Recombinase Polymerase Amplification. Analyst 2019, 144, 31–67. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yan, C.; Yu, H.; Zhang, Y.; Zhang, C.Q. Establishment of the Recombinase Polymerase Amplification-Lateral Flow Dipstick Detection Technique for Fusarium oxysporum. Plant Dis. 2023, 107, 2665–2672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, W.T.; Lu, R.S.; Chen, M.; Liu, J.; Sun, X.Q.; Zhang, Y.M. Genome Sequence Resource for Colletotrichum gloeosporioides, an Important Pathogenic Fungus Causing Anthracnose of Dioscorea alata. Plant Dis. 2023, 107, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.; Sultana, T.; Kesanakurti, P.; Hambleton, S. Genome Sequencing and Comparison of Five Tilletia Species to Identify Candidate Genes for the Detection of Regulated Species Infecting Wheat. IMA Fungus 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Alzohairy, A.M. BioEdit: An Important Software for Molecular Biology. Gerf Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Aamir, S. A Rapid and Efficient Method of fungal Genomic DNA Extraction, Suitable for PCR Based Molecular Methods. Plant Pathol. Quar. 2015, 5, 74–81. [Google Scholar] [CrossRef]
- Lalitha, S.J.B.S. Primer Premier 5. Biotech Softw. Internet Rep. Comput. Softw. J. Sci. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Huang, L.; Li, Q.C.; Zhang, Y.; Li, D.W.; Ye, J.R. Colletotrichum gloeosporioides Sensu Stricto is a Pathogen of Leaf Anthracnose on Evergreen Spindle Tree (Euonymus japonicus). Plant Dis. 2016, 100, 672–678. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Hu, Z.J.; Huang, Y.Y.; Lin, X.Y.; Feng, H.; Zhou, S.X.; Xie, Y.; Liu, X.X.; Liu, C.; Zhao, R.M.; Zhao, W.S.; et al. Loss and Natural Variations of Blast Fungal Avirulence Genes Breakdown Rice Resistance Genes in the Sichuan Basin of China. Front. Plant Sci. 2022, 13, 788876. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Ye, Q.; Chen, M.; Li, F.; Xiang, X.; Shang, Y.; Wang, C.; Zhang, J.; Xue, L.; Wang, J.; et al. Novel species-specific targets for real-time PCR detection of four common pathogenic Staphylococcus spp. Food Control 2022, 131, 108478. [Google Scholar] [CrossRef]
- Shang, Y.; Ye, Q.; Wu, Q.; Pang, R.; Zhou, B.; Wang, C.; Xiang, X.; Li, F.; Wang, J.; Zhang, Y.; et al. PCR and multiplex PCR assays for the detection of Cronobacter species using specific targets obtained by a bioinformatics approach. Food Control 2021, 125, 107896. [Google Scholar] [CrossRef]
- da Silva, L.L.; Moreno, H.L.A.; Correia, H.L.N.; Santana, M.F.; de Queiroz, M.V. Colletotrichum: Species complexes, lifestyle, and peculiarities of some sources of genetic variability. Appl. Microbiol. Biotechnol. 2020, 104, 1891–1904. [Google Scholar] [CrossRef] [PubMed]
- Raj, M.; Jeeva, M.; Hegde, V.; Vidyadharan, P.; Archana, P.; Senthil alias Sankar, M.; Nath, S.V. Polymerase Chain Reaction Assay for Rapid, Sensitive Detection, and Identification of Colletotrichum gloeosporioides Causing Greater Yam Anthracnose. Mol. Biotechnol. 2012, 52, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Yang, L.; Zhang, S.; Gao, J.; Jiang, L.; Luo, L. Real-time PCR for Detection and Quantification of C. gloeosporioides s.l. Growth in Stylosanthes and Arabidopsis. Crop Prot. 2022, 159, 106021. [Google Scholar] [CrossRef]
- Zou, Y.; Mason, M.G.; Wang, Y.; Wee, E.; Turni, C.; Blackall, P.J.; Trau, M.; Botella, J.R. Nucleic acid purification from plants, animals and microbes in under 30 seconds. PLoS Biol. 2017, 15, e2003916. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.-T.; Lu, X.-Y.; Wang, Y.; Li, M.-H.; Hu, K.; Shen, Z.-J.; Sun, X.-Q.; Zhang, Y.-M. A Lateral Flow-Recombinase Polymerase Amplification Method for Colletotrichum gloeosporioides Detection. J. Fungi 2024, 10, 315. https://0-doi-org.brum.beds.ac.uk/10.3390/jof10050315
Xu W-T, Lu X-Y, Wang Y, Li M-H, Hu K, Shen Z-J, Sun X-Q, Zhang Y-M. A Lateral Flow-Recombinase Polymerase Amplification Method for Colletotrichum gloeosporioides Detection. Journal of Fungi. 2024; 10(5):315. https://0-doi-org.brum.beds.ac.uk/10.3390/jof10050315
Chicago/Turabian StyleXu, Wei-Teng, Xin-Yu Lu, Yue Wang, Ming-Han Li, Ke Hu, Zi-Jie Shen, Xiao-Qin Sun, and Yan-Mei Zhang. 2024. "A Lateral Flow-Recombinase Polymerase Amplification Method for Colletotrichum gloeosporioides Detection" Journal of Fungi 10, no. 5: 315. https://0-doi-org.brum.beds.ac.uk/10.3390/jof10050315