
Prenylated Flavonoids of the Moraceae Family: A Comprehensive Review of Their Biological Activities
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Biosynthetic Pathway of Prenylated Flavonoids
3. Biological Activities of Prenylated Flavonoids
3.1. Anti-Diabetic Activity
3.2. Anti-Cancer Activity
3.3. Antimicrobial Activity
3.4. Antioxidant Activity
3.5. Anti-Inflammatory Activity
3.6. Enzyme Inhibition Activity
3.7. Anti-Alzheimer and Neuroprotective Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samec, D.; Karalija, E.; Sola, I.; Vujcic Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Tomás-Barberán, F.A.; García-Villalba, R. Structural Diversity of Polyphenols and Distribution in Foods. In Dietary Polyphenols; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 1–29. [Google Scholar]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Linic, I.; Samec, D.; Gruz, J.; Vujcic Bok, V.; Strnad, M.; Salopek-Sondi, B. Involvement of Phenolic Acids in Short-Term Adaptation to Salinity Stress is Species-Specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, W.; Li, W.Z.; Li, Y. A facile synthetic approach to prenylated flavanones: First total syntheses of (+/−)-bonannione A and (+/−)-sophoraflavanone A. J. Nat. Prod. 2001, 64, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.S.; Wei, J.G.; Peng, W.B.; Wang, S.M.; Sun, C.; Yang, Y.P.; Liu, K.C.; Li, X.L. Cytotoxic prenylated bibenzyls and flavonoids from Macaranga kurzii. Fitoterapia 2014, 99, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-X.; Liu, G.-Y.; Lou, J.; Wang, H.; Li, L.-M.; Hu, Q.-F.; Ye, Y.-Q. A New Cytotoxic Prenylated Chalcone from Desmodium renifolium. Asian J. Chem. 2015, 27, 2733–2734. [Google Scholar] [CrossRef]
- Chen, R.; Liu, X.; Zou, J.; Yin, Y.; Ou, B.; Li, J.; Wang, R.; Xie, D.; Zhang, P.; Dai, J. Regio- and Stereospecific Prenylation of Flavonoids by Sophora flavescens Prenyltransferase. Adv. Synth. Catal. 2013, 355, 1817–1828. [Google Scholar] [CrossRef]
- Li, H.; Ban, Z.; Qin, H.; Ma, L.; King, A.J.; Wang, G. A heteromeric membrane-bound prenyltransferase complex from hop catalyzes three sequential aromatic prenylations in the bitter acid pathway. Plant Physiol. 2015, 167, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Mito, K.; Ohara, K.; Yamamoto, H.; Yazaki, K. Cloning and characterization of naringenin 8-prenyltransferase, a flavonoid-specific prenyltransferase of Sophora flavescens. Plant Physiol. 2008, 146, 1075–1084. [Google Scholar] [CrossRef]
- Shen, G.; Huhman, D.; Lei, Z.; Snyder, J.; Sumner, L.W.; Dixon, R.A. Characterization of an isoflavonoid-specific prenyltransferase from Lupinus albus. Plant Physiol. 2012, 159, 70–80. [Google Scholar] [CrossRef]
- Mukai, R. Prenylation enhances the biological activity of dietary flavonoids by altering their bioavailability. Biosci. Biotechnol. Biochem. 2018, 82, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; Máñez, S. Recent trends in the pharmacological activity of isoprenyl phenolics. Curr. Med. Chem. 2013, 20, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.-W.; Wang, Q.-L.; Luo, M.; Zhu, M.-D.; Liang, H.-M.; Li, W.-J.; Cai, H.; Zhou, Z.-B.; Wang, H.; Tong, S.-Q.; et al. Phytochemistry and pharmacology of natural prenylated flavonoids. Arch. Pharmacal Res. 2023, 46, 207–272. [Google Scholar] [CrossRef] [PubMed]
- Frodin, D.G. History and concepts of big plant genera. Taxon 2004, 53, 753–776. [Google Scholar] [CrossRef]
- Levisson, M.; Araya-Cloutier, C.; de Bruijn, W.J.C.; van der Heide, M.; Salvador Lopez, J.M.; Daran, J.M.; Vincken, J.P.; Beekwilder, J. Toward Developing a Yeast Cell Factory for the Production of Prenylated Flavonoids. J. Agric. Food Chem. 2019, 67, 13478–13486. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef]
- Li, Z.; Sharkey, T.D. Metabolic profiling of the methylerythritol phosphate pathway reveals the source of post-illumination isoprene burst from leaves. Plant Cell Environ. 2013, 36, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Fang, X.; Yang, J.; Yao, Y.; Nandakumar, K.S.; Salem, M.L.; Cheng, K. Recent Research on Flavonoids and their Biomedical Applications. Curr. Med. Chem. 2021, 28, 1042–1066. [Google Scholar] [CrossRef]
- Shi, S.; Li, J.; Zhao, X.; Liu, Q.; Song, S.J. A comprehensive review: Biological activity, modification and synthetic methodologies of prenylated flavonoids. Phytochemistry 2021, 191, 112895. [Google Scholar] [CrossRef]
- Inui, S.; Hosoya, T.; Shimamura, Y.; Masuda, S.; Ogawa, T.; Kobayashi, H.; Shirafuji, K.; Moli, R.T.; Kozone, I.; Shin-ya, K.; et al. Solophenols B-D and solomonin: New prenylated polyphenols isolated from propolis collected from the Solomon Islands and their antibacterial activity. J. Agric. Food Chem. 2012, 60, 11765–11770. [Google Scholar] [CrossRef]
- Sun, D.; Hurdle, J.G.; Lee, R.; Lee, R.; Cushman, M.; Pezzuto, J.M. Evaluation of flavonoid and resveratrol chemical libraries reveals abyssinone II as a promising antibacterial lead. ChemMedChem 2012, 7, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Hatano, T.; Aga, Y.; Shintani, Y.; Ito, H.; Okuda, T.; Yoshida, T. Minor flavonoids from licorice. Phytochemistry 2000, 55, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2022, 21, 1049–1079. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.W.; Lee, B.W.; Curtis-Long, M.J.; Jung, S.; Ryu, Y.B.; Lee, W.S.; Park, K.H. Polyphenols from Broussonetia papyrifera displaying potent alpha-glucosidase inhibition. J. Agric. Food Chem. 2010, 58, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Quang, T.H.; Ngan, N.T.; Yoon, C.S.; Cho, K.H.; Kang, D.G.; Lee, H.S.; Kim, Y.C.; Oh, H. Protein Tyrosine Phosphatase 1B Inhibitors from the Roots of Cudrania tricuspidata. Molecules 2015, 20, 11173–11183. [Google Scholar] [CrossRef] [PubMed]
- Tabopda, T.K.; Ngoupayo, J.; Awoussong, P.K.; Mitaine-Offer, A.C.; Ali, M.S.; Ngadjui, B.T.; Lacaille-Dubois, M.A. Triprenylated flavonoids from Dorstenia psilurus and their alpha-glucosidase inhibition properties. J. Nat. Prod. 2008, 71, 2068–2072. [Google Scholar] [CrossRef]
- Shi, Z.F.; Lei, C.; Yu, B.W.; Wang, H.Y.; Hou, A.J. New Alkaloids and alpha-Glucosidase Inhibitory Flavonoids from Ficus hispida. Chem. Biodivers. 2016, 13, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Li, W.; Huang, X.; Zhang, R.; Tian, K.; Hou, S.; Li, Y. Antioxidant and alpha-glucosidase inhibitory activities of isoflavonoids from the rhizomes of Ficus tikoua Bur. Nat. Prod. Res. 2018, 32, 399–405. [Google Scholar] [CrossRef]
- Jo, Y.H.; Lee, S.; Yeon, S.W.; Ryu, S.H.; Turk, A.; Hwang, B.Y.; Han, Y.K.; Lee, K.Y.; Lee, M.K. Anti-alpha-glucosidase and anti-oxidative isoflavonoids from the immature fruits of Maclura tricuspidata. Phytochemistry 2022, 194, 113016. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Y.; Wang, Y.; Zhang, Y. Bioassay-guided screening and isolation of α-glucosidase and tyrosinase inhibitors from leaves of Morus alba. Food Chem. 2012, 131, 617–625. [Google Scholar] [CrossRef]
- Li, M.; Wu, X.; Wang, X.; Shen, T.; Ren, D. Two novel compounds from the root bark of Morus alba L. Nat. Prod. Res. 2018, 32, 36–42. [Google Scholar] [CrossRef]
- Ha, M.T.; Seong, S.H.; Nguyen, T.D.; Cho, W.K.; Ah, K.J.; Ma, J.Y.; Woo, M.H.; Choi, J.S.; Min, B.S. Chalcone derivatives from the root bark of Morus alba L. act as inhibitors of PTP1B and alpha-glucosidase. Phytochemistry 2018, 155, 114–125. [Google Scholar] [CrossRef]
- Zhao, Y.; Kongstad, K.T.; Jager, A.K.; Nielsen, J.; Staerk, D. Quadruple high-resolution alpha-glucosidase/alpha-amylase/PTP1B/radical scavenging profiling combined with high-performance liquid chromatography-high-resolution mass spectrometry-solid-phase extraction-nuclear magnetic resonance spectroscopy for identification of antidiabetic constituents in crude root bark of Morus alba L. J. Chromatogr. A 2018, 1556, 55–63. [Google Scholar] [CrossRef]
- Qu, K.J.; Wang, B.; Jiang, C.S.; Xie, B.G.; Liu, A.H.; Li, S.W.; Guo, Y.W.; Li, J.; Mao, S.C. Rearranged Diels-Alder Adducts and Prenylated Flavonoids as Potential PTP1B Inhibitors from Morus nigra. J. Nat. Prod. 2021, 84, 2303–2311. [Google Scholar] [CrossRef]
- Moraes-Vieira, P.M.; Saghatelian, A.; Kahn, B.B. GLUT4 Expression in Adipocytes Regulates De Novo Lipogenesis and Levels of a Novel Class of Lipids With Antidiabetic and Anti-inflammatory Effects. Diabetes 2016, 65, 1808–1815. [Google Scholar] [CrossRef]
- Hu, X.; Wu, J.W.; Zhang, X.D.; Zhao, Q.S.; Huang, J.M.; Wang, H.Y.; Hou, A.J. Isoprenylated flavonoids and adipogenesis-promoting constituents from Morus nigra. J. Nat. Prod. 2011, 74, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jiang, Y.; Yang, J.; He, J.; Sun, J.; Chen, F.; Zhang, M.; Yang, B. Prenylated flavonoids, promising nutraceuticals with impressive biological activities. Trends Food Sci. Technol. 2015, 44, 93–104. [Google Scholar] [CrossRef]
- Chen, X.; Mukwaya, E.; Wong, M.S.; Zhang, Y. A systematic review on biological activities of prenylated flavonoids. Pharm. Biol. 2014, 52, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, D.F.; Selmin, O.I. Flavonoids and cancer prevention: A review of the evidence. J. Nutr. Gerontol. Geriatr. 2012, 31, 206–238. [Google Scholar] [CrossRef]
- Zakaria, I.; Ahmat, N.; Jaafar, F.M.; Widyawaruyanti, A. Flavonoids with antiplasmodial and cytotoxic activities of Macaranga triloba. Fitoterapia 2012, 83, 968–972. [Google Scholar] [CrossRef]
- Cho, N.; Choi, J.H.; Yang, H.; Jeong, E.J.; Lee, K.Y.; Kim, Y.C.; Sung, S.H. Neuroprotective and anti-inflammatory effects of flavonoids isolated from Rhus verniciflua in neuronal HT22 and microglial BV2 cell lines. Food Chem. Toxicol. 2012, 50, 1940–1945. [Google Scholar] [CrossRef]
- Pedro, M.; Ferreira, M.M.; Cidade, H.; Kijjoa, A.; Bronze-da-Rocha, E.; Nascimento, M.S. Artelastin is a cytotoxic prenylated flavone that disturbs microtubules and interferes with DNA replication in MCF-7 human breast cancer cells. Life Sci. 2005, 77, 293–311. [Google Scholar] [CrossRef]
- Pedro, M.; Cerqueira, F.; Sousa, M.E.; Nascimento, M.S.; Pinto, M. Xanthones as inhibitors of growth of human cancer cell lines and their effects on the proliferation of human lymphocytes in vitro. Bioorg Med. Chem. 2002, 10, 3725–3730. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, L.; Liu, X.; Wang, F.; An, Y.; Zhao, W.; Tian, J.; Kong, D.; Zhang, W.; Xu, Y.; et al. The Genus Broussonetia: An Updated Review of Phytochemistry, Pharmacology and Applications. Molecules 2022, 27, 5344. [Google Scholar] [CrossRef]
- Etti, I.C.; Rasedee, A.; Hashim, N.M.; Abdul, A.B.; Kadir, A.; Yeap, S.K.; Waziri, P.; Malami, I.; Lim, K.L.; Etti, C.J. Artonin E induces p53-independent G1 cell cycle arrest and apoptosis through ROS-mediated mitochondrial pathway and livin suppression in MCF-7 cells. Drug Des. Devel Ther. 2017, 11, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Tanjung, M.; Hakim, E.H.; Elfahmi; Latip, J.; Syah, Y.M. Dihydroflavonol and Flavonol Derivatives from Macaranga recurvata. Nat. Product. Commun. 2012, 7, 1934578X1200701013. [Google Scholar] [CrossRef]
- Liu, Y.P.; Sun, L.L.; Zhang, X.L.; Niu, H.Y.; Pan, Z.H.; Fu, Y.H. Prenylated isoflavones with potential antiproliferative activities from Mappianthus iodoides. Nat. Prod. Res. 2020, 34, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Kuang, X.-D.; He, X.-R.; Ren, G.; Wang, Y.; Xu, L.-Y.; Feng, L.-H.; Wang, B.; Zhou, Z.-W. Prenylflavonoids from the Twigs of Artocarpus nigrifolius. Chem. Pharm. Bull. 2018, 66, 434–438. [Google Scholar] [CrossRef]
- Ramli, F.; Rahmani, M.; Ismail, I.S.; Sukari, M.A.; Abd Rahman, M.; Zajmi, A.; Akim, A.M.; Hashim, N.M.; Go, R. A New Bioactive Secondary Metabolite from Artocarpus elasticus. Nat. Prod. Commun. 2016, 11, 1103–1106. [Google Scholar] [CrossRef]
- Yao, J.; Wang, Z.; Wang, R.; Wang, Y.; Xu, J.; He, X. Anti-proliferative and anti-inflammatory prenylated isoflavones and coumaronochromones from the fruits of Ficus altissima. Bioorganic Chem. 2021, 113, 104996. [Google Scholar] [CrossRef]
- Zelová, H.; Hanáková, Z.; Čermáková, Z.; Šmejkal, K.; Dalĺ Acqua, S.; Babula, P.; Cvačka, J.; Hošek, J. Evaluation of anti-inflammatory activity of prenylated substances isolated from Morus alba and Morus nigra. J. Nat. Prod. 2014, 77, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, A.; Khan, Z.; Armaghan, M.; Khan, K.; Sönmez Gürer, E.; Abdull Razis, A.F.; Modu, B.; Almarhoon, Z.M.; Setzer, W.N.; Sharifi-Rad, J. Exploring the therapeutic and anti-tumor properties of morusin: A review of recent advances. Front. Mol. Biosci. 2023, 10, 1168298. [Google Scholar] [CrossRef] [PubMed]
- Pu, H.; Cao, D.; Zhou, X.; Li, F.; Wang, L.; Wang, M. Isoprenylated Flavonoids and 2-Arylbenzofurans from the Root Bark of Morus alba L. and Their Cytotoxic Activity against HGC27 Cancer Cells. Molecules 2024, 29, 30. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Thakur, A.; Pan, G.; Yan, J.; Gaurav, I.; Thakur, S.; Yang, Z.; Cili, A.; Zhang, K. Morus alba derived Kuwanon-A combined with 5-fluorouracil reduce tumor progression via synergistic activation of GADD153 in gastric cancer. MedComm. Oncol. 2023, 2, e24. [Google Scholar] [CrossRef]
- Dat, N.T.; Binh, P.T.; Quynh, L.T.P.; Van Minh, C.; Huong, H.T.; Lee, J.J. Cytotoxic prenylated flavonoids from Morus alba. Fitoterapia 2010, 81, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Ramli, F.; Rahmani, M.; Kassim, N.K.; Hashim, N.M.; Sukari, M.A.; Akim, A.M.; Go, R. New diprenylated dihyrochalcones from leaves of Artocarpus elasticus. Phytochem. Lett. 2013, 6, 582–585. [Google Scholar] [CrossRef]
- Dej-Adisai, S.; Meechai, I.; Puripattanavong, J.; Kummee, S. Antityrosinase and antimicrobial activities from Thai medicinal plants. Arch. Pharm. Res. 2014, 37, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Meenu, M.T.; Kaul, G.; Shukla, M.; Radhakrishnan, K.V.; Chopra, S. Cudraflavone C from Artocarpus hirsutus as a Promising Inhibitor of Pathogenic, Multidrug-Resistant S. aureus, Persisters, and Biofilms: A New Insight into a Rational Explanation of Traditional Wisdom. J. Nat. Prod. 2021, 84, 2700–2708. [Google Scholar] [CrossRef] [PubMed]
- Meenu, M.T.; Kaul, G.; Akhir, A.; Shukla, M.; Radhakrishnan, K.V.; Chopra, S. Developing the Natural Prenylflavone Artocarpin from Artocarpus hirsutus as a Potential Lead Targeting Pathogenic, Multidrug-Resistant Staphylococcus aureus, Persisters and Biofilms with No Detectable Resistance. J. Nat. Prod. 2022, 85, 2413–2423. [Google Scholar] [CrossRef]
- Soekamto, N.H.; Syah, Y.M. Prenylated flavon and antibacterial activities of Artocarpus lanceifolius Roxb bark. J. Phys. Conf. Ser. 2019, 1341, 072014. [Google Scholar] [CrossRef]
- Hu, Q.-F.; Yao, S.; Ma, Y.-Y.; Xiong, R.-F.; Kong, G.-H.; Wu, Y.-P.; Zhao, G.-K.; Dong, M.; Wang, W.-G.; Zhou, M.; et al. Prenylated flavonoids isolated from the twigs of Artocarpus champeden as potential activators for tobacco powdery mildew and their mode of actions. Chem. Biol. Technol. Agric. 2023, 10, 94. [Google Scholar] [CrossRef]
- Boonphong, S.; Baramee, A.; Kittakoop, P.; Puangsombat, P. Antitubercular and Antiplasmodial Prenylated Flavones from the Roots of Artocarpus altilis. Chiang Mai J. Sci. 2007, 34, 339–344. [Google Scholar]
- Widyawaruyanti, A.; Subehan; Kalauni, S.K.; Awale, S.; Nindatu, M.; Zaini, N.C.; Syafruddin, D.; Asih, P.B.S.; Tezuka, Y.; Kadota, S. New prenylated flavones from Artocarpus champeden, and their antimalarial activity in vitro. J. Nat. Med. 2007, 61, 410–413. [Google Scholar] [CrossRef]
- Mustapha, I.; Hakim, E.; Juliawaty, L.; Syah, Y.; Achmad, S. Prenylated flavones from some Indonesian Artocarpus and their antimalarial properties. Med. Plants 2010, 2, 157–160. [Google Scholar] [CrossRef]
- Suhartati, T.; Yandri; Suwandi, J.F.; Hadi, S. In vitro and in vivo antiplasmodial activity of oxyresveratrol and artonine isolated from two Artocarpus plants in Indonesia. Orient. J. Chem. 2010, 26, 825–830. [Google Scholar]
- Govender, N.; Mohd Kaspi, S.N.A.; Krishnan, T.; Mohamed-Hussein, Z.-A. In Silico Screening of Breadfruit (Artocarpus altilis) Prenylated Flavonoids Identify Potential SARS-CoV Inhibitors. Pertanika J. Sci. Technol. 2023, 31, 2145–2159. [Google Scholar] [CrossRef]
- Sohn, H.Y.; Son, K.H.; Kwon, C.S.; Kwon, G.S.; Kang, S.S. Antimicrobial and cytotoxic activity of 18 prenylated flavonoids isolated from medicinal plants: Morus alba L., Morus mongolica Schneider, Broussonetia papyrifera (L.) Vent, Sophora flavescens Ait and Echinosophora koreensis Nakai. Phytomedicine 2004, 11, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Peniche-Pavía, H.A.; Vera-Ku, M.; Peraza-Sánchez, S.R. Phytochemical and Pharmacological Studies on Species of Dorstenia Genus. J. Mex. Chem. Soc. 2018, 62, 9–23. [Google Scholar] [CrossRef]
- Kuete, V.; Simo, I.K.; Ngameni, B.; Bigoga, J.D.; Watchueng, J.; Kapguep, R.N.; Etoa, F.-X.; Tchaleu, B.N.; Beng, V.P. Antimicrobial activity of the methanolic extract, fractions and four flavonoids from the twigs of Dorstenia angusticornis Engl. (Moraceae). J. Ethnopharmacol. 2007, 112, 271–277. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Ngameni, B.; Kuete, V.; Simo, I.K.; Ambassa, P.; Roy, R.; Bezabih, M.; Etoa, F.X.; Ngadjui, B.T.; Abegaz, B.M.; et al. Antimicrobial activity of the crude extracts and five flavonoids from the twigs of Dorstenia barteri (Moraceae). J. Ethnopharmacol. 2008, 116, 483–489. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Li, T.; Liu, X.; Hao, Z.; Ding, S.; Panichayupakaranant, P.; Zhu, K.; Shen, J. Plant Natural Flavonoids Against Multidrug Resistant Pathogens. Adv. Sci. 2021, 8, e2100749. [Google Scholar] [CrossRef] [PubMed]
- Dzoyem, J.P.; Hamamoto, H.; Ngameni, B.; Ngadjui, B.T.; Sekimizu, K. Antimicrobial action mechanism of flavonoids from Dorstenia species. Drug Discov. Ther. 2013, 7, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Polbuppha, I.; Suthiphasilp, V.; Maneerat, T.; Charoensup, R.; Limtharakul, T.; Cheenpracha, S.; Pyne, S.G.; Laphookhieo, S. Macluracochinones A-E, antimicrobial flavonoids from Maclura cochinchinensis (Lour.) Corner. Phytochemistry 2021, 187, 112773. [Google Scholar] [CrossRef] [PubMed]
- Özçelik, B.; Orhan, I.; Toker, G. Antiviral and antimicrobial assessment of some selected flavonoids. Z. Naturforsch C J. Biosci. 2006, 61, 632–638. [Google Scholar] [CrossRef]
- Pang, D.; Wang, W.; Li, E.; Shen, W.; Mu, L.; Liao, S.; Liu, F. Sanggenon D from root bark of mulberry inhibits the growth of Staphylococcus aureus by moderating the fatty acid biosynthesis system. Ind. Crop. Prod. 2019, 140, 111719. [Google Scholar] [CrossRef]
- Čulenová, M.; Sychrová, A.; Hassan, S.T.S.; Berchová-Bímová, K.; Svobodová, P.; Helclová, A.; Michnová, H.; Hošek, J.; Vasilev, H.; Suchý, P.; et al. Multiple In vitro biological effects of phenolic compounds from Morus alba root bark. J. Ethnopharmacol. 2020, 248, 112296. [Google Scholar] [CrossRef]
- Wu, S.-C.; Han, F.; Song, M.-R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.-Z. Natural Flavones from Morus alba against Methicillin-Resistant Staphylococcus aureus via Targeting the Proton Motive Force and Membrane Permeability. J. Agric. Food Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, Z.-J.; He, Y.-J.; Qin, Y.; Zhou, Y.; Qi, Z.-H.; Zhou, Z.-S.; Zhu, Y.-Y.; Jin, D.-N.; Chen, S.-S.; et al. Bioguided isolation, identification and bioactivity evaluation of anti-MRSA constituents from Morus alba Linn. J. Ethnopharmacol. 2021, 281, 114542. [Google Scholar] [CrossRef] [PubMed]
- Zuo, G.Y.; Yang, C.X.; Han, J.; Li, Y.Q.; Wang, G.C. Synergism of prenylflavonoids from Morus alba root bark against clinical MRSA isolates. Phytomedicine 2018, 39, 93–99. [Google Scholar] [CrossRef]
- Škovranová, G.; Čulenová, M.; Treml, J.; Dzurická, L.; Marova, I.; Sychrová, A. Prenylated phenolics from Morus alba against MRSA infections as a strategy for wound healing. Front. Pharmacol. 2022, 13, 1068371. [Google Scholar] [CrossRef]
- Aelenei, P.; Rimbu, C.M.; Horhogea, C.E.; Lobiuc, A.; Neagu, A.-N.; Dunca, S.I.; Motrescu, I.; Dimitriu, G.; Aprotosoaie, A.C.; Miron, A. Prenylated phenolics as promising candidates for combination antibacterial therapy: Morusin and kuwanon G. Saudi Pharm. J. 2020, 28, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Wu, D.; Jiang, Y.; Prasad, K.N.; Lin, S.; Jiang, G.; He, J.; Zhao, M.; Luo, W.; Yang, B. Identification of flavonoids in litchi (Litchi chinensis Sonn.) leaf and evaluation of anticancer activities. J. Funct. Foods 2014, 6, 555–563. [Google Scholar] [CrossRef]
- Prakash, O.; Kumar, R.; Mishra, A.; Gupta, R. Artocarpus heterophyllus (Jackfruit): An overview. Pharmacogn. Rev. 2009, 3, 353. [Google Scholar]
- Thongnest, S.; Lhinhatrakool, T.; Wetprasit, N.; Sutthivaiyakit, P.; Sutthivaiyakit, S. Eriosema chinense: A rich source of antimicrobial and antioxidant flavonoids. Phytochemistry 2013, 96, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.M.M.; Silva, A.M.S. The Antioxidant Activity of Prenylflavonoids. Molecules 2020, 25, 696. [Google Scholar] [CrossRef] [PubMed]
- Lathiff, S.M.A.; Jemaon, N.; Abdullah, S.A.; Jamil, S. Flavonoids from Artocarpus anisophyllus and their Bioactivities. Nat. Product. Commun. 2015, 10, 1934578X1501000305. [Google Scholar] [CrossRef]
- Arriffin, N.M.; Jamil, S.; Basar, N.; Khamis, S.; Abdullah, S.A.; Lathiff, S.M.A. Phytochemical Studies and Antioxidant Activities of King. Rec. Nat. Prod. 2017, 11, 299–303. [Google Scholar]
- Omisore, N.O.; Adewunmi, C.O.; Iwalewa, E.O.; Ngadjui, B.T.; Adenowo, T.K.; Abegaz, B.M.; Ojewole, J.A.; Watchueng, J. Antitrichomonal and antioxidant activities of Dorstenia barteri and Dorstenia convexa. Braz. J. Med. Biol. Res. 2005, 38, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.H.; Lu, Y.H.; Yang, S.Z.; Won, S.J.; Lin, C.N. Cytotoxic prenylflavonoids from Artocarpus elasticus. J. Nat. Prod. 2005, 68, 1692–1695. [Google Scholar] [CrossRef]
- Lin, K.-W.; Liu, C.-H.; Tu, H.-Y.; Ko, H.-H.; Wei, B.-L. Antioxidant prenylflavonoids from Artocarpus communis and Artocarpus elasticus. Food Chem. 2009, 115, 558–562. [Google Scholar] [CrossRef]
- Ko, F.N.; Cheng, Z.J.; Lin, C.N.; Teng, C.M. Scavenger and Antioxidant Properties of Prenylflavones Isolated From Artocarpus heterophyllus. Free Radic. Biol. Med. 1998, 25, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.N.; Lu, C.M.; Lin, H.C.; Fang, S.C.; Shieh, B.J.; Hsu, M.F.; Wang, J.P.; Ko, F.N.; Teng, C.M. Novel antiplatelet constituents from formosan moraceous plants. J. Nat. Prod. 1996, 59, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, M.; Manisankar, P.; Gandhidasan, R.; Murugesan, R. Free Radicals Scavenging Efficiency of a Few Naturally Occurring Flavonoids: A Comparative Study. J. Agric. Food Chem. 2004, 52, 7389–7394. [Google Scholar] [CrossRef] [PubMed]
- Hashim, N.; Rahmani, M.; Sukari, M.A.; Ali, A.M.; Alitheen, N.B.; Go, R.; Ismail, H.B. Two new xanthones from Artocarpus obtusus. J. Asian Nat. Prod. Res. 2010, 12, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Ee, G.C.L.; Teo, S.H.; Rahmani, M.; Lim, C.K.; Lim, Y.M.; Go, R. Artomandin, a new xanthone from Artocarpus kemando (Moraceae). Nat. Product. Res. 2011, 25, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Huong, T.T.; Cuong, N.X.; Tram, L.H.; Quang, T.T.; Duong, L.V.; Nam, N.H.; Dat, N.T.; Huong, P.T.; Diep, C.N.; Kiem, P.V.; et al. A new prenylated aurone from Artocarpus altilis. J. Asian Nat. Prod. Res. 2012, 14, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.-C.; Tzeng, C.-W.; Lin, C.-C.; Yen, F.-L.; Ko, H.-H. Prenylated flavonoids from Artocarpus altilis: Antioxidant activities and inhibitory effects on melanin production. Phytochemistry 2013, 89, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.; Nunuk Hariani, S.; Yana, M.s.; Firdaus, F. Antibacterial Activity of Artocarpus Integer (thunb.) Merr. Fraction by Difusi Agar Method. J. Ind. Has. Perkeb. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- Lee, B.W.; Lee, J.H.; Gal, S.W.; Moon, Y.H.; Park, K.H. Selective ABTS radical-scavenging activity of prenylated flavonoids from Cudrania tricuspidata. Biosci. Biotechnol. Biochem. 2006, 70, 427–432. [Google Scholar] [CrossRef]
- Park, K.H.; Park, Y.D.; Han, J.M.; Im, K.R.; Lee, B.W.; Jeong, I.Y.; Jeong, T.S.; Lee, W.S. Anti-atherosclerotic and anti-inflammatory activities of catecholic xanthones and flavonoids isolated from. Bioorg Med. Chem. Lett. 2006, 16, 5580–5583. [Google Scholar] [CrossRef]
- Kyekyeku, J.O.; Kusari, S.; Adosraku, R.K.; Zuhlke, S.; Spiteller, M. Prenylated 2-arylbenzofuran derivatives with potent antioxidant properties from Chlorophora regia (Moraceae). Fitoterapia 2016, 108, 41–47. [Google Scholar] [CrossRef]
- Dufall, K.G.; Ngadjui, B.T.; Simeon, K.F.; Abegaz, B.M.; Croft, K.D. Antioxidant activity of prenylated flavonoids from the West African medicinal plant Dorstenia mannii. J. Ethnopharmacol. 2003, 87, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Fongang, Y.S.F.; Bankeu, J.J.K.; Ali, M.S.; Awantu, A.F.; Zeeshan, A.; Assob, C.N.; Mehreen, L.; Lenta, B.N.; Ngouela, S.A.; Tsamo, E. Flavonoids and other bioactive constituents from Ficus thonningii Blume (Moraceae). Phytochem. Lett. 2015, 11, 139–145. [Google Scholar] [CrossRef]
- Martins, B.A.; Sande, D.; Solares, M.D.; Takahashi, J.A. Antioxidant role of morusin and mulberrofuran B in ethanol extract of Morus alba roots. Nat. Prod. Res. 2021, 35, 5993–5996. [Google Scholar] [CrossRef]
- Laskin, D.L.; Sunil, V.R.; Gardner, C.R.; Laskin, J.D. Macrophages and Tissue Injury: Agents of Defense or Destruction? Annu. Rev. Pharmacol. Toxicol. 2011, 51, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Hanáková, Z.; Hošek, J.; Babula, P.; Dall’Acqua, S.; Václavík, J.; Šmejkal, K. C-Geranylated Flavanones from Paulownia tomentosa Fruits as Potential Anti-inflammatory Compounds Acting via Inhibition of TNF-alpha Production. J. Nat. Prod. 2015, 78, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Hanáková, Z.; Hošek, J.; Kutil, Z.; Temml, V.; Landa, P.; Vaněk, T.; Schuster, D.; Dall’Acqua, S.; Cvačka, J.; Polanský, O.; et al. Anti-inflammatory Activity of Natural Geranylated Flavonoids: Cyclooxygenase and Lipoxygenase Inhibitory Properties and Proteomic Analysis. J. Nat. Prod. 2017, 80, 999–1006. [Google Scholar] [CrossRef]
- Ko, W.; Liu, Z.; Kim, K.W.; Dong, L.; Lee, H.; Kim, N.Y.; Lee, D.S.; Woo, E.R. Kuwanon T and Sanggenon a Isolated from Morus alba Exert Anti-Inflammatory Effects by Regulating NF-kappaB and HO-1/Nrf2 Signaling Pathways in BV2 and RAW264.7 Cells. Molecules 2021, 26, 7642. [Google Scholar] [CrossRef]
- Nishikawa, S.; Inoue, Y.; Hori, Y.; Miyajima, C.; Morishita, D.; Ohoka, N.; Hida, S.; Makino, T.; Hayashi, H. Anti-Inflammatory Activity of Kurarinone Involves Induction of HO-1 via the KEAP1/Nrf2 Pathway. Antioxidants 2020, 9, 842. [Google Scholar] [CrossRef]
- Owor, R.O.; Bedane, K.G.; Zuhlke, S.; Derese, S.; Ong’amo, G.O.; Ndakala, A.; Spiteller, M. Anti-inflammatory Flavanones and Flavones from Tephrosia linearis. J. Nat. Prod. 2020, 83, 996–1004. [Google Scholar] [CrossRef]
- Ryu, H.W.; Park, M.H.; Kwon, O.K.; Kim, D.Y.; Hwang, J.Y.; Jo, Y.H.; Ahn, K.S.; Hwang, B.Y.; Oh, S.R. Anti-inflammatory flavonoids from root bark of Broussonetia papyrifera in LPS-stimulated RAW264.7 cells. Bioorg Chem. 2019, 92, 103233. [Google Scholar] [CrossRef] [PubMed]
- Shahinozzaman, M.; Taira, N.; Ishii, T.; Halim, M.A.; Hossain, M.A.; Tawata, S. Anti-Inflammatory, Anti-Diabetic, and Anti-Alzheimer’s Effects of Prenylated Flavonoids from Okinawa Propolis: An Investigation by Experimental and Computational Studies. Molecules 2018, 23, 2479. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, F.; Fan, L.; Zhang, W.; Wang, T.; Du, Y.; Bai, X. Baicalin alleviates atherosclerosis by relieving oxidative stress and inflammatory responses via inactivating the NF-kappaB and p38 MAPK signaling pathways. Biomed. Pharmacother. 2018, 97, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-A.; Fang, S.-C.; Wu, C.-H.; Huang, S.-M.; Yen, G.-C. Anti-inflammatory Effect of the 5,7,4′-Trihydroxy-6-geranylflavanone Isolated from the Fruit of Artocarpus communis in S100B-Induced Human Monocytes. J. Agric. Food Chem. 2011, 59, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Gantner, B.N.; LaFond, K.M.; Bonini, M.G. Nitric oxide in cellular adaptation and disease. Redox Biol. 2020, 34, 101550. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.B.; Chen, Z.R.; Yang, S.L.; Hong, F.F. Nitric Oxide Synthases in Rheumatoid Arthritis. Molecules 2023, 28, 4414. [Google Scholar] [CrossRef] [PubMed]
- Passante, E.; Ehrhardt, C.; Sheridan, H.; Frankish, N. RBL-2H3 cells are an imprecise model for mast cell mediator release. Inflamm. Res. 2009, 58, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Han, A.-R.; Kang, Y.-J.; Windono, T.; Lee, S.K.; Seo, E.-K. Prenylated Flavonoids from the Heartwood of Artocarpus communis with Inhibitory Activity on Lipopolysaccharide-Induced Nitric Oxide Production. J. Nat. Prod. 2006, 69, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Guo, J.M.; Yan, G.; Zhang, M.M.; Zhang, W.H.; Qiang, L.; Fu, Y.H. Anti-Inflammatory and Antiproliferative Prenylated Isoflavone Derivatives from the Fruits of Ficus carica. J. Agric. Food Chem. 2019, 67, 4817–4823. [Google Scholar] [CrossRef]
- Lee, T.; Kwon, J.; Lee, D.; Mar, W. Effects of Cudrania tricuspidata Fruit Extract and Its Active Compound, 5,7,3′,4′-Tetrahydroxy-6,8-diprenylisoflavone, on the High-Affinity IgE Receptor-Mediated Activation of Syk in Mast Cells. J. Agric. Food Chem. 2015, 63, 5459–5467. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed]
- Arung, E.T.; Shimizu, K.; Kondo, R. Inhibitory effect of isoprenoid-substituted flavonoids isolated from Artocarpus heterophyllus on melanin biosynthesis. Planta Med. 2006, 72, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Arung, E.T.; Shimizu, K.; Kondo, R. Structure-activity relationship of prenyl-substituted polyphenols from Artocarpus heterophyllus as inhibitors of melanin biosynthesis in cultured melanoma cells. Chem. Biodivers. 2007, 4, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Kondo, R.; Sakai, K.; Takeda, N.; Nagahata, T. The skin-lightening effects of artocarpin on UVB-induced pigmentation. Planta Med. 2002, 68, 79–81. [Google Scholar] [CrossRef]
- Nguyen, M.T.T.; Le, T.H.; Nguyen, H.X.; Dang, P.H.; Do, T.N.V.; Abe, M.; Takagi, R.; Nguyen, N.T. Artocarmins G–M, Prenylated 4-Chromenones from the Stems of Artocarpus rigida and Their Tyrosinase Inhibitory Activities. J. Nat. Prod. 2017, 80, 3172–3178. [Google Scholar] [CrossRef] [PubMed]
- Dej-Adisai, S.; Parndaeng, K.; Wattanapiromsakul, C.; Hwang, J.S. Three New Isoprenylated Flavones from Artocarpus chama Stem and Their Bioactivities. Molecules 2021, 27, 3. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef]
- Lee, D.; Bhat, K.P.; Fong, H.H.; Farnsworth, N.R.; Pezzuto, J.M.; Kinghorn, A.D. Aromatase inhibitors from Broussonetia papyrifera. J. Nat. Prod. 2001, 64, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wu, J.-W.; Wang, M.; Yu, M.-H.; Zhao, Q.-S.; Wang, H.-Y.; Hou, A.-J. 2-Arylbenzofuran, Flavonoid, and Tyrosinase Inhibitory Constituents of Morus yunnanensis. J. Nat. Prod. 2012, 75, 82–87. [Google Scholar] [CrossRef]
- Jeong, S.H.; Ryu, Y.B.; Curtis-Long, M.J.; Ryu, H.W.; Baek, Y.S.; Kang, J.E.; Lee, W.S.; Park, K.H. Tyrosinase Inhibitory Polyphenols from Roots of Morus lhou. J. Agric. Food Chem. 2009, 57, 1195–1203. [Google Scholar] [CrossRef]
- Lee, N.K.; Son, K.H.; Chang, H.W.; Kang, S.S.; Park, H.; Heo, M.Y.; Kim, H.P. Prenylated flavonoids as tyrosinase inhibitors. Arch. Pharm. Res. 2004, 27, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tao, G.; Chen, J.; Zheng, Z.P. Characterization of a New Flavone and Tyrosinase Inhibition Constituents from the Twigs of Morus alba L. Molecules 2016, 21, 1130. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Tang, G.H.; Lou, L.L.; Li, W.; Zhang, B.; Liu, B.; Yin, S. Prenylated flavonoids as potent phosphodiesterase-4 inhibitors from Morus alba: Isolation, modification, and structure-activity relationship study. Eur. J. Med. Chem. 2018, 144, 758–766. [Google Scholar] [CrossRef]
- Savonenko, A.V.; Wong, P.C.; Li, T. Chapter 19—Alzheimer diseases. In Neurobiology of Brain Disorders, 2nd ed.; Zigmond, M.J., Wiley, C.A., Chesselet, M.-F., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 313–336. [Google Scholar]
- Kuk, E.B.; Jo, A.R.; Oh, S.I.; Sohn, H.S.; Seong, S.H.; Roy, A.; Choi, J.S.; Jung, H.A. Anti-Alzheimer’s disease activity of compounds from the root bark of Morus alba L. Arch. Pharm. Res. 2017, 40, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, W.S.; Kim, Y.S.; Curtis-Long, M.J.; Lee, B.W.; Ryu, Y.B.; Park, K.H. Isolation of Cholinesterase-Inhibiting Flavonoids from Morus lhou. J. Agric. Food Chem. 2011, 59, 4589–4596. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-S.; Ko, W.; Kim, D.-C.; Kim, Y.-C.; Jeong, G.-S. Cudarflavone B Provides Neuroprotection against Glutamate-Induced Mouse Hippocampal HT22 Cell Damage through the Nrf2 and PI3K/Akt Signaling Pathways. Molecules 2014, 19, 10818–10831. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.; Yoon, C.-S.; Kim, K.-W.; Lee, H.; Kim, N.; Woo, E.-R.; Kim, Y.-C.; Kang, D.G.; Lee, H.S.; Oh, H.; et al. Neuroprotective and Anti-Inflammatory Effects of Kuwanon C from Cudrania tricuspidata Are Mediated by Heme Oxygenase-1 in HT22 Hippocampal Cells, RAW264.7 Macrophage, and BV2 Microglia. Int. J. Mol. Sci. 2020, 21, 4839. [Google Scholar] [CrossRef]
- Hiep, N.T.; Kwon, J.; Kim, D.-W.; Hwang, B.Y.; Lee, H.-J.; Mar, W.; Lee, D. Isoflavones with neuroprotective activities from fruits of Cudrania tricuspidata. Phytochemistry 2015, 111, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Hiep, N.T.; Kim, D.-W.; Hong, S.; Guo, Y.; Hwang, B.Y.; Lee, H.J.; Mar, W.; Lee, D. Chemical Constituents Isolated from the Root Bark of Cudrania tricuspidata and Their Potential Neuroprotective Effects. J. Nat. Prod. 2016, 79, 1938–1951. [Google Scholar] [CrossRef]
- Groh, J.; Martini, R. Neuroinflammation as modifier of genetically caused neurological disorders of the central nervous system: Understanding pathogenesis and chances for treatment. Glia 2017, 65, 1407–1422. [Google Scholar] [CrossRef]
- Kim, K.-W.; Quang, T.H.; Ko, W.; Kim, D.-C.; Yoon, C.-S.; Oh, H.; Kim, Y.-C. Anti-neuroinflammatory effects of cudraflavanone A isolated from the chloroform fraction of Cudrania tricuspidata root bark. Pharm. Biol. 2018, 56, 192–200. [Google Scholar] [CrossRef]
- Lewerenz, J.; Ates, G.; Methner, A.; Conrad, M.; Maher, P. Oxytosis/Ferroptosis—(Re-) Emerging Roles for Oxidative Stress-Dependent Non-apoptotic Cell Death in Diseases of the Central Nervous System. Front. Neurosci. 2018, 12, 214. [Google Scholar] [CrossRef]
- Wen, L.; Shi, D.; Zhou, T.; Tu, J.; He, M.; Jiang, Y.; Yang, B. Identification of two novel prenylated flavonoids in mulberry leaf and their bioactivities. Food Chem. 2020, 315, 126236. [Google Scholar] [CrossRef]
- Ye, X.S.; Tian, W.J.; Zhou, M.; Zeng, D.Q.; Lin, T.; Wang, G.H.; Yao, X.S.; Chen, H.F. Prenylated flavonoids from Ficus hirta induces HeLa cells apoptosis via MAPK and AKT signaling pathways. Bioorg Med. Chem. Lett. 2021, 38, 127859. [Google Scholar] [CrossRef]
- Nie, J.; Luo, Y.; Huang, X.N.; Gong, Q.H.; Wu, Q.; Shi, J.S. Icariin inhibits beta-amyloid peptide segment 25-35 induced expression of beta-secretase in rat hippocampus. Eur. J. Pharmacol. 2010, 626, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.W.; Ko, H.; Yang, H.O.; Wang, X.M. Icariin attenuates beta-amyloid-induced neurotoxicity by inhibition of tau protein hyperphosphorylation in PC12 cells. Neuropharmacology 2010, 59, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Cidade, H.M.; Nacimento, M.S.; Pinto, M.M.; Kijjoa, A.; Silva, A.M.; Herz, W. Artelastocarpin and carpelastofuran, two new flavones, and cytotoxicities of prenyl flavonoids from Artocarpus elasticus against three cancer cell lines. Planta Med. 2001, 67, 867–870. [Google Scholar] [CrossRef] [PubMed]
- Hakim, E.H.; Achmad, S.A.; Juliawaty, L.D.; Makmur, L.; Syah, Y.M.; Aimi, N.; Kitajima, M.; Takayama, H.; Ghisalberti, E.L. Prenylated flavonoids and related compounds of the Indonesian Artocarpus (Moraceae). J. Nat. Med. 2006, 60, 161–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.C.; Wang, S.; Wu, Y.; Chen, R.Y.; Yu, D.Q. Five new diprenylated flavonols from the leaves of Broussonetia kazinoki. J. Nat. Prod. 2001, 64, 1206–1209. [Google Scholar] [CrossRef]
- Hošek, J.; Bartos, M.; Chudík, S.; Dall’Acqua, S.; Innocenti, G.; Kartal, M.; Kokoška, L.; Kollár, P.; Kutil, Z.; Landa, P.; et al. Natural Compound Cudraflavone B Shows Promising Anti-inflammatory Properties in Vitro. J. Nat. Prod. 2011, 74, 614–619. [Google Scholar] [CrossRef]
- Daus, M.; Chaithada, P.; Phongpaichit, S.; Watanapokasin, R.; Carroll, A.R.; Mahabusarakam, W. New prenylated dihydrochalcones from the leaves of Artocarpus elasticus. Phytochem. Lett. 2017, 19, 226–230. [Google Scholar] [CrossRef]
- Abdel Bar, F.M.; Abbas, G.M.; Gohar, A.A.; Lahloub, M.I. Antiproliferative activity of stilbene derivatives and other constituents from the stem bark of Morus nigra L. Nat. Prod. Res. 2020, 34, 3506–3513. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
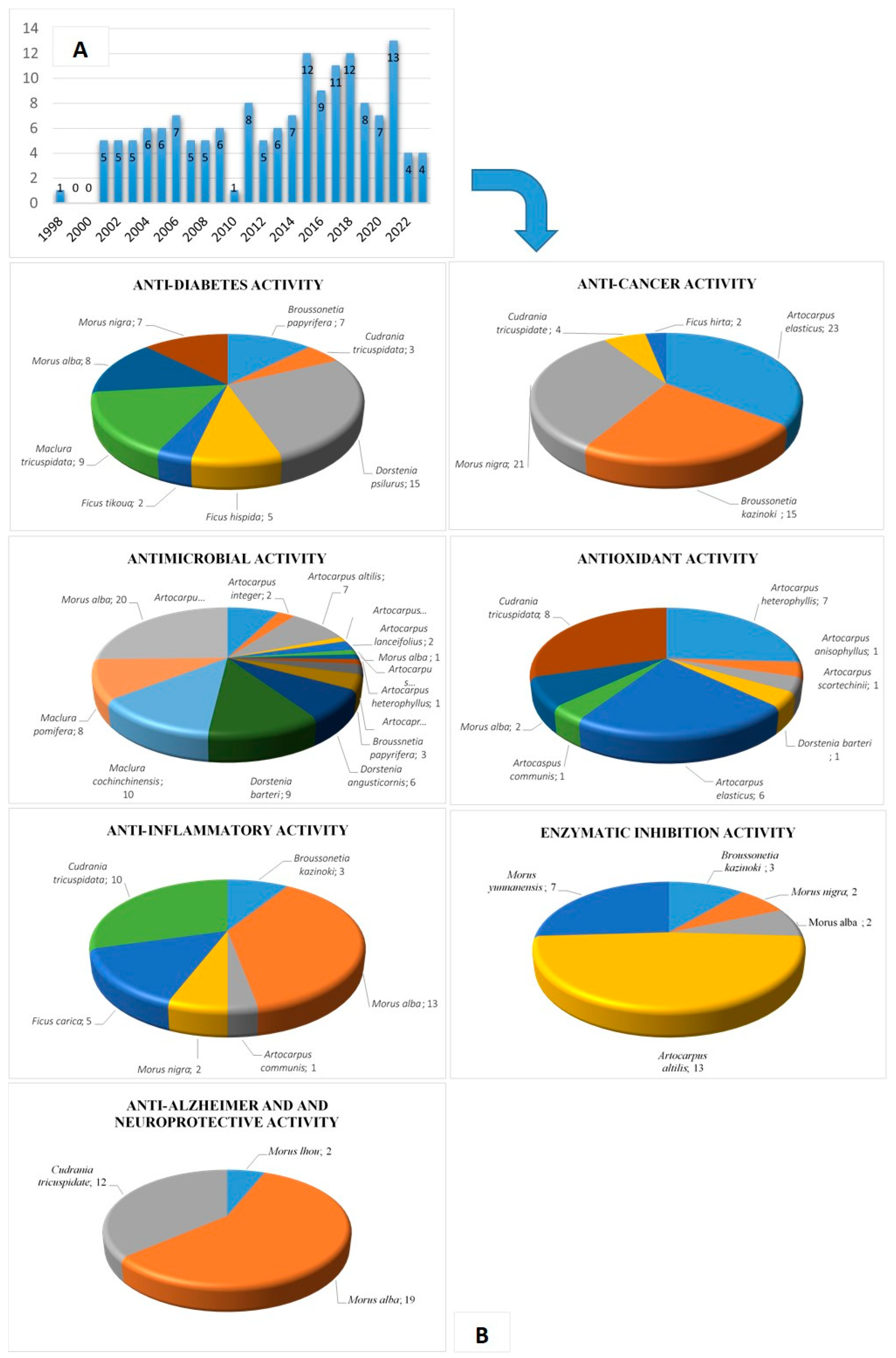{kind=link}
| Compounds | Measured Parameters | Species | Biological Activities | References | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 12-hydroxyartonin E | IC50 = 0.9 µg mL−1 | Artocarpus rigida | AM | [66] | ||||||
| Cudraflavone A | Tyrosinase inhibition: IC50 = 88.4 µM | Artocarpus altilis | EI | [98] | ||||||
| (2R)-Cudraflavanone H | Against MPP+-induced cell death: EC50 > 50 µM | Cudrania tricuspidata | AA | [141] | ||||||
| Against OGD-induced neurotoxicity: EC50 > 50 µM | AA | |||||||||
| Against 6-OHDA-induced cell death: EC50 > 50 µM | AA | |||||||||
| (2R)-ficusflavonoid A | Hela, MCF-7, HepG2, H460: IC50 > 40 µM | Ficus hirta | AC | [146] | ||||||
| (2R)/(2S)-euchrenone a7 | IC50 = 6.28 ± 1.01 µM | Morus alba | AD | [31] | ||||||
| (2S)-2′,4′-dihydroxy-7-methoxy-8-prenylflavan | Tyrosinase inhibition: IC50 = 0.81 µM | Morus yunnanensis | EI | [130] | ||||||
| (2S)-7,2′-dihydroxy-4′-methoxy-8-prenylflavan | Tyrosinase inhibition: IC50 > 100 µM | EI | [130] | |||||||
| (2S)-7,4′-dihydroxy-8-prenylflavan | Tyrosinase inhibition: IC50 > 100 µM | EI | [130] | |||||||
| (2S)-cudraflavanone H | Against MPP+-induced cell death: EC50 > 50 µM Against OGD-induced neurotoxicity: EC50 > 50 µM Against 6-OHDA-induced cell death: EC50 > 50 µM | Cudrania tricuspidata | AA | [141] | ||||||
| (2S)-Ficusflavonid A | Hela, MCF-7, HepG2, H460: IC50 > 40 µM | Ficus hirta | AC | [146] | ||||||
| 4ʹ-hydroxy-8-methoxy-6-(4-methyl-1H-pyrrol-2-yl)-flavone | C50 = 51.5 μg mL−1 IR = 88.3% ± 6.2 µM | Artocarpus champeden | AM | [62] | ||||||
| 10-Oxoartogomezianone | Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | ||||||
| 14-hydroxyartonin E | ND | Artocarpus lanceifolius | AM | [61] | ||||||
| 2-[{3-hydroxy-2′,2-dimethyl-8-(3-methyl-2-butenyl)}chroman-6-yl]-7-hydroxy-8-(3-methyl-2-butenyl)-chroman-4-one | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [121] | ||||||
| 2″,2″-dimethylpyran-(5″,6″:6,7)-5,4′-dihydroxy-4′-methoxy-flavonol | IR = >65% | Artocarpus champeden | AM | [62] | ||||||
| 3′-(3-methylbut-2-en-1-yl) biochanin A | IC50 = 22.1 ± 7.6 µM | Ficus hispida | AD | [28] | ||||||
| 3′-(3-methylbut-2-enyl)-3′,4′,7-trihydroxyflavane | IC50 = 75.7 ± 2.0 µM | Broussonetia papyrifera | AD | [25] | ||||||
| 3′-geranyl-3-prenyl-2′,4′,5,7-tetrahydroxyflavone | HeLa: IC50 = 1.32 ± 0.51 µM MCF-7: IC50 = 3.92 ± 0.91 µM | Morus alba | AC | [56] | ||||||
| 4-hydroxylonchocarpin | MIC = 1.2 µg mL−1 MIC = 16 μg mL−1 | Dorstenia barteri | AM | [71] | ||||||
| 4′,5,7-trihydroxy-6-[(1R*,6R*)-3-methyl-6-(1-methylethenyl)cyclohex-2-en-1-yl] isoflavone | ND | Ficus hispida | AD | [28] | ||||||
| 4′-hydroxy-5,7-dimethoxy-6-(3-methyl-2-buteny)-isoflavone | Inhibition of NO production: IC50 = 2.46 µM | Ficus carica | AI | [28] | ||||||
| 4′-O-methylalpinumisoflavone | Inhibition of β-hexosaminidase release: IC50 > 40 | Cudrania tricuspidata | AI | [62] | ||||||
| 4′-hydroxy-8-methoxy-6-(4-methylfuran-2-yl)-flavone) | IR = 65% | Artocarpus champeden | AM | [62] | ||||||
| 5,7-dihydroxy-6-(2″-hydroxy-3″-methylbut-3″-enyl)-4′-methoxylisoflavone | ND | Maclura tricuspidata | AD | [30] | ||||||
| 5,7,3′,4′-tetrahydroxy-6,8-diprenylisoflavone | Inhibition of β-hexosaminidase release: IC50 = 20.4 µM | Cudrania tricuspidata | AI | [62] | ||||||
| 5,7,4′-trihydroxy-6-geranylflavanone | Inhibition of TNF-α and IL-1β: IC50 ≤ 2.5 µM | Artocarpus communis | AI | [115] | ||||||
| 5,7,4′-trihydroxy-6,8-diprenylisoflavone | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| 5′-geranyl-4′-methoxy-5,7,2′-trihydroxyflavone | AChE inhibition: IC50 = 3.1 µM BChE inhibition: IC50 = 1.74 µM | Morus alba | AA | [136,147,148] | ||||||
| 5′-geranyl-5,7,2′,4′-tetrahydroxy-flavone | BChE inhibition: IC50 = 16.21 µM | AA | [136,147,148] | |||||||
| MIC =2–4 μg mL−1 MBC = 2–8 μg mL−1 | AM | [82] | ||||||||
| 5RE4-artocarpin | MBA = −6.6 kcal mol−1 | Artocarpus altilis | AM | [67] | ||||||
| 5RE4-artoindonesianin V | MBA = −6.6 kcal mol−1 | AM | [67] | |||||||
| 6-[(1R*,6R*)-3-methyl-6-(1-methylethenyl)-2-cyclohexen-1-yl]-5,7,4′-trihydroxyisoflavone | IC50 = 32.5 ± 6.7 µM | Ficus tikoua | AD | [29] | ||||||
| 6-prenylapigenin | MIC = 0.8 μg mL−1 | Dorstenia barteri | AM | [71] | ||||||
| 6,8-diprenyleriodictyol | MIC = 0.5 μg mL−1 | AM | [71] | |||||||
| 7-O-(2,2-dimethylallyl)-aromadendrin | MIC = 32–128 μg mL−1 | Maclura pomifera | AM | [75] | ||||||
| 8-(1,1-dimethylallyl)-5′-(3-methyl-but-2-enyl)-3′,4′,5,7-tetrahydroxyflavonol | IC50 = 3.6 ± 0.4 µM | Broussonetia papyrifera | AD | [25] | ||||||
| 8-Geranyl-3-(hydroxyprenyl) isoetin | Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | ||||||
| 8-Geranylapigenin | Tyrosinase inhibition: IC50 > 300.0 µM | EI | [98] | |||||||
| HeLa: IC50 = 2.24 ± 0.48 µM MCF-7: IC50 = 3.21 ± 0.87 µM | Morus alba | AC | [56] | |||||||
| Albanin T | IC50 = 35.35 ± 0.42 µM | AD | [33] | |||||||
| Albanol B | AChE inhibition: IC50 = 2.47 ± 0.06 BChE inhibition: IC50 = 1.39 ± 0.06 | AA | [136] | |||||||
| Alpinumisoflavone | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| MIC = 4–128 μg mL−1 | Maclura pomifera | AM | [75] | |||||||
| Alpinumisoflavone, 6,8-diprenylgenistein | ND | Maclura tricuspidata | AD | [30] | ||||||
| Amentoflavone | MIC = 0.7 µg mL−1 | Dorstenia barteri | AM | [71] | ||||||
| Angusticornin B | MIC = 78.12 µg mL−1 | Dorstenia angusticornis | AM | [70] | ||||||
| Artelasticin | MCF-7: IC50 = 8.9 ± 2.4 | Artocarpus elasticus | AC | [149] | ||||||
| TK-10: IC50 = 10.6 ± 3.1 µM UACC-62: IC50 = 8.8 ± 0.2 µM | AC | |||||||||
| MCF-7: IC50 = 2.2 ± 0.3 µM | AC | [43] | ||||||||
| MCF-7: IC50 = 2.2 ± 0.3 µM | AC | |||||||||
| TK-10: IC50 = 4.6 ± 1.7 µM | AC | |||||||||
| UACC-62: IC50 = 2.2 ± 0.5 µM | AC | |||||||||
| NCI–H460: IC50 = 5.2 ± 1.5 µM | AC | |||||||||
| SF-268: IC50 = 10.8 ± 1.8 µM | AC | |||||||||
| Artelastocarpin | MCF-7: IC50 = 7.1 ± 0.3 µM TK-10: IC50 = 12.5 ± 0.1 µM UACC-62: IC50 = 8.5 ± 0.3 µM | AC | [149] | |||||||
| Artelastochromene | MCF-7: IC50 = 9.6 ± 0.7 µM TK-10: IC50 = 10.5 ± 1.1 µM UACC-62: IC50 = 10.5 ± 1.7 µM | AC | [43] | |||||||
| Artelastoheterol | IC50 = 24.2 ± 0.8 µM | AO | [90] | |||||||
| Artocarpesin | MCF-7: IC50 = 42.4 ± 1.6 µM TK-10: IC50 = 64.1 ± 5.7 µM UACC-62: IC50 = 38.1 ± 2.4 µM | AC | [149] | |||||||
| Artocarpetin | ND | Artocarpus heterophyllis | AO | [84,85] | ||||||
| Artocarpetin A | ND | AO | ||||||||
| Artocarpin | MIC = 3.12 μg mL−1 | Artocarpus altilis | AM | [98] | ||||||
| MIC = 2 µg mL−1 MIC = 4 µg mL−1 | Artocarpus integer | AM | [58] | |||||||
| Tyrosinase inhibition: IC50 = 270.3 µM | Artocarpus altilis | EI | [98] | |||||||
| ND | Artocarpus heterophyllis | AO | [84] | |||||||
| Artoflavone A | Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | ||||||
| ND | Artocaspus communis | AO | [91] | |||||||
| Artogomezianone | Tyrosinase inhibition: IC50 = 84.8 µM | Artocarpus altilis | EI | [98] | ||||||
| Artonin A | ND | Artocarpus heterophyllis | AO | [85] | ||||||
| Artonin B | ND | AO | ||||||||
| Artonin E | IC50 = 0.2 μg mL−1 | Artocarpus rigida | AM | [66] | ||||||
| P388: IC50 = 0.06 µM | Artocarpus elasticus | AC | [150] | |||||||
| MDA-MB231: IC50 = 9.77 ± 0.50 µM | AC | [46] | ||||||||
| MCF-10A: IC50 = 45.80 ± 3.60 µM | AC | |||||||||
| IC50 = 0.2 µg mL−1 | AM | [66] | ||||||||
| IC50 = 0.1 µg mL−1 | Artocarpus heterophyllus | AM | ||||||||
| IC50 = 0.3 µg mL−1 | Artocarpus lanceifolius | AM | ||||||||
| Atalantoflavone | HeLa: IC50 = 1.25 ± 0.46 µM MCF-7: IC50 = 6.54 ± 1.23 µM | Morus alba | AC | AI | [56] | |||||
| Auriculatin | MIC = 1–8 μg mL−1 | Maclura pomifera | AM | [75] | ||||||
| Bartericin | ND | Dorstenia barteri | AO | [70] | ||||||
| Bartericin A | MIC = 0.31 µg mL−1 | Dorstenia angusticornis | AM | [70] | ||||||
| MIC = 39.06 µg mL−1 | AM | [70] | ||||||||
| Broussochalcone A | IC50 = 5.3 ± 0.3 µM | Broussonetia papyrifera | AD | [25] | ||||||
| MIC = 45 µg mL−1 | AM | [25] | ||||||||
| Broussochalcone B | IC50 = 11.1 ± 0.5 µM | AD | [25] | |||||||
| Broussonol A | A549: IC50 = 8.74 µM | Broussonetia kazinoki | AC | [45] | ||||||
| HCT-8: IC50 = 9.10 µM | AC | [45] | ||||||||
| KB: IC50 > 10 µM | AC | [45] | ||||||||
| Broussonol B | A549: IC50 = 5.52 µM | AC | [45] | |||||||
| HCT-8: IC50 = 8.80 µM | AC | [45] | ||||||||
| KB: IC50 > 10 µM | AC | [45] | ||||||||
| Broussonol C | A549: IC50 = 7.77 µM | AC | [45] | |||||||
| HCT-8: IC50 = 9.63 µM | AC | [45] | ||||||||
| KB: IC50 > 10 µM | AC | [45] | ||||||||
| Broussonol D | A549: IC50 > 10 µM | AI | [151] | |||||||
| HCT-8: IC50 > 10 µM | AI | [151] | ||||||||
| KB: IC50 = 4.15 µM | AI | [151] | ||||||||
| A549: IC50 > 10 µM | AC | [45] | ||||||||
| HCT-8: IC50 > 10 µM | AC | [45] | ||||||||
| KB: IC50 = 4.15 µM | AC | [45] | ||||||||
| Broussonol E | A549: IC50 > 10 µM | AC | [45] | |||||||
| HCT-8: IC50 > 10 µM | AC | [45] | ||||||||
| KB: IC50 > 10 µM | AC | [45] | ||||||||
| A549: IC50 > 10 µM | EI | [151] | ||||||||
| HCT-8: IC50 > 10 µM | EI | [151] | ||||||||
| KB: IC50 > 10 µM | EI | [151] | ||||||||
| Carpelastofuran | MCF-7: IC50 = 8.7 ± 0.5 µM | Artocarpus elasticus | AC | [149] | ||||||
| TK-10: IC50 = 12.3 ± 1.9 µM UACC-62: IC50 = 8.9 ± 0.3 µM | Artocarpus elasticus | AC | [43] | |||||||
| Chalcomoracin | IC50 = 2.59 ± 0.24 µM | Morus alba | AD | [31] | ||||||
| Cochinchinone A | MIC= 32–128 μg mL−1 | Maclura pomifera | AM | [75] | ||||||
| Corylifol C | IR = >65% | Artocarpus altilis | AM | [62] | ||||||
| Cudracusisoflavone M | ND | Maclura tricuspidata | AD | [30] | ||||||
| Cudracuspixanthone A | IC50 = 1.9 ± 0.4 µM | Cudrania tricuspidata | AD | [26] | ||||||
| Cudraflavanone D | IC50 = 5.7 ± 1.5 µM | AD | [26] | |||||||
| Cudraflavanone G | Against MPP+-induced cell death: EC50 > 50 µM | AA | [141] | |||||||
| Against OGD-induced neurotoxicity: EC50 > 50 µM | AA | [141] | ||||||||
| Against 6-OHDA-induced cell death: EC50 > 50 µM | AA | [141] | ||||||||
| Cudraflavone B | MIC = 12.5–50 μg mL−1 | Artocarpus altilis | AM | [98] | ||||||
| COX-1 inhibition: IC50 = 1.5 µM | Morus alba | AI | [152] | |||||||
| COX-2 inhibition: IC50 = 2.5 | AI | [109] | ||||||||
| IC50 = 20.7 ± 2.5 µM | Morus nigra | AD | [35] | |||||||
| Cudraflavone C | MIC = 12.5–50 μg mL−1 | Artocarpus altilis | AM | [98] | ||||||
| MIC = 4 μg mL−1 | Artocarpus hirsutus | AM | [58] | |||||||
| Cudraflavone H | Against MPP+-induced cell death: EC50 > 50 µM | Cudrania tricuspidata | AA | [141] | ||||||
| Against OGD-induced neurotoxicity: EC50 > 50 µM | AA | [141] | ||||||||
| Against 6-OHDA-induced cell death: EC50 > 50 µM | AA | [141] | ||||||||
| Cudraxanthone L | IC50 = 4.6 ± 0.8 µM | AD | [26] | |||||||
| Cycloartelastoxanthone | IC50 = 18.7 ± 2.2 µM | Artocarpus elasticus | AO | [90] | ||||||
| IC50 = 26.8 ± 1.2 µM | AO | |||||||||
| Cycloartocarpin | Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | ||||||
| Cyclocommunin | MCF-7: IC50 = 21.8 ± 1.3 µM | Artocarpus elasticus | AC | [149] | ||||||
| TK-10: IC50 = 40.5 ± 1.4 µM UACC-62: IC50 = 26.6 ± 3.2 µM | AC | [149] | ||||||||
| Cyclocommunol | Tyrosinase inhibition: IC50 = 209.1 µM | Artocarpus altilis | EI | [98] | ||||||
| P388: IC50 = 0.2 µM | Artocarpus elasticus | AC | [150] | |||||||
| MIC = 8–16 μg mL−1 | Morus alba | AM | [80] | |||||||
| Cyclogeracommunin | Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | ||||||
| IC50 of 73.3 ± 19.1 µM | Artocarpus elasticus | AO | [90] | |||||||
| Cycloheterophyllin | ND | Artocarpus heterophyllis | AO | [92] | ||||||
| ND | AO | |||||||||
| Cyclomorusin | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | Morus alba | AA | [136,147,148] | ||||||
| IC50 = 38.81 ± 10.39 µM | AD | [34] | ||||||||
| Tyrosinase inhibition: IC50 > 300.0 µM | Artocarpus altilis | EI | [98] | |||||||
| HeLa: IC50 = 1.66 ± 0.27 µM | Morus alba | AC | [56] | |||||||
| MCF-7: IC50 = 7.85 ± 1.30 | AC | [56] | ||||||||
| Cyclomulberrin | HeLa: IC50 = 3.69 ± 0.86 µM | AC | [56] | |||||||
| MCF-7: IC50 = 7.19 ± 0.77 µM | AC | [56] | ||||||||
| HeLa: IC50 = 3.69 ± 0.86 µM | EI | [56] | ||||||||
| MCF-7: IC50 = 7.19 ± 0.77 µM | EI | [56] | ||||||||
| Derrone | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| MIC = 4–128 μg mL−1 | Maclura pomifera | AM | [75] | |||||||
| Dorsilurin C | α-D-glucosidase inhibition: IC50 = 11.17 µM | Dorstenia psilurus | AD | [27] | ||||||
| β-D-glucosidase inhibition: IC50 = 422.21 µM β-D-mannosidase inhibition: IC50 = 358.21 µM | AD | [27] | ||||||||
| Dorsilurin F (6,8,4′-triprenyl-5,7,3′-trihydroxyflavonol) | α-D-glucosidase inhibition: IC50 = 4.13 ± 0.12 µM | AD | [27] | |||||||
| β-D-glucosidase inhibition: IC50 = 117.33 ± 0.15 µM | AD | [27] | ||||||||
| β-D-mannosidase inhibition: IC50 = 192.09 ± 0.63 µM | AD | [27] | ||||||||
| Dorsilurin G | α-D-glucosidase inhibition: IC50 = 7.51 µM | AD | [27] | |||||||
| β-D-glucosidase inhibition: IC50 = 431.14 µM | AD | [27] | ||||||||
| β-D-mannosidase inhibition: IC50 = 231.99 µM | AD | [27] | ||||||||
| Dorsilurin H | α-D-glucosidase inhibition: IC50 = 24.01 µM | AD | [27] | |||||||
| β-D-glucosidase inhibition: IC50 = 671.03 µM | AD | [27] | ||||||||
| Dorsilurin I | α-D-glucosidase inhibition: IC50 = 21.49 µM | AD | [27] | |||||||
| β-D-glucosidase inhibition: IC50 = 431.14 µM | AD | [27] | ||||||||
| Dorsilurin J | α-D-glucosidase inhibition: IC50 = 16.91 µM | AD | [27] | |||||||
| β-D-glucosidase inhibition: IC50 = 316.55 µM | AD | [27] | ||||||||
| β-D-mannosidase inhibition: IC50 = 518.27 µM | AD | [27] | ||||||||
| Dorsilurin K (5,6-7,8-bis(2,2-imethyldihydropyrano)-3′-hydroxy-4′-prenyl-flavonol) | α-D-glucosidase inhibition: IC50 = 43.95 ± 0.46 µM | AD | [27] | |||||||
| Elastichalcone B | MBC = 156.02 | Artocarpus elasticus | AM | [57] | ||||||
| Elastichalcone C | SA: IC50 = 19.5 µM MRSA: IC50 = 9.75 µM | AM | [153] | |||||||
| Elastichalcone D | ND | AM | [153] | |||||||
| Elastichalcone E | ND | AM | [153] | |||||||
| Elastichalcone F | MRSA: MBC 174.79 µM | AM | [153] | |||||||
| Erysenegalensein E | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| Erythrinin C | IC50 = 23.5 µM | Maclura tricuspidata | AD | [30] | ||||||
| Euchrenone b10 | ND | AD | [30] | |||||||
| Ficucaricone D | Inhibition of NO production: IC50 = 2.06 µM | Ficus carica | AI | [28] | ||||||
| Ficusin A | ND | Ficus hispida | AD | [28] | ||||||
| IC50 = 84.6 ± 7.8 µM | Ficus tikoua | AD | [29] | |||||||
| Furanocyclocommunin | Tyrosinase inhibition: IC50 > 220.0 µM | Artocarpus altilis | EI | [98] | ||||||
| Gancaonin A | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| Gancaonin M | ND | Maclura tricuspidata | AD | [30] | ||||||
| MIC = 2–8 μg mL−1 | Maclura cochinchinensis | AM | [74] | |||||||
| Gancaonin Q | MIC = 0.61 µg mL−1 | Dorstenia angusticornis | AM | [70] | ||||||
| Hesperetin | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | Morus alba | AA | [136,147,148] | ||||||
| Heteroflavone C | IC50 = 6.3 nmol L–1 | Artocarpus champeden | AM | [64] | ||||||
| Icariin | ND | Morus alba | AA | [136] | ||||||
| Isobavachalcone | MIC = 0.3 µg mL−1 | Dorstenia barteri | AM | [71] | ||||||
| MIC = 1–4 μg mL−1 | AM | [72] | ||||||||
| MIC = 0.5 μg mL−1 | AM | [72] | ||||||||
| ND | Artocarpus anisophyllus | AO | [87] | |||||||
| ND | Artocarpus scortechinii | AO | [88] | |||||||
| Isocycloartobiloxanthone | Tyrosinase inhibition: IC50 = 279.5 µM | Artocarpus altilis | EI | [98] | ||||||
| Isoderrone | IC50 = 108.1 ± 10.8 µM | Ficus hispida | AD | [28] | ||||||
| Isoerysenegalensein E | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| ND | Maclura tricuspidata | AD | [30] | |||||||
| Kanzonol C | MIC = 4.9 µg mL−1 | Dorstenia barteri | AM | [71] | ||||||
| Kazinol A | IC50 = 12.0 ± 0.8 µM | Broussonetia papyrifera | AD | [25] | ||||||
| Kazinol B | MIC = 20 µg mL−1 | AM | [68] | |||||||
| Kazinol E | IC50 = 10.6 ± 1.5 µM | AD | [25] | |||||||
| Kuwanon A | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | Morus alba | AA | [136,147,148] | ||||||
| Kuwanon B | MIC = 1.6 μmol L−1 µM | AM | [79] | |||||||
| Kuwanon C | ND | Morus lhou | AA | [137] | ||||||
| MICs of 4–8 μg mL−1 | Morus alba | AM | [77,78] | |||||||
| MIC = 2 and 8 μg mL−1 | AM | [81] | ||||||||
| Reducing the secretion of TNF-α: IC50 < 10 µM | AI | [52] | ||||||||
| MCF-7: IC50 = 3.92 µM | Morus nigra | AC | [154] | |||||||
| HepG2: IC50 = 9.54 µM | AC | [154] | ||||||||
| MCF-7: IC50 = 3.92 µM | AI | [154] | ||||||||
| HepG2: IC50 = 9.54 µM | AI | [154] | ||||||||
| MCF-7: IC50 = 3.92 µM | EI | [154] | ||||||||
| HepG2: IC50 = 9.54 µM | EI | [154] | ||||||||
| Kuwanon E | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | Morus alba | AA | [136,147,148] | ||||||
| AChE, BChE, BACE1 inhibition: IC50 > 100 µM | AA | [136,147,148] | ||||||||
| MICs of 2–8 μg mL−1 | AM | [80] | ||||||||
| MIC = 2 and 8 μg mL−1 | AM | [81] | ||||||||
| MIC = 4–16 μg mL−1 | AM | [81] | ||||||||
| Reducing the secretion of TNF-α and IL-1β: IC50 < 100 µM | AI | [52] | ||||||||
| Kuwanon G | ChEs inhibition: IC50 = 20.4–37.07 μM | AA | [136] | |||||||
| MCF-7: IC50 = 34.35 µM | Morus nigra | AC | [154] | |||||||
| HepG2: IC50 = 35.79 µM | AC | [154] | ||||||||
| Kuwanon S | HeLa: IC50 = 1.64 ± 0.21 µM | Morus alba | AC | [56] | ||||||
| MCF-7: IC50 = 7.02 ± 1.66 µM | AC | [56] | ||||||||
| Kuwanon T | IC50 = 10.53 ± 1.10 µM | AD | [34] | |||||||
| MICs of 2–8 μg mL−1 | AM | [81] | ||||||||
| MIC = 2 and 8 μg mL−1 | AM | [81] | ||||||||
| Kuwanon U | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | AA | [136,147,148] | |||||||
| MIC = 4–16 μg mL−1 | AM | [79] | ||||||||
| MBC = 8–32 μg mL−1 | AM | [79] | ||||||||
| MIC = 2 and 8 μg mL−1 | AM | [81] | ||||||||
| Lupalbigenin | MIC = 1 μg mL−1 | Maclura cochinchinensis | AM | [74] | ||||||
| Lupiwighteone | MIC = 2–8 μg mL−1 | AM | [74] | |||||||
| MIC = 128 μg mL−1 | AM | [74] | ||||||||
| Macluracochinone E | MIC = 1–8 μg mL−1 | AM | [74] | |||||||
| Macluracochinone A | MIC= 2–8 μg mL−1 | AM | [74] | |||||||
| Macluracochinone B | MIC= 2–8 μg mL−1 | AM | [74] | |||||||
| Macluracochinone C | MIC= 1–8 μg mL−1 | AM | [74] | |||||||
| Macluracochinone D | MIC= 1–8 μg mL−1 | AM | [74] | |||||||
| Millewanin G | IC50 = 3.2 µM | Maclura tricuspidata | AD | [30] | ||||||
| Millexatin F | MIC = 2–8 μg mL−1 | Maclura pomifera | AM | [75] | ||||||
| Moracin C | IC50 = 4.04 ± 0.84 µM | Morus alba | AD | [31] | ||||||
| Moracin N | IC50 = 2.76 ± 0.3 µM | AD | [31] | |||||||
| Mormin | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | AA | [136,147,148] | |||||||
| Morunigrol B | IC50 = 7.7 ± 0.9 µM | Morus nigra | AD | [35] | ||||||
| Morunigrol A | IC50 = 12.5 ± 1.3 µM | AD | [35] | |||||||
| Morunigrol C | IC50 = 5.3 ± 1.8 µM | AD | [35] | |||||||
| Morusin | ND | Morus alba | AA | [136,147,148] | ||||||
| ND | Morus lhou | AA | [137] | |||||||
| IC50 = 22.1 ± 2.9 µM | Morus nigra | AD | [35] | |||||||
| MIC = 12.5–50 μg mL−1 | Artocarpus altilis | AM | [98] | |||||||
| HeLa: IC50 = 0.64 ± 0.14 | Morus alba | AC | [56] | |||||||
| MCF-7: IC50 = 7.88 ± 1.89 | AC | [56] | ||||||||
| MICs of 4–8 μg mL−1 | AM | [81] | ||||||||
| MIC = 1 μg mL−1 | AM | [82] | ||||||||
| IC50 = 1819.83 ± 144.53 µM | AO | [105] | ||||||||
| IC50 = 297.83 ± 7.27 µM | AO | |||||||||
| HeLa: IC50 = 0.64 ± 0.14 µM | AI | [56] | ||||||||
| MCF-7: IC50 = 7.88 ± 1.89 µM | AI | [56] | ||||||||
| MIC = 8–32 μg mL−1 | AM | [80] | ||||||||
| IC50 = 3.19 ± 2.10 µM | AD | [32] | ||||||||
| Morusinol | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | AA | [136,147,148] | |||||||
| MIC = 2 and 8 μg mL−1 | AM | [81] | ||||||||
| Reducing the secretion of TNF-α: IC50 < 10 µM | AI | [52] | ||||||||
| Morusyunnansin C | Tyrosinase inhibition: IC50 > 25 µM | Morus yunnanensis | EI | [130] | ||||||
| Morusyunnansin D | Tyrosinase inhibition: IC50 > 25 µM | EI | [130] | |||||||
| Morusyunnansin E | Tyrosinase inhibition: IC50 = 1.43 µM | EI | [130] | |||||||
| Morusyunnansin F | Tyrosinase inhibition: IC50 = 0.15 µM | EI | [130] | |||||||
| Mulberrofuran G | AChE inhibiton: IC50 = 2.13 ± 0.02 µM | Morus alba | AA | [136] | ||||||
| BACE1 inhibition: IC50 = 0.31 ± 0.01 | Morus alba | AA | [136] | |||||||
| Myrsininone A | ND | Ficus hispida | AD | [28] | ||||||
| Neocyclomorusin | AChE, BChE, BACE1 inhibition: IC50 > 100 µM | Morus alba | AA | [136,147,148] | ||||||
| Nigrasin H | IC50 = >50 µM | Morus nigra | AD | [36] | ||||||
| Nigrasin I | IC50 = >25 µM | Morus nigra | AD | [36] | ||||||
| Papyriflavonol A | IC50 = 2.1 ± 0.2 µM | Broussonetia papyrifera | AD | [25] | ||||||
| MIC = 10 MIC = 12.5 µg mL−1 | Broussnetia papyrifera | AM | [68] | |||||||
| Sanggenol A | ND | Morus alba | AM | [75] | ||||||
| Sanggenon C | MIC = 70.6 μmol L−1 | AM | [78,79] | |||||||
| Sanggenon D | MIC = 17.6 μmol L−1 | AM | [76] | |||||||
| Sanggenon E | Reducing the secretion of TNF-α: IC50 < 10 µM | AI | [52] | |||||||
| Sanggenon H | Reducing the secretion of TNF-α: IC50 < 10 µM | AI | [52] | |||||||
| Sanggenon J | HeLa: IC50 = 2.28 ± 0.04 µM | AC | [56] | |||||||
| MCF-7: IC50 = 4.56 ± 0.71 µM | AC | [56] | ||||||||
| Sanggenon K | HeLa: IC50 = 2.29 ± 1.64 µM | AC | [56] | |||||||
| MCF-7: IC50 = 3.51 ± 0.59 µM | AC | [56] | ||||||||
| Senegalensin | ND | Maclura tricuspidata | AD | [30] | ||||||
| Soroceal | Reducing the secretion of TNF-α: IC50 < 10 µM | Morus alba | AI | [52] | ||||||
| Stipulin | MIC = 78.12 µg mL−1 | Dorstenia angusticornis | AM | [70] | ||||||
| Tomentodiplacone L | Reducing the secretion of TNF-α: IC50 = 6.7 µM | Morus alba | AI | [52] | ||||||
| Warangalone (scandenone) | Inhibition of β-hexosaminidase release: IC50 > 40 µM | Cudrania tricuspidata | AI | [62] | ||||||
| MIC = 1–8 μg mL−1 | Maclura cochinchinensis | AM | [74] | |||||||
| MIC = 4 μg mL−1 | Maclura pomifera | AM | [75] | |||||||
| Xanthone V1 | MIC = 32–128 μg mL−1 | Maclura pomifera | AM | [75] | ||||||
| Xanthone-like compound | IC50 = 42.2 ± 2.8 | Artocarpus elasticus | AO | [86] | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morante-Carriel, J.; Živković, S.; Nájera, H.; Sellés-Marchart, S.; Martínez-Márquez, A.; Martínez-Esteso, M.J.; Obrebska, A.; Samper-Herrero, A.; Bru-Martínez, R. Prenylated Flavonoids of the Moraceae Family: A Comprehensive Review of Their Biological Activities. Plants 2024, 13, 1211. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091211
Morante-Carriel J, Živković S, Nájera H, Sellés-Marchart S, Martínez-Márquez A, Martínez-Esteso MJ, Obrebska A, Samper-Herrero A, Bru-Martínez R. Prenylated Flavonoids of the Moraceae Family: A Comprehensive Review of Their Biological Activities. Plants. 2024; 13(9):1211. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091211
Chicago/Turabian StyleMorante-Carriel, Jaime, Suzana Živković, Hugo Nájera, Susana Sellés-Marchart, Ascensión Martínez-Márquez, María José Martínez-Esteso, Anna Obrebska, Antonio Samper-Herrero, and Roque Bru-Martínez. 2024. "Prenylated Flavonoids of the Moraceae Family: A Comprehensive Review of Their Biological Activities" Plants 13, no. 9: 1211. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091211