
Tannin-Tolerant Saccharomyces cerevisiae Isolated from Traditional Fermented Tea Leaf (Miang) and Application in Fruit Wine Fermentation Using Longan Juice Mixed with Seed Extract as Substrate
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Chemicals
2.2. Sampling Sites and Sample Collection
2.3. Isolation and Screening of Tannin-Tolerant S. cerevisiae from Miang
2.3.1. Isolation of Yeast
2.3.2. Screening of Gas Formation and Ethanol Production
2.3.3. Tannin Tolerance Assessment of Ethanol-Producing Yeast
2.4. Molecular Identification of Tannin-Tolerant S. cerevisiae Isolated from Miang
2.5. Assessment of Ethanol Production Capability in Tannin-Enriched Medium
2.5.1. Viable Cell Count and pH Monitoring
2.5.2. Evaluation of Sugar Utilization and Ethanol Production
2.6. Application of Tannin-Tolerant S. cerevisiae in Longan Fruit Wine Fermentation Supplemented Seed Extract
2.6.1. Longan Seed Extraction
2.6.2. Preparation of Longan Fruit Wine Fermentation Supplemented Seed Extract
2.6.3. Analysis of Total Polyphenols, Total Tannin, and Total Flavonoids
2.6.4. Antioxidant and α-Glucosidase Inhibitory Activity
2.7. Statistical Analysis
3. Results and Discussion
3.1. Isolation of Tannin-Tolerant and Ethanol-Producing Yeast
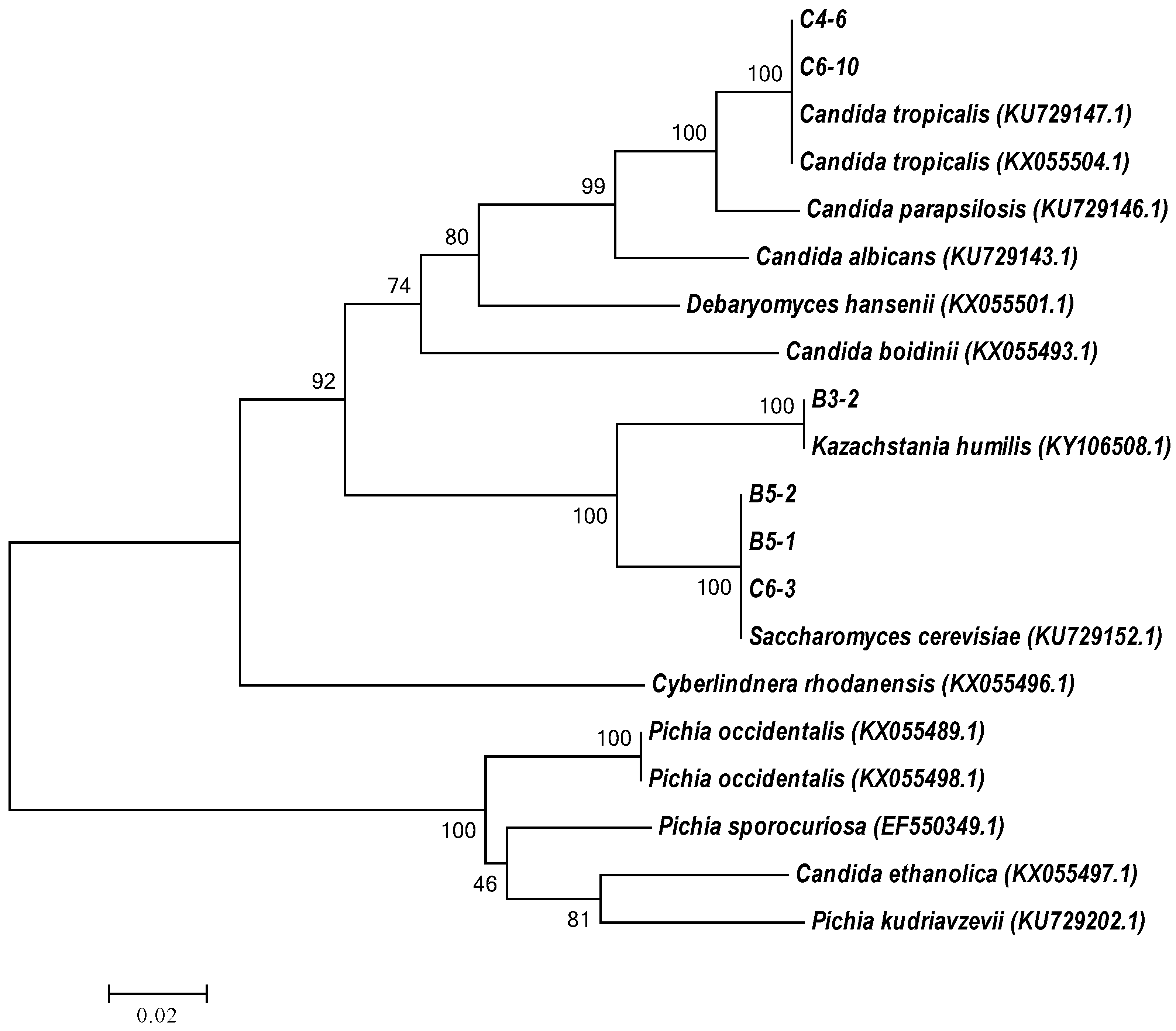
3.2. Molecular Identification of Tannin-Tolerant Selected Yeasts by rRNA Gene Sequence Analysis
3.3. Evaluation of Tannin-Tolerant S. cerevisiae Isolated from Miang on Ethanol Production with Tannin-Enriched Medium
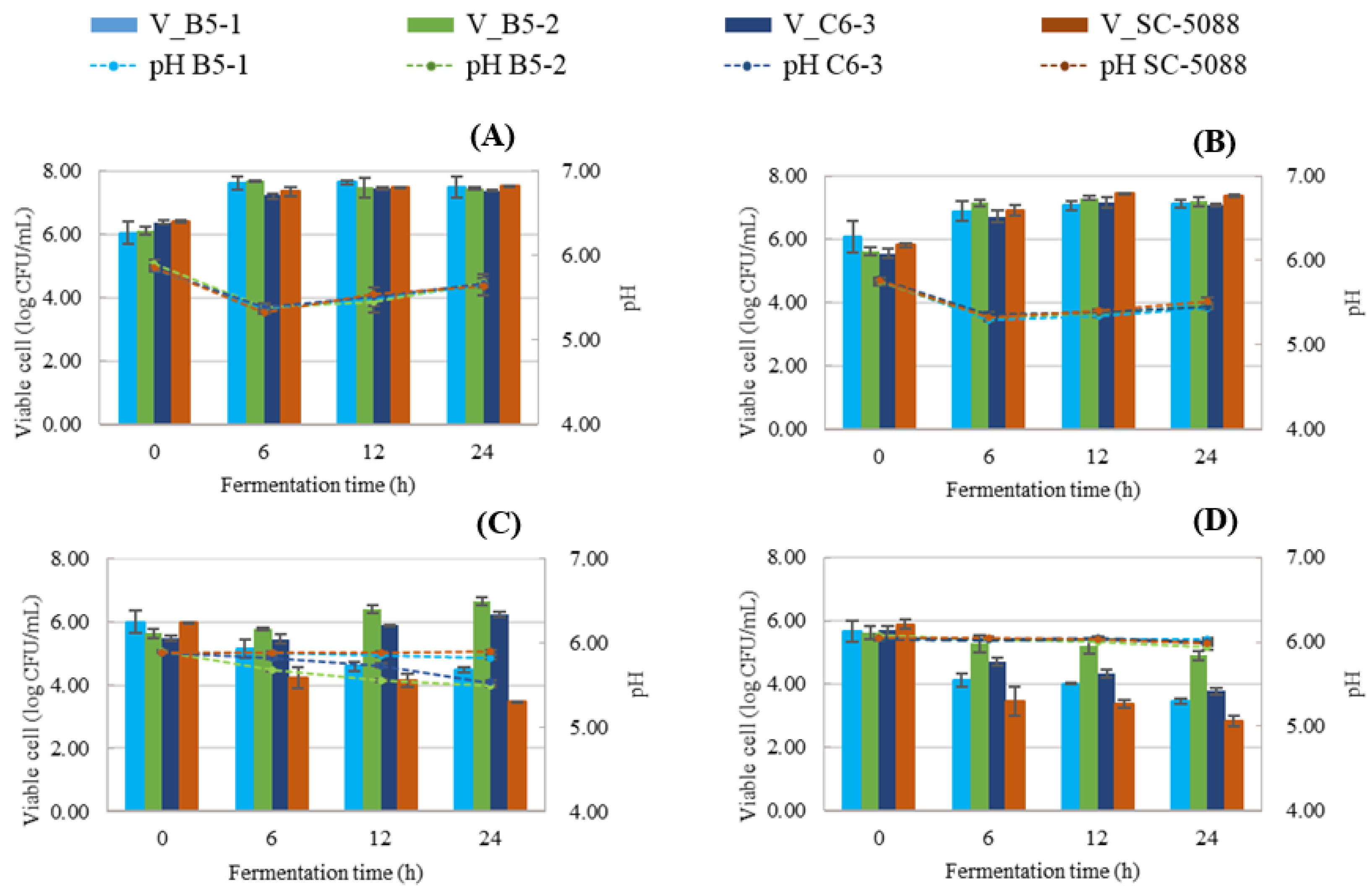
3.3.1. Viable Cell Count and pH of Tannin-Tolerant S. cerevisiae
3.3.2. Glucose Consumption and Ethanol Production
3.4. Dynamic Changes in Viable Cell Count and Biochemical Property during Longan Fruit Wine Fermentation Supplemented Longan Seed Extract
3.4.1. Viable Cell Count and pH of Tannin-Tolerant S. cerevisiae
3.4.2. Sugar Utilization and Ethanol Production
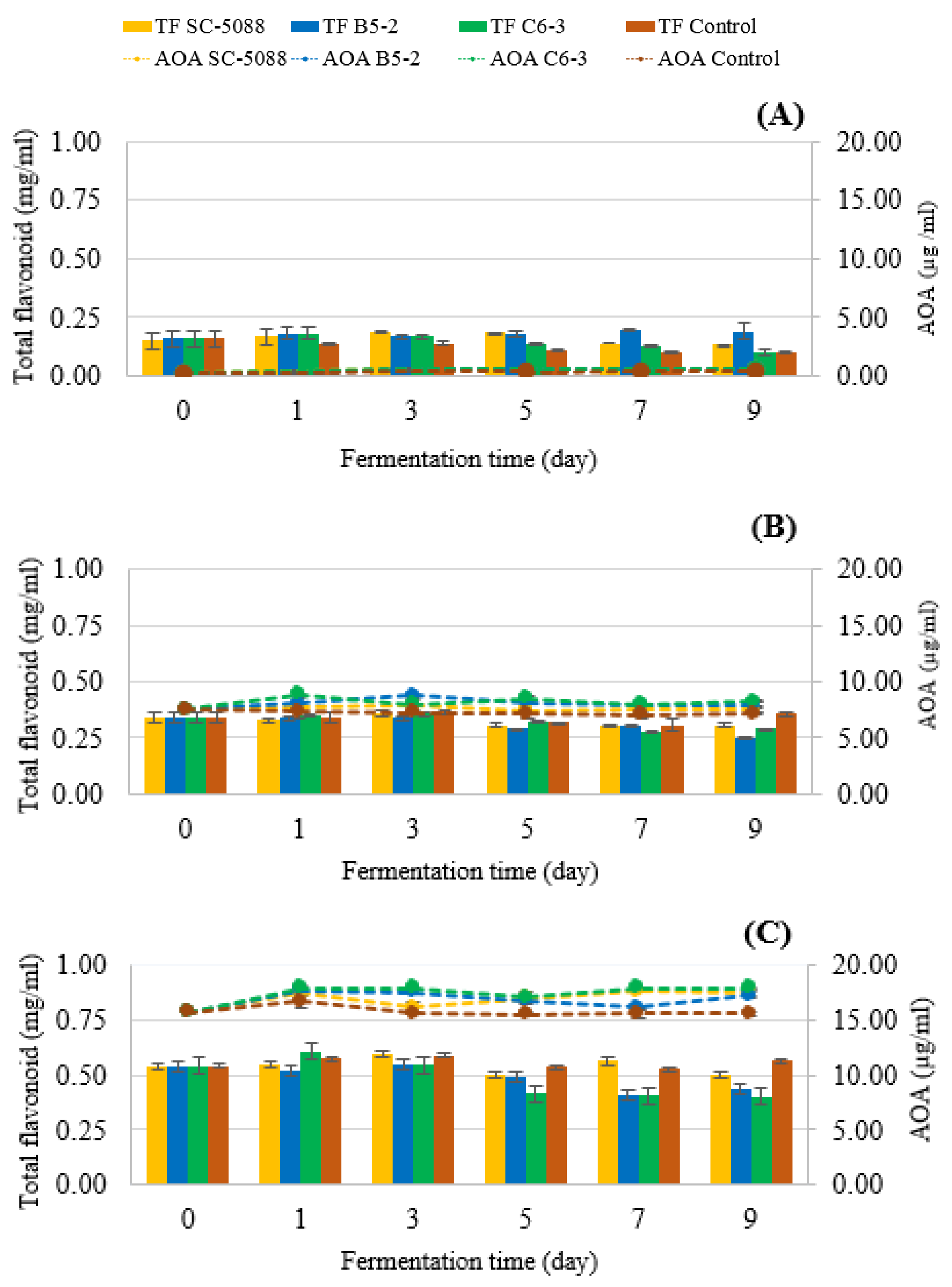
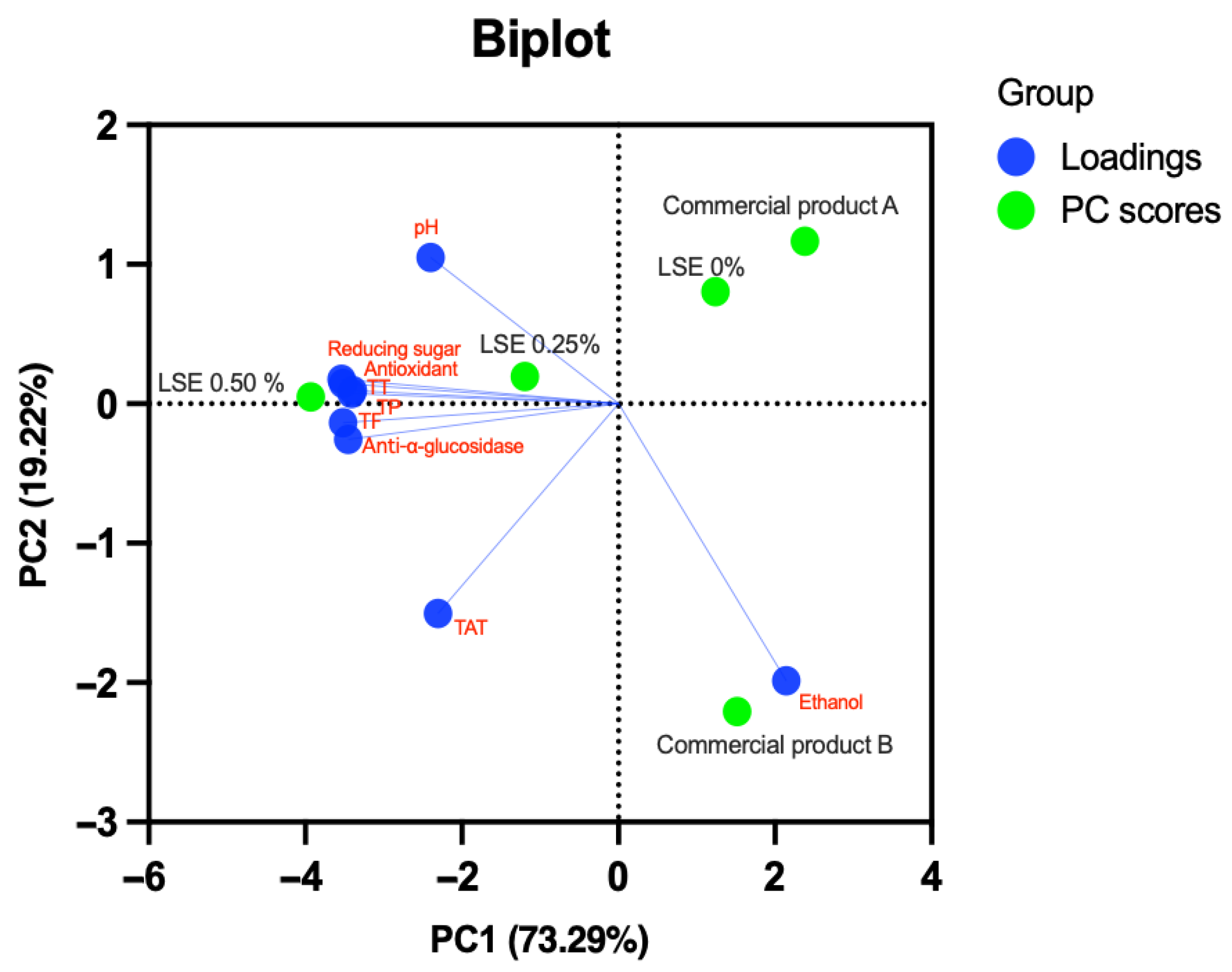
3.4.3. Bioactive Compound, Antioxidant Analysis, and α-Glucosidase Inhibitory Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces cerevisiae metabolites produced during fermentation on bread quality parameters: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1152–1164. [Google Scholar] [CrossRef] [PubMed]
- Le Jeune, C.; Erny, C.; Demuyter, C.; Lollier, M. Evolution of the population of Saccharomyces cerevisiae from grape to wine in a spontaneous fermentation. Food Microbiol. 2006, 23, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Gerőcs, A.; Nagy, T.; Nemes-Barnás, K.; Májer, J.; Szőke, B.Á.; Kővágó, R.; Magalhães, F.; Gibson, B.; Szekeres, A.; Juhász, Á.; et al. Characterization of Saccharomyces strains isolated from “Kéknyelű” grape must and their potential for wine production. Fermentation 2022, 8, 416. [Google Scholar] [CrossRef]
- Bouphun, T.; Wei, X.; Dan, W.; Zhao, R.; Qi, Z. Dynamic changes in chemical constituents during processing of Miang (Thai fermented tea leaf) in various degree of tea leaf maturity. Int. J. Food Eng. 2018, 4, 178–185. [Google Scholar] [CrossRef]
- Chaikaew, S.; Baipong, S.; Sone, T.; Kanpiengjai, A.; Chui-Chai, N.; Asano, K.; Khanongnuch, C. Diversity of lactic acid bacteria from Miang, a traditional fermented tea leaf in northern Thailand and their tannin-tolerant ability in tea extract. J. Microbiol. 2017, 55, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Unban, K.; Chaichana, W.; Baipong, S.; Abdullahi, A.D.; Kanpiengjai, A.; Shetty, K.; Khanongnuch, C. Probiotic and antioxidant properties of lactic acid bacteria isolated from indigenous fermented tea leaves (Miang) of north Thailand and promising application in synbiotic formulation. Fermentation 2021, 7, 195. [Google Scholar] [CrossRef]
- Kanpiengjai, A.; Chui-Chai, N.; Chaikaew, S.; Khanongnuch, C. Distribution of tannin-‘tolerant yeasts isolated from Miang, a traditional fermented tea leaf (Camellia sinensis var. assamica) in northern Thailand. Int. J. Food Microbiol. 2016, 238, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Unban, K.; Muangkajang, N.; Kodchasee, P.; Kanpiengjai, A.; Shetty, K.; Khanongnuch, C. Tannin-tolerant and extracellular tannase producing Bacillus isolated from traditional fermented tea leaves and their probiotic functional properties. Foods 2020, 9, 490. [Google Scholar] [CrossRef]
- Unban, K.; Muangkajang, N.; Kodchasee, P.; Kanpiengjai, A.; Shetty, K.; Khanongnuch, C. Tannin-tolerant Saccharomyces cerevisiae isolated from traditional fermented tea (Miang) of northern Thailand and its feasible applications. Microbiol. Res. 2023, 14, 1969–1983. [Google Scholar] [CrossRef]
- Kodchasee, P.; Pharin, N.; Suwannarach, N.; Unban, K.; Saenjum, C.; Kanpiengjai, A.; Shetty, K.; Zarnkow, M.; Khanongnuch, C. Assessment of Tannin Tolerant Non-Saccharomyces Yeasts Isolated from Miang for Production of Health-Targeted Beverage Using Miang Processing Byproducts. J. Fungi 2023, 9, 165. [Google Scholar] [CrossRef] [PubMed]
- Merle, J.; Birot, M.; Deleuze, H.; Trinsoutrot, P.; Carré, H.; Huyette, Q.; Charrier-El Bouhtoury, F. Valorization of Kraft black liquor and tannins via porous material production. Arab. J. Chem. 2019, 12, 4731–4739. [Google Scholar] [CrossRef]
- Gonçalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentão, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in Mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Kaprasob, R.; Sarkar, D.; Kerdchoechuen, O.; Laohakunjit, N.; Khanongnuch, C.; Shetty, K. Beneficial lactic acid bacteria based bioprocessing of cashew apple juice for targeting antioxidant nutraceutical inhibitors as relevant antidotes to type 2 diabetes. Process Biochem. 2019, 82, 40–50. [Google Scholar] [CrossRef]
- Foglia, F.; Drapcho, C.; Nghiem, J. Effectiveness of tannin removal by alkaline pretreatment on sorghum ethanol production. Fermentation 2022, 8, 274. [Google Scholar] [CrossRef]
- Timotheo, C.A.; Lauer, C.M. Toxicity of vegetable tannin extract from Acacia mearnsii in Saccharomyces cerevisiae. Int. J. Environ. Sci. Technol. 2018, 15, 659–664. [Google Scholar] [CrossRef]
- Farha, A.K.; Yang, Q.Q.; Kim, G.; Li, H.B.; Zhu, F.; Liu, H.Y.; Gan, R.Y.; Corke, H.K. Tannins as an alternative to antibiotics. Food Biosci. 2020, 38, 100751. [Google Scholar] [CrossRef]
- Lim, S.H.; Darah, I.; Jain, K. Antimicrobial activities of tannins extracted from Rhizophora apiculata barks. J. Trop. For. Sci. 2006, 18, 59–65. [Google Scholar]
- Chen, M.; Lin, H.; Zhang, S.; Lin, Y.; Chen, Y.; Lin, Y. Effects of adenosine triphosphate (ATP) treatment on postharvest physiology, quality and storage behavior of longan fruit. Food Bioprocess Technol. 2015, 8, 971–982. [Google Scholar] [CrossRef]
- Sai-Ut, S.; Kingwascharapong, P.; Mazumder, M.A.R.; Rawdkuen, S. Optimization of ethanolic extraction of phenolic antioxidants from lychee and longan seeds using response surface methodology. Foods 2023, 12, 2827. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, T.; Wang, Q.; Shi, Y. Antioxidant, bacteriostatic and breservative Effects of extractable condensed tannins isolated from longan pericarps and seeds. Plants 2023, 12, 512. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liang, Y.; Zhang, Y.; Yi, X.; Wang, H.; Pan, Y. Antioxidant activity of longan (Dimocarpus longan L.) seed extract. Nuts Seeds Health Dis. Prev. 2011, 83, 705–710. [Google Scholar]
- Zheng, G.; Xu, L.; Wu, P.; Xie, H.; Jiang, Y.; Chen, F.; Wei, X. Polyphenols from longan seeds and their radical-scavenging activity. Food Chem. 2009, 116, 433–436. [Google Scholar] [CrossRef]
- Lagariya, L.; Soni, K.; Shah, J.S. Antitumor effects of polyphenol-rich extract of Euphoria longana seed by vascular endothelial growth factor and transforming growth factor-beta signaling inhibition in experimentally induced oral cancer in rats. Indian J. Pharmacol. 2022, 54, 329. [Google Scholar] [PubMed]
- Tandee, K.; Kittiwachana, S.; Mahatheeranont, S. Antioxidant activities and volatile compounds in longan (Dimocarpus longan Lour.) wine produced by incorporating longan seeds. Food Chem. 2021, 348, 128921. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Jiang, Y.; Shi, J.; Chen, F.; Ashraf, M. Extraction and pharmacological properties of bioactive compounds from longan (Dimocarpus Longan Lour.) fruit—A review. Food Res. Int. 2011, 44, 1837–1842. [Google Scholar] [CrossRef]
- Tseng, H.C.; Wu, W.T.; Huang, H.S.; Wu, M.C. Antimicrobial activities of various fractions of longan (Dimocarpus longan Lour. Fen Ke) seed extract. Int. J. Food Sci. Nutr. 2014, 65, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5’end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, Z.; Malik, S.; Mehta, A. In vitro and in vivo anti-cancer efficacy of Adiantum capillus-veneris L. against some selected human cancer cell lines and on ehrlich ascites carcinoma mouse model. Int. J. Pharm. Sci. Drug Res. 2020, 12, 267–274. [Google Scholar] [CrossRef]
- Makkar, H.P.; Becker, K. Behaviour of tannic acid from various commercial sources towards redox, metal complexing and protein precipitation assays of tannins. J. Sci. Food Agric. 1993, 62, 295–299. [Google Scholar] [CrossRef]
- Eom, S.H.; Park, H.J.; Jin, C.W.; Kim, D.O.; Seo, D.W.; Jeong, Y.H.; Cho, D.H. Changes in antioxidant activity with temperature and time in Chrysanthemum indicum L.(Gamguk) teas during elution processes in hot water. Food Sci. Biotechnol. 2008, 17, 408–412. [Google Scholar]
- Kodchasee, P.; Nain, K.; Abdullahi, A.D.; Unban, K.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Microbial dynamics-linked properties and functional metabolites during Miang fermentation using the filamentous fungi growth-based process. Food Biosci. 2021, 41, 100998. [Google Scholar] [CrossRef]
- Kham, N.N.N.; Phovisay, S.; Unban, K.; Kanpiengjai, A.; Saenjum, C.; Lumyong, S.; Shetty, K.; Khanongnuch, C. A Thermotolerant yeast Cyberlindnera rhodanensis DK isolated from Laphet-so capable of extracellular thermostable β-glucosidase production. J. Fungi 2024, 10, 243. [Google Scholar] [CrossRef]
- Zuza-Alves, D.L.; Silva-Rocha, W.P.; Chaves, G.M. An update on Candida tropicalis based on basic and clinical approaches. Front. Microbiol. 2017, 8, 1927. [Google Scholar] [CrossRef] [PubMed]
- Ou, Q.X.; Nikolic-Jaric, M.; Gänzle, M. Mechanisms of inactivation of Candida humilis and Saccharomyces cerevisiae by pulsed electric fields. Bioelectrochemistry 2017, 115, 47–55. [Google Scholar] [CrossRef]
- Al Bagieh, N.H.; Al Bageah, H.N. Effect of tannic acid on growth and acid production of Candida albicans. Pak. Oral Dent. J. 2014, 34, 405–408. [Google Scholar]
- Chung, K.T.; Wei, C.I.; Johnson, M.G. Are tannins a double-edged sword in biology and health? Trends Food Sci. Technol. 1998, 9, 168–175. [Google Scholar] [CrossRef]
- Salamah, A.; Srihardyastutie, A.; Prasetyawan, S.; Safitri, A. Influence of mixed cultures of Saccharomyces cerevisiae and Acetobacter aceti for hydrolysis of tannins in the cabbage fermentation (Brassica oleracea L. var. capitata). IOP Conf. Ser. Mater. Sci. Eng. 2019, 546, 062028. [Google Scholar] [CrossRef]
- Margaritis, A.; Merchant, F.J.; Abbott, B.J. Advances in ethanol production using immobilized cell systems. Crit. Rev. Biotechnol. 1983, 1, 339–393. [Google Scholar] [CrossRef]
- Mohapatra, S.; Mishra, S.S.; Das, S.K.; Thatoi, H. Influence of reactors, microbial carbohydrate uptake, and metabolic pathways on ethanol production from grass biomass: A review. Int. J. Energy Res. 2019, 43, 1615–1646. [Google Scholar] [CrossRef]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N.; Lavigne, V. Synthesis of volatile phenols by Saccharomyces cerevisiae in wines. J. Sci. Food Agric. 1993, 62, 191–202. [Google Scholar] [CrossRef]
- Koschwanez, J.H.; Foster, K.R.; Murray, A.W. Sucrose utilization in budding yeast as a model for the origin of undifferentiated multicellularity. PLoS Biol. 2011, 9, e1001122. [Google Scholar] [CrossRef]
- Wauters, T.; Iserentant, D.; Verachtert, H. Sensitivity of Saccharomyces cerevisiae to tannic acid is due to iron deprivation. Can. J. Microbiol. 2001, 47, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Sauer, U. The importance of post-translational modifications in regulating Saccharomyces cerevisiae metabolism. FEMS Yeast Res. 2012, 12, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Alberti, A.; dos Santos, T.P.M.; Zielinski, A.A.F.; dos Santos, C.M.E.; Braga, C.M.; Demiate, I.M.; Nogueira, A. Impact on chemical profile in apple juice and cider made from unripe, ripe and senescent dessert varieties. LWT-Food Sci. Technol. 2016, 65, 436–443. [Google Scholar] [CrossRef]
- Alcalde-Eon, C.; Ferreras-Charro, R.; Ferrer-Gallego, R.; Rivero, F.J.; Heredia, F.J.; Escribano-Bailón, M.T. Monitoring the effects and side-effects on wine colour and flavonoid composition of the combined post-fermentative additions of seeds and mannoproteins. Food Res. Int. 2019, 126, 108650. [Google Scholar] [CrossRef] [PubMed]
- Bimpilas, A.; Tsimogiannis, D.; Balta-Brouma, K.; Lymperopoulou, T.; Oreopoulou, V. Evolution of phenolic compounds and metal content of wine during alcoholic fermentation and storage. Food Chem. 2015, 178, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Minnaar, P.P.; Du Plessis, H.W.; Paulsen, V.; Ntushelo, N.; Jolly, N.P.; Du Toit, M. Saccharomyces cerevisiae, non-Saccharomyces yeasts and lactic acid bacteria in sequential fermentations: Effect on phenolics and sensory attributes of South African Syrah wines. S. Afr. J. Enol. Vitic. 2017, 38, 237–244. [Google Scholar] [CrossRef]
- De Nicola, R. Interaction of Zinc with the Yeast Saccharomyces cerevisiae. Doctoral Dissertation, University of Abertay Dundee, Dundee, UK, 2006. [Google Scholar]
- Walker, G.M.; Stewart, G.G. Saccharomyces cerevisiae in the production of fermented beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Y.; He, X.M.; Sun, J.; Li, C.B.; Li, L.; Sheng, J.F.; Xin, M.; Li, Z.C.; Zheng, F.J.; Liu, G.M.; et al. Polyphenols and alkaloids in byproducts of longan fruits (Dimocarpus Longan Lour.) and their bioactivities. Molecules 2019, 24, 1186. [Google Scholar] [CrossRef]
- Rakariyatham, K.; Zhou, D.; Rakariyatham, N.; Shahidi, F. Sapindaceae (Dimocarpus Longan and Nephelium Lappaceum) seed and peel by-products: Potential sources for phenolic compounds and use as functional ingredients in food and health applications. J. Funct. Foods 2020, 67, 103846. [Google Scholar] [CrossRef]
- Lin, C.C.; Chung, Y.C.; Hsu, C.P. Potential roles of longan flower and seed extracts for anti-cancer. World J. Exp. Med. 2012, 2, 78. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Mishra, S.S.; Das, S.K.; Thatoi, H. Modern pharmacological actions of longan fruits and their usages in traditional herbal remedies. J. Med. Plants Stud. 2019, 7, 179–185. [Google Scholar]
- Buntum, T.; Kakumyan, P.; Surassmo, S.; Thanomsilp, C.; Suwantong, O. Potential of longan seed extract-loaded alginate-chitosan beads as drug delivery system. Front. Mater. 2022, 9, 818595. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Regions | No. of Samples | No. of Yeast Isolate | No. of Gas Formation | No. of Ethanol Producing Yeast | Location |
|---|---|---|---|---|---|---|
| 1 | Xayaboury district | 9 | 7 | 2 | 0 | 19.268576″ N, 101.707712″ E |
| 2 | Xienghorn district | 15 | 13 | 4 | 3 | 19.587956″ N, 100.807902″ E |
| 3 | Khop district | 9 | 15 | 4 | 3 | 19.41′03.2″ N, 100.30′07.1″ E |
| 4 | Saysathan district | 10 | 0 | 0 | 0 | 19.405888″ N, 101.387887″ E |
| Total | 43 | 35 | 10 | 6 |
| Isolate (Code) | Species Name | Similarity (%) | Query Length (bps) | Accession Number |
|---|---|---|---|---|
| B3-2 | Kazachstania humilis | 100 | 605 | KY106507.1 |
| B5-1 | Saccharomyces cerevisiae | 100 | 603 | NG_042623.1 |
| B5-2 | Saccharomyces cerevisiae | 100 | 583 | NG_042623.1 |
| C4-6 | Candida tropicalis | 100 | 594 | KU729147.1 |
| C6-3 | Saccharomyces cerevisiae | 99.64 | 596 | NG_042623.1 |
| C6-10 | Candida tropicalis | 100 | 600 | KU729147.1 |
| Parameter | TA Conc. (%) | S. cerevisiae B5-1 | S. cerevisiae B5-2 | S. cerevisiae C6-3 | S. cerevisiae 5088 |
|---|---|---|---|---|---|
| Ethanol Productivity (g/L·h) at 24 h | 0.00 | 0.37 ± 0.01 | 0.37 ± 0.01 | 0.37 ± 0.01 | 0.37 ± 0.01 |
| 1.00 | 0.29 + 0.01 b | 0.36 ± 0.01 a | 0.36 ± 0.01 a | 0.35 ± 0.01 a | |
| 2.50 | - | 0.23 ± 0.01 a | 0.21 ± 0.01 b | - | |
| 5.00 | - | - | - | - | |
| Ethanol fermentation efficiency (%) at 24 h | 0.00 | 87.31 ± 0.21 | 87.43 ± 0.98 | 86.80 ± 0.31 | 87.86 ± 1.87 |
| 1.00 | 74.81 ± 1.77 b | 83.79±1.54 a | 84.39 ± 0.29 a | 83.75 ± 0.81 a | |
| 2.50 | - | 54.03 ± 0.88 a | 49.92 ± 0.76 b | - | |
| 5.00 | - | - | - | - |
| Parameters | LSE 0% | LSE 0.25% | LSE 0.50% | Commercial Product A | Commercial Product B |
|---|---|---|---|---|---|
| pH | 4.49 ± 0.01 a | 4.46 ± 0.01 a | 4.48 ± 0.01 a | 3.52 ± 0.01 b | 3.37 ± 0.01 b |
| TAT (g/L) | 0.05 ± 0.01 b | 0.07 ± 0.01 ab | 0.08 ± 0.01 a | 0.03 ± 0.01 c | 0.08 ± 0.01 a |
| Reducing sugar (g/L) | 1.61 ± 0.03 c | 2.97 ± 0.16 b | 4.34 ± 0.24 a | 0.63 ± 0.01 d | 1.00 ± 0.10 d |
| Ethanol (% w/v) | 6.65 ± 0.72 b | 6.61 ± 0.17 b | 6.64 ± 0.9 b | 5.45 ± 0.13 bc | 10.81 ± 0.28 a |
| TP (g/L) | 0.33 ± 0.01 c | 1.59 ± 0.05 b | 5.49 ± 0.09 a | 0.15 ± 0.02 c | 0.29 ± 0.30 c |
| TT (g/L) | 0.12 ± 0.02 c | 1.27 ± 0.05 b | 4.97 ± 0.11 a | 0.03 ± 0.01 c | 0.07 ± 0.22 c |
| TF (g/L) | 0.10 ± 0.01 c | 0.29 ± 0.01 b | 0.40 ± 0.04 a | 0.07 ± 0.01 c | 0.13 ± 0.01 c |
| Antioxidant (µg/mL) | 0.59 ± 0.01 c | 8.25 ± 0.39 b | 17.88 ± 0.13 a | 0.14 ± 0.01 c | 0.26 ± 0.02 c |
| Anti α-glucosidase (%) | 28.78 ± 2.12 d | 77.71 ±0.03 b | 95.02 ±0.44 a | 19.90 ± 0.58 e | 39.59 ± 0.82 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phovisay, S.; Kodchasee, P.; Abdullahi, A.D.; Kham, N.N.N.; Unban, K.; Kanpiengjai, A.; Saenjum, C.; Shetty, K.; Khanongnuch, C. Tannin-Tolerant Saccharomyces cerevisiae Isolated from Traditional Fermented Tea Leaf (Miang) and Application in Fruit Wine Fermentation Using Longan Juice Mixed with Seed Extract as Substrate. Foods 2024, 13, 1335. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13091335
Phovisay S, Kodchasee P, Abdullahi AD, Kham NNN, Unban K, Kanpiengjai A, Saenjum C, Shetty K, Khanongnuch C. Tannin-Tolerant Saccharomyces cerevisiae Isolated from Traditional Fermented Tea Leaf (Miang) and Application in Fruit Wine Fermentation Using Longan Juice Mixed with Seed Extract as Substrate. Foods. 2024; 13(9):1335. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13091335
Chicago/Turabian StylePhovisay, Somsay, Pratthana Kodchasee, Aliyu Dantani Abdullahi, Nang Nwet Noon Kham, Kridsada Unban, Apinun Kanpiengjai, Chalermpong Saenjum, Kalidas Shetty, and Chartchai Khanongnuch. 2024. "Tannin-Tolerant Saccharomyces cerevisiae Isolated from Traditional Fermented Tea Leaf (Miang) and Application in Fruit Wine Fermentation Using Longan Juice Mixed with Seed Extract as Substrate" Foods 13, no. 9: 1335. https://0-doi-org.brum.beds.ac.uk/10.3390/foods13091335