
Functional Traits of Male and Female Leaves of Hippophae tibetana on the Eastern Edge of the Tibetan Plateau and Their Altitudinal Variability
Abstract
:1. Introduction
2. Results
2.1. Physical and Chemical Properties of Soil
2.2. Functional Traits of Leaves of Hippophae tibetana
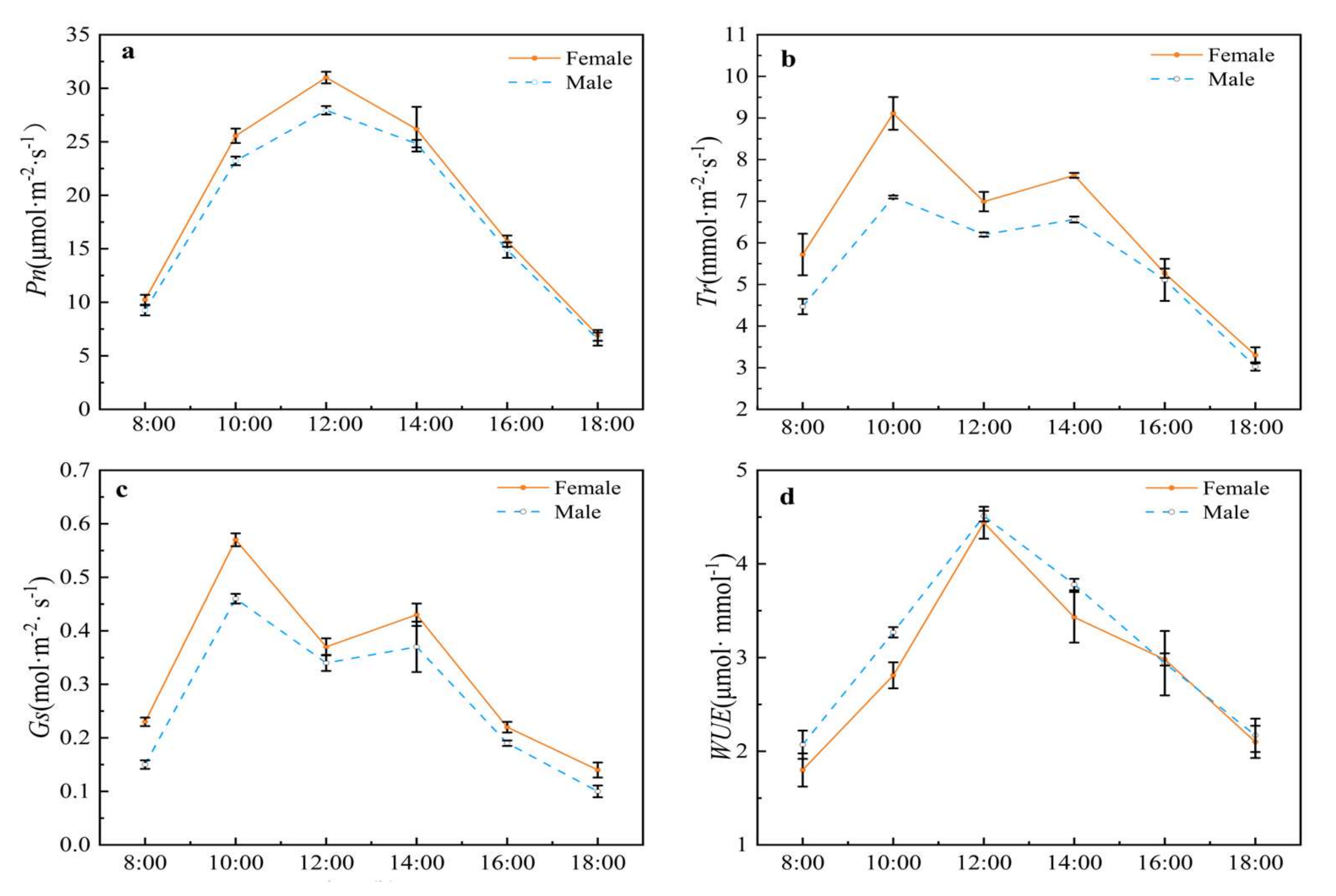
2.2.1. Photosynthetic Physiological Characteristics of Male and Female Plants of Hippophae tibetana
2.2.2. Photosynthesis-Light Response Curves of Male and Female Plants of H. tibetana
2.3. Altitudinal Changes in Physical Leaf Parameters of Male and Female H. tibetana Plants
2.3.1. Functional Traits of Male and Female Leaves of H. tibetana at Different Altitudes
2.3.2. Photosynthetic Physiological Differences between Male and Female Plants of H. tibetana at Different Altitudes
2.3.3. Correlation Analysis of Functional Traits of H. tibetana Leaves at Different Altitudes
2.4. Redundancy Analysis between Leaf Functional Traits and Environmental Factors of H. tibetana
3. Discussion
3.1. Variation of Soil Physicochemical Factors among Altitudes
3.2. Photosynthetic Physiological Characteristics of Male and Female of H. tibetana
3.3. Adaptation Strategies of Leaf Functional Traits to Altitude in Male and Female of H. tibetana
3.4. Trade-Offs in Functional Traits of Male and Female Leaves of H. tibetana at Different Altitudes and Their Correlation with Environmental Factors
4. Materials and Methods
4.1. Study Area
4.2. Sample Plot Setting and Sampling
4.3. Determination of Leaf Functional Traits
4.4. Photosynthetic Physiological Measurements
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Duan, B.; Wang, M.; Korpelainen, H.; Li, C.; Watling, J. Intra- and inter-sexual competition of Populus cathayanaunder different watering regimes. Funct. Ecol. 2014, 28, 124–136. [Google Scholar] [CrossRef]
- Delph, L.F.; Bell, D.L. A test of the differential-plasticity hypothesis for variation in the degree of sexual dimorphism in Silene latifolia. Evol. Ecol. Res. 2008, 10, 61–75. [Google Scholar]
- Li, S.; Wang, H.; Gou, W.; White, J.F.; Kingsley, K.L.; Wu, G.; Su, P. Leaf functional traits of dominant desert plants in the Hexi Corridor, Northwestern China: Trade-off relationships and adversity strategies. Glob. Ecol. Conserv. 2021, 28, e01666. [Google Scholar] [CrossRef]
- Wang, M.; Wan, P.; Guo, J.; Xu, J.; Chai, Y.; Yue, M. Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China. Isr. J. Ecol. Evol. 2017, 63, 25–32. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Xiong, K.; Yu, Y.; Min, X. Changes of leaf functional traits in karst rocky desertification ecological environment and the driving factors. Glob. Ecol. Conserv. 2020, 24, e01381. [Google Scholar] [CrossRef]
- Graae, B.J.; De Frenne, P.; Kolb, A.; Brunet, J.; Chabrerie, O.; Verheyen, K.; Pepin, N.; Heinken, T.; Zobel, M.; Shevtsova, A.; et al. On the use of weather data in ecological studies along altitudinal and latitudinal gradients. Oikos 2012, 121, 3–19. [Google Scholar] [CrossRef]
- Qiang, W.-Y.; Wang, X.-L.; Chen, T.; Feng, H.-Y.; An, L.-Z.; He, Y.-Q.; Wang, G. Variations of stomatal density and carbon isotope values of Picea crassifolia at different altitudes in the Qilian Mountains. Trees 2003, 17, 258–262. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef]
- Reich, P.B.; Uhl, C.; Walters, M.B.; Ellsworth, D.S. Leaf lifespan as a determinant of leaf structure and function among 23 amazonian tree species. Oecologia 1991, 86, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Lee, W.G. Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand. Oecologia 2003, 134, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Bartish, I.V.; Jeppsson, N.; Nybom, H.; Swenson, U. Phylogeny of Hippophae (Elaeagnaceae) inferred from parsimony analysis of chloroplast DNA and morphology. Syst. Bot. 2002, 27, 41–54. [Google Scholar]
- Sun, K.; Chen, X.-L.; Ma, R.-J.; Li, C.; Wang, Q.; Ge, S. Molecular phylogenetics of Hippophae L. (Elaeagnaceae) based on the internal transcribed spacer (ITS) sequences of nrDNA. Plant Syst. Evol. 2002, 235, 121–134. [Google Scholar] [CrossRef]
- Wang, H.; Laqiong, L.; Sun, K.; Lu, F.; Wang, Y.; Song, Z.; Wu, Q.; Chen, J.; Zhang, W. Phylogeographic structure of Hippophae tibetana (Elaeagnaceae) highlights the highest microrefugia and the rapid uplift of the Qinghai-Tibetan Plateau. Mol. Ecol. 2010, 19, 2964–2979. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liu, Q.; Zhang, Z.; Zhang, Z.; Zhou, Z.; Jiang, Y.; Huang, X. Effects of photosynthetic models on the calculation results of photosynthetic response parameters in young Larix principis-rupprechtii Mayr. plantation. PLoS ONE 2021, 16, e0261683. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Liu, X.G.; Hao, K.; Yang, Q.L.; Yang, X.Q.; Zhang, W.H.; Cong, Y. Light-response curve of photosynthesis and model fitting in leavesof Mangifera indica under different soil water conditions. Photosynthetica 2019, 57, 796–803. [Google Scholar] [CrossRef]
- Naveed, S.; Khan, I.; Sajid, M.; Mahmood, T.; Hussain, I.; Ilyas, M.; Ali, I.; Ullah, S.; Kamal, A.; Altawaha, A. Relationship of soil physico chemical properties with elevation and geographical directions. In Proceedings of the IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; p. 012172. [Google Scholar]
- Kang, E.; Li, Y.; Zhang, X.; Yan, Z.; Wu, H.; Li, M.; Yan, L.; Zhang, K.; Wang, J.; Kang, X. Soil pH and nutrients shape the vertical distribution of microbial communities in an alpine wetland. Sci. Total Environ. 2021, 774, 145780. [Google Scholar] [CrossRef]
- Ma, H.-P.; Yang, X.-L.; Guo, Q.-Q.; Zhang, X.-J.; Zhou, C.-N. Soil organic carbon pool along different altitudinal level in the Sygera Mountains, Tibetan Plateau. J. Mt. Sci. 2016, 13, 476–483. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Urbanski, L.; Hobley, E.; Lang, B.; von Lützow, M.; Marin-Spiotta, E.; van Wesemael, B.; Rabot, E.; Ließ, M.; Garcia-Franco, N. Soil organic carbon storage as a key function of soils-A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Che, M.; Gong, Y.; Xu, M.; Kang, C.; Lv, C.; He, S.; Zheng, J. Effects of elevation and slope aspect on the distribution of the soil organic carbon associated with Al and Fe mineral phases in alpine shrub-meadow soil. Sci. Total Environ. 2021, 753, 141933. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Ulhassan, Z.; Brestic, M.; Zivcak, M.; Zhou, W.; Allakhverdiev, S.I.; Yang, X.; Safdar, M.E.; Yang, W.; Liu, W. Photosynthesis research under climate change. Photosynth. Res. 2021, 150, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Hultine, K.R.; Grady, K.C.; Wood, T.E.; Shuster, S.M.; Stella, J.C.; Whitham, T.G. Climate change perils for dioecious plant species. Nat. Plants 2016, 2, 16109. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Wu, G.; Liu, X.; Chen, T.; Xu, G.; Wang, W.; Zeng, X.; Zhang, X. Elevation-dependent variations of tree growth and intrinsic water-use efficiency in Schrenk spruce (Picea schrenkiana) in the western Tianshan Mountains, China. Front. Plant Sci. 2015, 6, 309. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Yamori, W.; Kusumi, K.; Iba, K.; Terashima, I. Increased stomatal conductance induces rapid changes to photosynthetic rate in response to naturally fluctuating light conditions in rice. Plant Cell Environ. 2020, 43, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.M. Temperature and determine of plant. Plants 1998, 30–31. [Google Scholar]
- Long, S.; Humphries, S.; Falkowski, P.G. Photoinhibition of photosynthesis in nature. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 633–662. [Google Scholar] [CrossRef]
- Tao, W.; Ju, T.; Milne, R.; Miehe, G.; Wang, W.; Li, J.; Chen, L.; Mao, K. Improved photosynthesis by leaf nutrient content enhances water use efficiency in Juniperus tibetica along elevation gradients above 4000 m a.s.l. Plant Ecol. Divers. 2021, 14, 81–92. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Zhou, Z.-K.; Hu, H.; Xu, K.; Yan, N.; Li, S.-Y. Photosynthetic performances of Quercus pannosa vary with altitude in the Hengduan Mountains, southwest China. For. Ecol. Manag. 2005, 212, 291–301. [Google Scholar] [CrossRef]
- Vats, S.; Kumar, S. Photosynthetic response of Podophyllum hexandrum Royle from different altitudes in Himalayan ranges. Photosynthetica 2006, 44, 136–139. [Google Scholar] [CrossRef]
- Wu, J.; Shi, Z.; Liu, S.; Centritto, M.; Cao, X.; Zhang, M.; Zhao, G. Photosynthetic capacity of male and female Hippophae rhamnoides plants along an elevation gradient in eastern Qinghai-Tibetan Plateau, China. Tree Physiol. 2021, 41, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, G.; Zang, R.; Korpelainen, H.; Berninger, F. Sex-related differences in leaf morphological and physiological responses in Hippophae rhamnoides along an altitudinal gradient. Tree Physiol. 2007, 27, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Baldos, A.P.; Corre, M.D.; Veldkamp, E. Response of N cycling to nutrient inputs in forest soils across a 1000–3000 m elevation gradient in the Ecuadorian Andes. Ecology 2015, 96, 749–761. [Google Scholar] [CrossRef]
- Hölscher, D.; Schmitt, S.; Kupfer, K. Growth and Leaf Traits of Four Broad-Leaved Tree Species along a Hillside Gradient: Wachstum und Blatteigenschaften von vier Laubbaumarten entlang eines Hanggradienten. Forstwiss. Cent. 2002, 121, 229–239. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Bongers, F.; Poorter, L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Yan, Z.; Niklas, K.J.; Han, W.; Kattge, J.; Reich, P.B.; Luo, Y.; Chen, Y.; Tang, Z.; Hu, H. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent. Natl. Sci. Rev. 2018, 5, 728–739. [Google Scholar] [CrossRef]
- Kogami, H.; Hanba, Y.; Kibe, T.; Terashima, I.; Masuzawa, T. CO2 transfer conductance, leaf structure and carbon isotope composition of Polygonum cuspidatum leaves from low and high altitudes. Plant Cell Environ. 2001, 24, 529–538. [Google Scholar] [CrossRef]
- Hikosaka, K.; Nagamatsu, D.; Ishii, H.S.; Hirose, T. Photosynthesis–nitrogen relationships in species at different altitudes on Mount Kinabalu, Malaysia. Ecol. Res. 2002, 17, 305–313. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Keenan, T.F.; Niinemets, Ü. Global leaf trait estimates biased due to plasticity in the shade. Nat. Plants 2016, 3, 16201. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Xu, L.; Fu, Q.; Wang, Y. Effects of region and elevation on adaptation of leaf functional traits of an invasive plant Erigeron annuus in China. Phyton 2022, 91, 115. [Google Scholar] [CrossRef]
- Chen, L.; Gong, J.; Fu, B.; Huang, Z.; Huang, Y.; Gui, L. Effect of land use conversion on soil organic carbon sequestration in the loess hilly area, loess plateau of China. Ecol. Res. 2007, 22, 641–648. [Google Scholar] [CrossRef]
- Bremner, J.; Mulvaney, C. Nitrogen-Total. In Methods of Soil Analysis: Part 2. Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Wang, Z.; Sun, Y.; Chi, X.; Hong, S. Effects of Grazing Intensity on Soil Bacterial Community Structure of Stipa grandis Grasslands in Inner Mongolia, China. In Proceedings of the Fifth International Conference on Biological Information and Biomedical Engineering; Association for Computing Machinery: New York, NY, USA, 2021; pp. 1–5. [Google Scholar]
- Artur, A.; Mendonça, E.; Nóbrega, G.; Ferreira, T.; O Leão, R.; Teixeira, A.; Otero, X. Evaluation of methods for quantifying organic carbon in mangrove soils from semi-arid region. J. Soils Sediments Prot. Risk Assess. Remediat. 2015, 15, 282–291. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Low Altitude | Middle Altitude | High Altitude |
|---|---|---|---|
| SWC (%) | 21.52 ± 0.30 b | 14.98 ± 0.24 c | 27.07 ± 0.20 a |
| pH | 7.59 ± 0.04 a | 7.55 ± 0.03 a | 6.56 ± 0.05 b |
| SBD (g·cm−3) | 0.78 ± 0.003 a | 0.79 ± 0.004 a | 0.78 ± 0.004 a |
| SOC (g·kg−1) | 42.61 ± 0.46 b | 33.67 ± 0.54 c | 54.37 ± 0.90 a |
| STN (g·kg−1) | 4.59 ± 0.020 b | 3.65 ± 0.076 c | 6.18 ± 0.020 a |
| STP (g·kg−1) | 0.45 ± 0.003 c | 0.65 ± 0.004 a | 0.55 ± 0.012 b |
| Time (h) | PAR (μmol·m−2·s−1) | T leaf (°C) | ||
|---|---|---|---|---|
| Female | Male | Female | Male | |
| 8:00 | 255.75 ± 1.23 b | 299.13 ± 38.89 a | 9.67 ± 0.01 b | 10.23 ± 0.12 a |
| 10:00 | 1495.41 ± 26.49 a | 1471.35 ± 10.18 b | 13.2 ± 0.05 b | 14.18 ± 0.10 a |
| 12:00 | 1797.26 ± 162.03 a | 1704.48 ± 52.98 a | 24.26 ± 0.07 a | 23.51 ± 0.53 b |
| 14:00 | 1753.87 ± 493.28 a | 1938.45 ± 40.94 a | 27.86 ± 0.01 a | 26.35 ± 0.06 b |
| 16:00 | 1583.02 ± 56.67 a | 1472.77 ± 21.17 b | 22.39 ± 0.01 b | 24.8 ± 0.04 a |
| 18:00 | 216.33 ± 29.16 a | 161.24 ± 4.55 b | 18.79 ± 0.04 a | 17.66 ± 0.08 b |
| LSP | LCP | AQE | Rd | Amax | |
|---|---|---|---|---|---|
| Female | 1857.6 ± 61.28 a | 57.6 ± 1.23 a | 0.051 ± 0.002 a | −2.96 ± 0.150 a | 31.16 ± 0.25 a |
| Male | 1596.8 ± 44.60 b | 43.2 ± 1.47 b | 0.046 ± 0.001 b | −1.97 ± 0.006 b | 28.00 ± 3.47 b |
| Axis | Eigenvalues | Leaf–Environment Correlation Coefficient | Leaf Cumulative Percentage Variance (%) | Leaf–Environment Cumulative Percentage Variance (%) | Sum of All Eigenvalues | Sum of All Canonical Eigenvalues |
|---|---|---|---|---|---|---|
| Axis1 | 0.5725 | 0.9427 | 57.25 | 72.84 | 1.0000 | 0.7860 |
| Axis2 | 0.2070 | 0.9203 | 77.95 | 99.18 | ||
| Axis3 | 0.0031 | 0.3068 | 78.27 | 99.58 | ||
| Axis4 | 0.0020 | 0.4124 | 78.46 | 99.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, B.; Ma, Z.; Gao, P.; Lu, J.; Ding, N.; Sun, K. Functional Traits of Male and Female Leaves of Hippophae tibetana on the Eastern Edge of the Tibetan Plateau and Their Altitudinal Variability. Plants 2022, 11, 2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192484
Fan B, Ma Z, Gao P, Lu J, Ding N, Sun K. Functional Traits of Male and Female Leaves of Hippophae tibetana on the Eastern Edge of the Tibetan Plateau and Their Altitudinal Variability. Plants. 2022; 11(19):2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192484
Chicago/Turabian StyleFan, Baoli, Zongqi Ma, Pengfei Gao, Jing Lu, Nana Ding, and Kun Sun. 2022. "Functional Traits of Male and Female Leaves of Hippophae tibetana on the Eastern Edge of the Tibetan Plateau and Their Altitudinal Variability" Plants 11, no. 19: 2484. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192484