
Toward the Exploitation of Sustainable Green Factory: Biotechnology Use of Nannochloropsis spp.

Department of Biotechnology, University of Verona, 37134 Verona, Italy
*
Author to whom correspondence should be addressed.
Biology 2024, 13(5), 292; https://0-doi-org.brum.beds.ac.uk/10.3390/biology13050292
Submission received: 24 March 2024
/
Revised: 19 April 2024
/
Accepted: 24 April 2024
/
Published: 25 April 2024
(This article belongs to the Special Issue Metabolic Engineering of Microalgae for Sustainable Bioproduction: Successes, Limits, and Future Perspectives)
Abstract
:Simple Summary
With the increase in the global population, food and energy production systems need to find immediately effective alternatives. There has been increased interest in microalgae thanks to their ability to grow exploiting photosynthesis with only water and simple nutrients, even derived from wastewater. Among microalgae, Nannochloropsis genus is one of the most interesting ones due to the capacity to accumulate “good” fatty acids, omega-3 and omega-6. Currently, the cultivation of microalgae is not often cost-effective, and strain domestication must be performed. Microalgae can be manipulated through metabolic engineering to increase their biomass productivity or to obtain high-value products that overcome the cultivation cost. Here, we review the main genetic manipulation tools and present some examples to explain the potential of Nannochloropsis spp. as a green cell bio-factory. The use of genetic engineering will finally bridge the gap between the probable and feasible use of microalgae.
Abstract
Securing food, energy, and raw materials for a growing population is one of the most significant challenges of our century. Algae play a central role as an alternative to plants. Wastewater and flue gas can secure nutrients and CO2 for carbon fixation. Unfortunately, algae domestication is necessary to enhance biomass production and reduce cultivation costs. Nannochloropsis spp. have increased in popularity among microalgae due to their ability to accumulate high amounts of lipids, including PUFAs. Recently, the interest in the use of Nannochloropsis spp. as a green bio-factory for producing high-value products increased proportionally to the advances of synthetic biology and genetic tools in these species. In this review, we summarized the state of the art of current nuclear genetic manipulation techniques and a few examples of their application. The industrial use of Nannochloropsis spp. has not been feasible yet, but genetic tools can finally lead to exploiting this full-of-potential microalga.
1. Introduction
Microalgae are photosynthetic unicellular microorganisms able to assimilate CO2 through photosynthesis. Among all the organisms able to fix carbon dioxide, they have the significant advantage of being sustainable and not in competition with traditional agriculture. Their cultivation can occur in closed artificial systems, even using wastewater and industrial gasses, without the use of arable lands. Algal adaptation capacity and the possibility of growing them in fully controlled closed systems allows microalgae cultivation in geographic areas where traditional agriculture is ineffective [1,2] including planetary human habitation [3]. Microalgae are well known to be a low-cost and highly nutritive source that is useful for food or feed. Interesting nutritional profiles characterize most microalgae due to their high protein content as well as the accumulation of essential fatty acids (Ω-3 and Ω-6), vitamins, pigments, and antioxidants [4].
Microalgal biotechnology received significantly increased attention in recent years for the possibility of using these photosynthetic organisms as biorefinery for high-value products [5]. Microalgal metabolic engineering and their genetic manipulation are not a choice, and they are mandatory to make their exploitation feasible. Expensive cultivation costs do not allow the establishment of an effective industrial application for these microorganisms. In this scenario, microalgal genetic manipulation can pivot by generating mutants with suitable characteristics such as higher growth rate, biomass production, or metabolite accumulation. Moreover, using microalgae as a bio-factory for high-value products can offer a green sustainable way to produce valuable molecules and reduce cultivation costs.
Nannochloropsis spp. is unique among all microalgae, with peculiar attributes including a rapid growth rate in diverse environmental conditions and a high lipid content. The Nannochloropsis species belong to the Eustigmatophyceae (Figure 1), and their cells are spherical, non-flagellate, with a diameter of 2-8 µm [6]. Four membranes surround their chloroplast due to the secondary endosymbiosis event (Figure 2) [7]. Interestingly, the Nannochloropsis plastid is characterized by a continuum between the external chloroplast membrane and the nucleus envelope [8]. The single chloroplast occupies most of the cell volume. It contains a peculiar pigment content: only chlorophyll a (Chl a) is present, with a lack of other accessory chlorophylls, while violaxanthin, β-carotene, and vaucheriaxanthin are the most represented carotenoids under control conditions (100 μmol photons m−2s−1, 22 °C, 5% CO2 enriched air) [9].
The Nannochloropsis genus was historically composed of six known species: Nannochloropsis gaditana [10], Nannochloropsis granulata [11], Nannochloropsis oceanica [12], Nannochloropsis limnetica [13], Nannochloropsis salina [14], and Nannochloropsis oculata [14]. More recently, the results of phylogenetic analyses of the Nannochloropsis taxa on 18S rDNA revealed the presence of an additional Nannochloropsis species, N. australis [15]. In addition, the analysis on the plastid rbcL gene allowed researchers to revisit the phylogenesis of Nannochloropsis taxa, introducing a new sister genus called Microchloropsis and shifting N. gaditana and N. salina into that [15]. With this new classification, the Nannochloropsis genus comprises five species (N. oculata, N. granulata, N. oceanica, N. limnetica, and N. australis) while the Microchloropsis two (M. gaditana and M. salina). Moreover, M. gaditana is suggested to be classified as a strain of M. salina, sharing up to 98% of the chloroplastic genome and 97% of the mitochondrial one [16]. For coherency with the literature in this review, M. gaditana and M. salina will be referred to as Nannochloropsis. Except for N. limnetica, all Nannochloropsis species are found in marine environments [17].
Nannochloropsis can be cultivated on an industrial scale thanks to its robust growth and acquired popularity for its ability to accumulate high amounts of lipids and PUFAs such as arachidonic acid (AA, 20:4 n-6) and eicosapentaenoic acid (EPA, 20:5 n-3) [18,19,20,21]. Nannochloropsis can reach up to 69% of lipids on dry weight (DW) [22], 90% of TAG on total glycerolipids, and 42% of EPA on total fatty acids [23]. Its lipids include mainly the following glycerolipids: TAG, glycolipids monogalactosyldiacylglycerol (MGDG), digalactosyl-diacylglycerol (DGDG), and sulfoquinovosyl-diacylglycerol (SQDG); phospholipids: phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylglycerol (PG), and phosphatidylinositol (PI); and non-phosphorus betaine lipid diacylglycerol-N, N, N-trimethylhomoserine (DGTS).
Oleaginous algae, including Nannochloropsis, produce only small quantities of TAG under optimal growth conditions [22]. Different stressing agents, such as chemical (nutrient starvation, salinity, and pH) or physical (light intensity and temperature) result in large amounts of TAG (up to 70% DW), accompanied by considerable alterations in lipid and fatty acid profiles [18,22,24,25,26,27]. The lipid accumulation increases in response to stressing growth conditions is a widespread trait among Nannochloropsis spp., but the amplitude of this phenomenon for each stimulus is strain-specific.
Light is essential to allow photosynthesis, but an excess of irradiation can irreversibly damage the photosynthetic apparatus, leading, in the worst case, to cell death [28]. Nannochloropsis grown in high-light saturating conditions shows a higher lipid, fatty acid, and carbohydrate content [18]. The increased lipid content is accompanied by a changed profile: eicosapentaenoic acid (C20:5) and arachidonic acid (C20:4) are reduced, while palmitic acid (C16:0) and palmitoleic acid (C16:1) are increased [18]. The changes occur as a consequence of the damage/resynthesis of the thylakoid membrane where EPA is mainly located. Moreover, besides intensity, light quality can modify the fatty acid accumulation and profile. The use of monochromatic red or blue light results in 1.2- and 1.4-fold higher percentages of saturated fatty acid accumulation than using white light in N. gaditana [29].
Temperature is another critical stimulus that can modify the fatty acid profile. Microalgae modulate membrane fluidity in response to temperature changes. In N. salina, it is reported that higher temperatures favor saturated fatty acid accumulation, while when the temperature decreases, the saturation level decreases as a consequence [30].
Lastly, nitrogen starvation is considered the stimulus with the highest response in triggering lipid accumulation [22]. In particular, TAG can be increased from around 0.5% to >30% of DW, and polar lipids can decrease from 5% to 1% of DW in N. oceanica [31,32,33]. EPA is mainly present in membrane lipids and, thus, decreases under nutrient depletion in response to membrane remodeling. The modulation of fatty acid composition and TAG, in response to nitrogen starvation, even if highly conserved, depends on Nannochloropsis species [31,33,34,35]. Nannochloropsis is mainly used as fish feed or a human food additive [10,36,37], and research on its exploitation for biofuel production is still ongoing. The aquaculture use of Nannochloropsis is also due to their ability to accumulate up to 20% DW of beta-glucans (βGs), which can stimulate the immune system of farmed fish [38].
N. gaditana, N. salina, N. oceanica, N. granulata, and N. oculata genomes were fully sequenced and can be used by the scientific community [39,40,41,42]. Nowadays, the genetic manipulation of Nannochloropsis is well established thanks to the optimization of several biotechnological tools, including CRISPR-Cas [43,44,45,46,47,48], or the possibility of performing homologous recombination in the safe harbor site [49] and to the availability of tens of genetic elements, ensuring an easy and fast generation of high-level expressing strains.
Here, we review the main biotechnological goals achieved in Nannochloropsis, focusing on technologies, limits, and potentials.
2. Biotechnological Tools
2.1. Genome Availability
As previously mentioned, the genomes of several Nannochloropsis species were fully sequenced and annotated [39,40,41,42,50,51]. These nuclear genomes have an approximative size of ~29 Mb, a reduced intron content, short intergenic regions, and over 10.000 genes [39,51]. Moreover, the chloroplast and the mitochondria genomes are ~117.5 Kb and ~38 Kb, respectively [42]. Genome assembly is well supported by transcriptome data under different conditions but mainly focuses on nutrient starvation, considering the increased lipid accumulation in this condition [35,52,53,54,55,56].
2.2. Transformation Methods
Since 2008, when the first successful attempt to transform the nuclear genome of Nannochloropsis oculala was reported [57], the scientific community has efficiently established genetic manipulation tools, including several transformation methods. However, the first report on an efficient transformation by means of electroporation on Nannochloropsis whole cells was published in 2011 [58]. Electroporation-mediated methods are well established and broadly used nowadays, allowing the breaking of the rigid cell wall and efficiently introducing exogenous genetic material [59]. Other nuclear transformation methods, such as agrobacterium-mediated transformation [60] or particle bombardment [61,62], have been proposed, but their use remains limited and poorly reported.
2.3. Promoters
Efficient transgene expression relies on regulatory elements such as promoters, enhancers, and terminators. The number of promoters available for Nannochloropsis is far lower with respect to other model microalgae or plants. However, several promoting elements, including constitutive, inducible, and bidirectional, have been identified and can be used by genetic engineering community (Table 1). For transgene expression, strong endogenous promoters are usually used. Some examples of broadly used DNA regions to initiate transcription are the sequence from the gene encoding β-tubulin [63,64], lipid droplet surface protein (LDSP) [39], ubiquitin extension protein (UEP) [64], violaxanthin/chlorophyll a-binding protein 1 (VCP1) [58], elongation factor (EF) [56,65], and the Heat Shock Protein (HSP) [63]. These sequences can guarantee a high-level transcription and are widely used for Nannochloropsis metabolic engineering. Two other promoters have acquired popularity for one peculiar characteristic: these are the VCP2 promoter [58,66] and the intergenic region between two ribosomal subunit genes (Ribi) [58,67], which are bidirectional [58,67]. Bidirectional promoters offer several advantages: integrating two genes that share the same promoter reduces the length of the expression cassette and halves the number of transformation events required. This increases the transformation efficiency and decreases the time needed to obtain the desired strains. The expression level is, however, not equal, with slight differences in the two opposite directions [68].
The last class of promoters available for Nannochloropsis are the inducible ones. The sequences from sulfoquinovosyl diacylglycerol synthase 2 gene (SQD2) [69] or nitrate reductase gene (NR) [70] can be used to modulate gene expression in response to phosphorus or nitrate concentration. Inducible promoters offer the possibility to accumulate products even if they are cytotoxic for the cells. The accumulation of the enzymes responsible for toxin catalysis could not happen in the absence of the inductor. The mechanisms by which SQD2 and NR promoters induce gene expression are different: the first starts the transcription in the absence of phosphorus, while the second starts in the presence of nitrate. Both promoters, as any inducible promoter, show a basal expression level that, if very low, needs to be considered [71]. An interesting application of inducible promoters was carried out by de Grahl and colleagues in Nannochloropsis oceanica [71]. They proposed the auto-induction of gene expression by switching the nitrogen source into the growth medium from ammonium to nitrate. N. oceanica utilizes preferentially ammonium rather than nitrate because the first can be directly inserted in the pathways for amino acid synthesis. Moreover, they demonstrated that the NR gene expression is suppressed in the presence of ammonium. Considering that, they developed an auto-inducible medium using a precise amount of ammonium in the presence of nitrate; once ammonium is wholly utilized, the expression of genes driven by NR can start. By calibrating the amount of ammonium, transgene expression can be modulated in time and optimized according to algal growth.
Recently, several constitutive as well as inducible promoters were tested by evaluating their ability to drive YFP expression. The results demonstrated that the promoting sequences from the EF, α-tubulin, and NR genes were able to ensure the highest reporter expression with respect to the promoters from VCP1, LDSP, and violaxanthin/chlorophyll a-binding like protein (VCP-L) [71]. It is essential to underscore that while the promoters from EF and TUB are constitutive, the one from NR is inducible and requires nitrate-containing growth media. In addition, the synergy between promoters and the initial part of their open reading frame sequences was underlined. Promoters from EF and NR showed the highest YFP expression if the sequence was extended by 42 bp [71].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of available promoters for Nannochloropsis spp.
| Promoter | Nannochloropsis Strain | References |
|---|---|---|
| Lipid droplet surface protein (LDSP) | N. oceanica CCMP1779 | [39,56,72] |
| N. oceanica IMET1 | [49] | |
| β-tubulin | N. gaditana CCMP526 | [73] |
| N. oceanica IMET1 | [46,63] | |
| N. salina CCMP1776 | [64,74,75] | |
| Ubiquitin extension protein (UEP) | N. gaditana CCMP526 | [73] |
| N. salina CCMP1776 | [64,74] | |
| Violaxanthin/chlorophyll a-binding protein 1 (VCP1) | N. oceanica IMET1 | [44,46,49,58,76] |
| Violaxanthin/chlorophyll a-binding protein 2 (VCP2) | N. oceanica W2J3B | [58] |
| N. oceanica CCMP1779 | [66] | |
| N. oceanica IMET1 | [63] | |
| Elongation factor (EF) | N. oceanica CCMP1779 | [56,65] |
| Heat Shock Protein (HSP) | N. gaditana CCMP526 | [73,77] |
| N. oceanica IMET1 | [63] | |
| Ribosomal subunits (Ribi) | N. oceanica CCMP1779 | [56] |
| Extrinsic protein in photosystem II (EPPSII) | N. gaditana CCMP526 | [77] |
| ATPase | N. gaditana CCMP526 | [77] |
| Sulfoquinovosyl diacylglycerol synthase 2 (SQD2) | N. oceanica NIES-2155 | [69] |
| Nitrate reductase (NIT) | N. gaditana CCMP526 | [70] |
2.4. Selection Markers and Reporters for Gene Expression
The co-integration of a selectable marker and the gene of interest is a typically used strategy in genetic manipulation, not only for microalgae. Conferring the ability to grow in the presence of selective agents, this strategy allows the isolation of cells that have integrated the exogenous DNA. The bleomycin resistance protein gene (Sh ble) conferring resistance to zeomycin is the most utilized selection marker in Nannochloropsis spp. Hygromycin-B phosphotransferase conferring resistance to Hygromycin, aminoglycoside 3′-phosphotransferase which allows growth on G418 medium, and nourseothricin acetyltransferase conferring resistance to Nourseothricin are other selection markers used [68]. The available selection markers for Nannochloropsis spp. are reported in Table 2.
It is essential to underscore that not all antibiotics have the same effect on all Nannochloropsis species, and the range of concentration can be highly different. Vieler and coworkers [39] compared the impact of 10 antibiotics in four Nannochloropsis species. Six (Rifampicin, Benomyl, Nystatin, Spectinomycin, Ampicillin, and Chloramphenicol) were reported not to cause negative growth effects at any tested condition. For that reason, their use as selection markers is not possible. On the contrary, zeomycin could completely abolish algal growth using the same concentration (5 μg/mL) in all tested species. The other antibiotics showed no evenly distributed dose effect among the different Nannochloropsis species. For example, Hygromycin B worked in N. oceanica; doubling the antibiotic dose allowed for selection also in N. granulata and N. gaditana, too, but even higher concentrations had no effects in N. salina.
A different use of antibiotics was recently proposed by Zhang and colleagues [78]. The antibiotic zeomycin was used in Nannochloropsis oceanica as a 48 h transformation pre-treatment at 0.2 μg/mL dose. Antibiotic exposure causes random double strain breaks in microalgal cells, leading to a 10- to 40-fold higher transgene integration [78].
Even though homologous recombination (HR) in Nannochloropsis has been reported [58], the efficiency of this type of integration is strain-specific and highly influenced by growth conditions. Most transgenes are randomly integrated during the nuclear transformation into the host genome, and the homologous recombination efficiency is poor. In addition, HR can be a valuable tool to generate knockout mutants or in the presence of specific sites in the genome to ensure a high expression level (see Section 2.4), but it is not strictly required to perform genetic engineering and requires the identification of a “neutral” site in order not to interfere with algal metabolism. The random insertion leads to differences in integration sites that greatly influence the expression via the “position effect”, with high variability among transformants. Therefore, target proteins are typically fused to fluorescent reporters to allow the identification of lines with the highest transgene expression. Creating chimeras with proteins of interest and fluorescent reporters provides a high throughput generation of expressing lines. Moreover, besides the fast identification of the highest expressing lines, fluorescent proteins allow the performance of immunodetection or purification in the absence of specific antibodies. Nevertheless, it is essential to underline that the presence of fluorescent proteins fused to the one of interest can interfere with protein folding and, in some cases, affect protein activity. To reduce this risk, linkers (such as the GSG linker) are usually placed between the two proteins. In contrast, fluorescent proteins can be easily used in basic research because they are simply foldable, stable, and fast-detectable. In that case, reporters are largely used to compare different promoters, to optimize or set transformation methods, or to evaluate target peptide activity.
Starting from the Aequorea victoria green fluorescent protein (GFP) [79], several variants, such as Yellow FP and Cyan FP, with a different absorption/emission, were developed [56]. In addition to those, researchers can use SfCherry or the DsRed variant tdTomato derived from Discosoma spp [45,61]. The most commonly used fluorescent reporters with their absorption and emission spectra are shown in Table 3.
2.5. Genetic Knockout Strategies
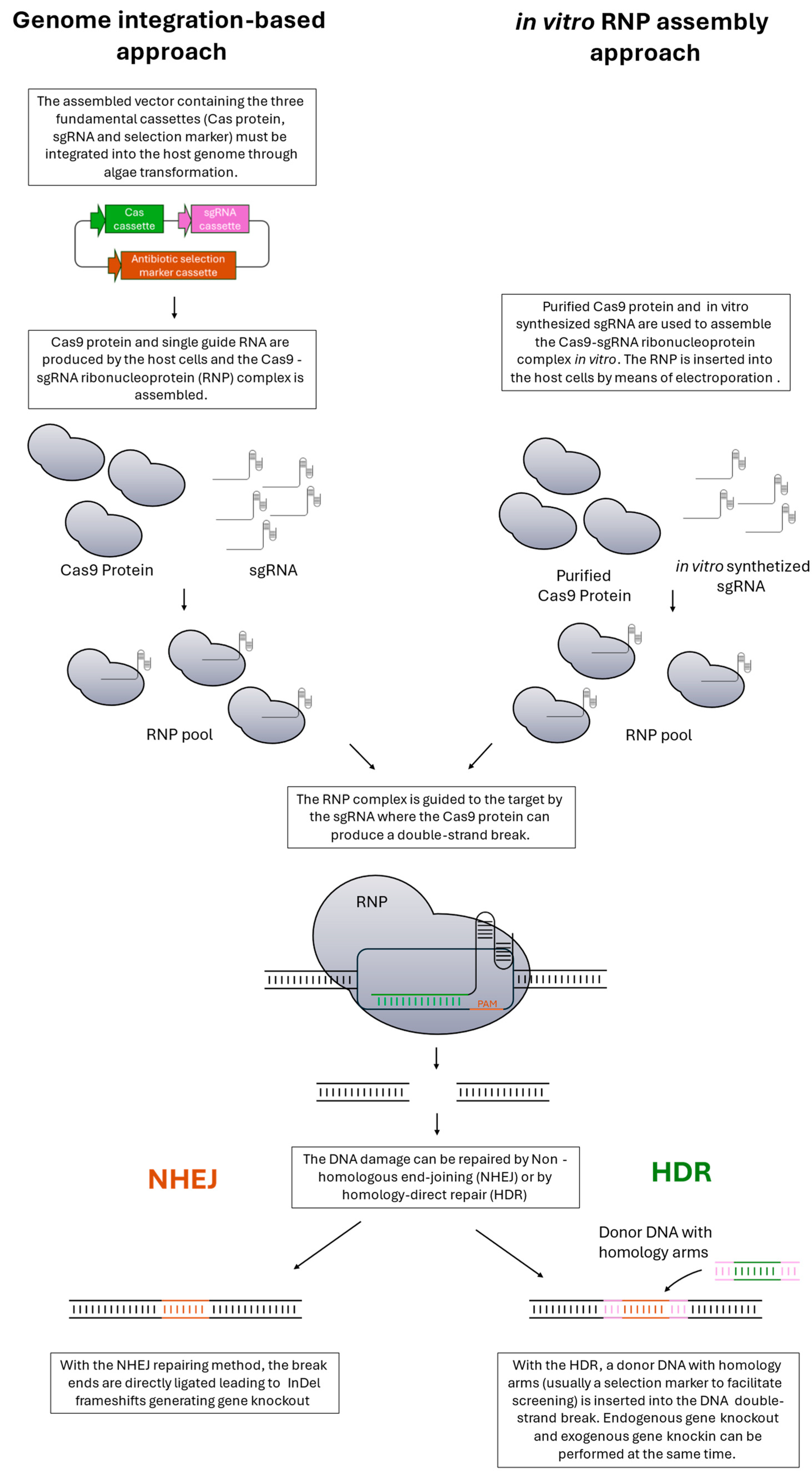
Gene downregulation or disruption is a common strategy to understand metabolic aspects or to generate backgrounds with tailored characteristics for engineering. The haploid nature of the Nannochloropsis genome facilitates the obtainment of mutants with the desired phenotype with respect to diploid or polyploid organisms. Commonly, proteins are translated starting from only one gene, causing perturbation in cell metabolism according to the importance of the enzyme when it is disrupted. Forward genetic methods (i.e., random mutagenesis or insertion) can thus be used in Nannochloropsis to obtain mutants with the desired phenotype and offer the possibility to identify key genes useful for reverse genetics. However, using random insertion or mutagenesis to generate knockout mutants has a low success rate. The insertion of the donor DNA, which must happen into an exon, is a rare event. On the other hand, the use of mutagenesis to knock out a gene presents even more constraints because the mutation can be silent or can lead to an amino acid substitution not being able to perturb the protein folding/activity. In both cases, identifying positive individuals is time-consuming because it requires the screening of many cells and only sometimes does it lead to a positive result. For that reason, targeted methods are preferred, but all reverse genetic approaches require a knowledge of genomes and pathways. Historically, the first targeted method to manipulate specific gene expression was RNA interference (RNAi). This powerful tool allows the silencing of specific genes, but their expression is not completely abolished. RNAi was largely used in Nannochloropsis to produce stains with reduced gene expression [75,82,83]. With the sequencing of genomes and the emergence of CRISPR/Cas systems, the generation of knockout mutants in Nannochloropsis became feasible and primarily applied. RNA-guided engineered nucleases (RGENs) have been largely applied to microalgae and rapidly superseded other targeted editing methods, such as zinc finger nucleases and transcription-activator-like effector nucleases. The optimization of RGEN methods, initially based on the Streptococcus pyogenes type II CRISPR/Cas9 system (SpCas9), made the generation of knockout mutants in Nannochloropsis feasible and easily accessible. The system uses a Cas9 nuclease and a single guide RNA to induce DNA breaks into the genome; during the repair of these breaks, random insertion/deletion (InDel) or exogenous DNA insertion by HR can occur, generating knockout mutants. For a more detailed description of the mechanism, see Figure 3. The first reported attempt dates to 2016, when Wang and colleagues successfully knocked out the nitrate reductase gene in N. oceanica using the CRISPR/Cas9 system [46]. Since then, several improvements in the techniques have been made to increase the knockout efficiency [43,44,45,46,47]. The ease and low cost of this technology resulted in a significant diffusion of RGENs CRISPR/Cas systems. However, the constitutively stable expression of Cas protein into the host genome presents few constraints. One of the already reported problems in this technique is the cytotoxicity of Cas once integrated into the algal genome. Previously identified and reported in the model microalga Chlamydomonas reinhardtii, this cytotoxic effect results in an extremely low targeted mutagenesis efficiency [84]. To overcome that, DNA-free RGEN-based methods are proposed for several organisms [85,86,87,88,89]. These approaches are based on in vitro preassembled Cas-gRNA ribonucleoproteins. As mentioned before, the DNA-free approach reduces the off-target and cytotoxic effects because the Cas protein is only transiently present. Furthermore, the technology does not require codon optimization or specific promoters being rapidly transferrable between different organisms. The preassembled Cas-sgRNA ribonucleoprotein-based method was also successfully used in Nannochloropsis oceanica by inserting the complex using electroporation [44]. Two other approaches to produce gene knockout without a stable Cas integration into genome must be cited. First, Poliner and coworkers developed an episomal-based system exploiting the CEN/ARS6 region from S. cerevisiae [48]. The main advantage of this technique is to represent a marker-free system; once the selective pressure is removed, the episomal is lost and no longer present in microalgae. With a similar episomal-based system exploiting the Cas12 activity, Naduthodi and coworkers recently performed multiple gene editing in N. oceanica [45]. In the same paper, the possibility of performing CRISPR-mediated interference was also presented [45]. Although Cas9 was historically the first Cas used and is currently the most used one, recently, Cas12a (formerly called Cpf1), a member of class V, has gained global attention as an interesting alternative. Cas12 presents a more minimalistic system than Cas9 since the latter consists of two nuclease domains that produce a blunt end cleavage, whereas Cas12a has a single domain that generates staggered ends on both the target and the non-target DNA strands (Naduthodi et al., 2018; Swarts et al., 2017). In addition, the protospacer adjacent motif (PAM) sequence recognized by Cas9 is placed downstream of the spacer sequence on the non-target strand; the Cas12a PAM sequence is located upstream of the spacer. For these reasons, Cas12a exhibits higher efficiencies than Cas9 in generating mutants through HDR and fewer off-target effects [45]. Lastly, it is important to cite that homologous recombination (HR), even with the abovementioned limits, is another possibility for generating knockout mutants [90,91].
2.6. Multiple Gene Expression
The possibility of overexpressing multiple genes represents one of the main desirable goals for the metabolic engineering scientific community. Metabolic engineering is the use of genetic manipulation to modify the metabolic network of cells to produce desirable products or to tune phenotype traits. Usually, manipulating a single gene does not lead to the accumulation of the desired product, and a complex rearrangement of the overall metabolic pathway needs to be performed by acting on several enzymes. The typical approach involves the use of sequential individual transformations, exponentially increasing transformation markers and fluorophores, and taking the time to generate expressing lines. Moreover, besides the limited number of markers available, the promoter pool is limited too: using the same promoting regions several times can lead to regulatory mechanisms decreasing the transcription level. Unless specific manipulations in that direction are made, the number of transcription factors able to bind a specific region is limited. Using the same promoter region to express several genes could lead to “compensative” mechanisms in which the last added gene is transcribed at the expense of the others already present. For these reasons, the insertion of multiple genes is a challenging topic. The first possibility, also exploited in other organisms, is to use the self-cleaving 2A peptide [94]. This peptide is known for generating independent proteins starting from the same transcript. This reduces the number of promoters/terminators and transformation events without generating fusion proteins. Although the 2A peptide technology has already been used in N. oceanica [80] and N. salina [95], the cleavage efficiency is less than 50%. As an alternative, Poliner and coworkers recently developed a high-capacity gene stacking toolkit to ensure the expression of several genes using a single vector, and thus one transformation event and one selection marker [68]. Using this new technology, which takes advantage of Gateway cloning and the bidirectional promoter Ribi, researchers could co-express three reporter genes (GFP, Flux, and Nlux) in addition to antibiotic resistance.
2.7. Genomic Safe Harbor as a Method of Maximizing Transgene Expression
Using microalgae as a bio-factory for producing metabolites has yet to be feasible and sustainable. This is mainly due to the poor transgene expression in microalgal hosts, primarily caused by the aforementioned “position effect”. Recently, Südfeld and coworkers identified the rDNA cistron on chromosome 3 of N. oceanica as a genomic safe harbor for transgene expression by Pol I [49]. A new expression system based on a chimeric internal ribosome entry site (IRES) and a landing pad strain allows a high-throughput generation of expressing lines. The system, summarized in Figure 4, uses a landing pad strain in which the fluorescent gene tdTomato is inserted into the safe harbor site. A gene of interest can be inserted in this site by homologous recombination, removing the fluorescent protein gene. Thus, this system has two main advantages: it ensures the highest transgene expression already reported and allows the fast and easy screening of transformants.
2.8. Chloroplast Genome Manipulation
The majority of genetically engineered organisms are generated by integrating transgenes in their nuclear genome. However, photosynthetic organisms also offer the opportunity for chloroplast transformation. Integrating transgenes into the chloroplast genome has several advantages, including the possibility of performing homologous recombination, avoiding position effects, and the absence of silencing processes in this organelle [96]. Chloroplast transformation has been reported in higher plants (see [97] for a review), but the plastid microalgal biotechnology is well established only in C. reinhardtii [98,99,100]. Only two reports showed chloroplast engineering in Nannochloropsis [101,102]. Gan and coworkers successfully integrated into the chloroplast genome of N. oceanica the GFP-encoding gene, leading to a clear GFP protein accumulation and fluorescence [101]. In addition, Cui et al. engineered the plastid genome of N. gaditana by inserting two codon-optimized antimicrobial peptide-coding genes in a polycistronic sequence. Besides the fact that, to our knowledge, these are the only successful attempts, the reported transformation efficiency is low and must be improved to feasibly exploit this technology. However, at the same time, they show that also in Nannochloropsis, the chloroplast is a suitable compartment for heterologous protein expression.
3. Biotechnological Exploitation: A Few Examples
Metabolic engineering in Nannochloropsis, albeit feasible, is rarely used with respect to other microalgae such as Chlamydomonas [103]. The potential of Nannochloropsis as a host for metabolite production is high and needs to be explored with the abovementioned genetic tools. Here are several examples of genetic manipulation approaches aimed at generating strains with suitable characteristics.
3.1. Lipids
Our current knowledge of the lipid pathways in Nannochloropsis spp. is limited, and the same applies to the evolutionary close groups of diatoms and brown algae. Nevertheless, these pathways are strongly conserved among microalgae and plants. Therefore, it is possible to take advantage of our understanding of the model organisms Chlamydomonas reinhardtii and Arabidopsis thaliana.
The de novo synthesis of fatty acids occurs mainly in the chloroplast, and, subsequently, they are exported to the ER. In contrast, the glycerolipids are assembled both in the chloroplast and ER (where the pathway is known as the Kennedy pathway). The chloroplast and ER contain similar enzymes: acyltransferases (GPAT, LPAAT), phosphatases (PAP), and synthases (CDS, PGPS). On the contrary, different acyl donors, acyl-acyl-carrier protein in the chloroplast and acyl-coA in ER, mediate the acylation step. Fatty acid modification, such as desaturation, primarily occurs in chloroplast and ER membranes, while elongation happens in the chloroplast and ER through acyl-ACP and acyl-coA, respectively. Lipid remodeling via acyltransferases and lipases is fundamental for fatty acid exchanges between membrane lipids and TAG [104]. An overview of Nannochloropsis lipid metabolism is reported in Figure 5.
To increase the oil production in Nannochloropsis spp. several metabolic engineering strategies have been used, including transcription factors, enzymes involved in lipid pathways, and enzymes involved in other pathways. The overexpression of the bZIP1 transcription factor in N. oceanica increased intracellular lipid content by 100% and enabled unprecedented lipid secretion [105]. The knockout of APETALA2-like transcription factor in N. oceanica increased neutral lipid content by 40% through the upregulation of genes involved in fatty acid biosynthesis, the CBB cycle, and sugar metabolism [76]. Moreover, the overexpression of acyl-CoA: diacylglycerol acyltransferase (DGAT1) in N. oceanica increased TAG and TFA by 50% and 14% of DW, respectively [82], while the upregulation of DGAT2 showed a slight effect on TAG, and DGAT5 increased TAG fatty acids on TFA from 10 to 40% [65]. Lastly, the overexpression of a putative endogenous gene for malonyl CoA: ACP malonyltransferase (MCMT) in N. oceanica improved lipid content by 30% of DW [106].
Lipid content was also tuned by acting on enzymes not directly involved in their biosynthesis. The overexpression of a deoxy-xylulose 5-phosphate synthase (AtDXSoe3) from A. thaliana in N. oceanica increased, under high light, total lipids and TAG on DW by 60% and 80%, respectively [107]. Moreover, mutation on a DGDG synthase in N. gaditana increased FAME content from 100 to 160 mg/g DW [26]. Lastly, the overexpression of malic enzyme (ME) in N. salina increased biomass and FAMEs on DW by 15% and 30%, respectively [108].
Recently, few efforts have been made to increase EPA content. The overexpression of Δ5 desaturase in N. oceanica increased EPA content from 17 to 22% of total lipids, and the co-expression of Δ5 and 12 desaturases enabled it to reach 25% of EPA [80]. Moreover, the co-expression of a DGAT from C. reinhardtii (CrDGTT1) and an endogenous Δ0 elongase (Δ0-ELO1) increased EPA content in TAG up to 7.3%, which was 5.9-fold higher than WT [109].
3.2. Carotenoids and Other Pigments
As mentioned, the Nannochloropsis chloroplast evolved from secondary endosymbiosis and contains only Chlorophyll a in its light-harvesting complexes. Violaxanthin, vaucheriaxanthin, and β-carotene are the most abundant carotenoids, with neoxanthin, antheraxanthin, zeaxanthin, canthaxanthin, and astaxanthin accumulated only in traces [110]. However, changing nutritional and environmental factors can vary the carotenoid and chlorophyll content.
Carotenoids and chlorophylls in Nannochloropsis are extensively studied. The fundamental role in the light harvesting, energy dissipation, and photosynthetic protein folding of carotenoids is broadly reported in all photosynthetic organisms. Their investigation is, thus, essential to disentangle molecular mechanisms and to increase the general knowledge about Nannochloropsis. On the other hand, biotechnological approaches aimed at manipulating pigment content can be performed to generate strains with tailored pigment content.
For example, the overexpression of the LHCYB (lycopene beta cyclase) gene in N. oceanica increased carotenoid content, especially β-carotene [110]. In another case, the knockout of the VDE gene generated by insertional mutagenesis allowed the reduction in the zeaxanthin content and, at the same time, demonstrated the fundamental role of this pigment for light energy dissipation and ROS protection [111]. Moreover, Dauterman and coworkers generated a VDL (violaxanthin-de-epoxidase-like) mutant in N. oceanica with an increased violaxanthin accumulation [112]. In addition, Liu and colleagues obtained the same result with a different strategy: they overexpressed in N. oceanica the ZEP enzymes responsible for the epoxidation of zeaxanthin to violaxanthin. This manipulation led the microalgae to increase violaxanthin accumulation at the expense of zeaxanthin [113]. Moreover, they also demonstrated that, on the contrary, the suppression of the ZEP gene results in a decrease in violaxanthin and an increase in zeaxanthin contents [113]. Gamma-rays were also used to increase the violaxanthin content; random mutagenesis in N. oceanica allowed for the identification of a mutant with an increased violaxanthin content and an enhanced biomass production [114]. Efforts were also made to improve the astaxanthin content of Nannochloropsis. By means of chemical mutagenesis, Cecchin and coworkers isolated a new Nannochloropsis gaditana strain characterized by increased lipid and ketocarotenoid accumulation [115].
Chlorophylls are also an interesting target for metabolic engineering. Koh and colleagues acted on chlorophyll biosynthesis to increase the productivity of microalgae biomass. Nannochloropsis spp. completely lack accessory chlorophylls, but they demonstrated that the heterologous expression of chlorophyllide a oxygenase from Chlamydomonas (CrCAO) in N. salina, catalyzing the two-step oxygenation of chlorophyllide a to chlorophyll b, leads to positive effects, increasing the growth rate [116].
3.3. Other High-Value Products and Biotechnological Manipulation
Nannochloropsis spp. are widely used in aquaculture. The biotechnological production of high-value products also reflects this application. Chen and coworkers were able to engineer N. oculata to produce fish growth hormone, which promotes the growth of Tilipia larvae [57]. N. oculata was also used for antimicrobial peptide synthesis; the bovine lactoferricin produced in this host increased the survival rate of fish after Vibrio parahaemolyticus infection [117]. Lastly, as a model vaccine for fish immunization, the viral surface protein 2 (VP2) of a pathogenic fish virus that causes infectious pancreatic necrosis was produced in N. oceanica [118].
Another important metabolite was produced by exploiting the safe harbor pipeline. A camelid-type VHH antibody was successfully synthesized in N. oceanica [49].
Another class of molecules that can be produced in microalgae and with a broad range of biotechnological applications, such as flavoring, biopharmaceuticals, and cosmetics, are terpenoids {Masyita, 2022 #710}. C. reinhardtii was broadly used as chassis for their production [119,120,121,122]. In N. oceanica, Du and colleagues produced the diterpene casbene, widely used as an intermediate in the pharmaceutical industry [123]. Despite terpenoids’ potential, no other cases are reported for them in Nannochloropsis.
The reduction in cultivation costs is strictly connected to increasing microalgal growth. Increasing biomass production is one of the main desirable goals to make microalgae use cost-effective and feasible. Perin and colleagues screened EMS-obtained N. gaditana random mutants for better photosynthetic performance. They identified mutants in which photosynthetic antennae, or the heat dissipation mechanism, are affected with higher growth in a photobioreactor [124]. In addition, Arora and coworkers identified, using the same technique, two mutants exhibiting higher carbohydrate and pigment productivity [125]. Lastly, with a completely different approach, the basic helix-loop-helix transcription factor (NsbHLH2) overexpression improved biomass and lipid production [126].
4. Conclusions
The state-of-the-art engineering strategies are summarized in this review, and the examples reported demonstrate the potential of Nannochloropsis as an emerging green bio-factory for valuable bioproducts. However, further research is necessary to enrich the number of available genetic blocks, increase production titers, and allow industrially relevant applications. The use of Nannochloropsis as an algal chassis still has a long road ahead but the potential for exploitation is high and the possibility is concrete with the latest advantages in genetic manipulation tools.
Author Contributions
F.P. and D.C. designed the structure of the review. E.C. prepared the tables and figures. All the authors contributed to searching for the relevant literature, discussed together, and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank Matteo Ballottari for his suggestions.
Conflicts of Interest
The authors declare the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Rasheed, R.; Thaher, M.; Younes, N.; Bounnit, T.; Schipper, K.; Nasrallah, G.K.; Al Jabri, H.; Gifuni, I.; Goncalves, O.; Pruvost, J. Solar cultivation of microalgae in a desert environment for the development of techno-functional feed ingredients for aquaculture in Qatar. Sci. Total Environ. 2022, 835, 155538. [Google Scholar] [CrossRef]
- Winckelmann, D.; Bleeke, F.; Thomas, B.; Elle, C.; Klöck, G. Open pond cultures of indigenous algae grown on non-arable land in an arid desert using wastewater. Int. Aquat. Res. 2015, 7, 221–233. [Google Scholar] [CrossRef]
- Revellame, E.D.; Aguda, R.; Chistoserdov, A.; Fortela, D.L.; Hernandez, R.A.; Zappi, M.E. Microalgae cultivation for space exploration: Assessing the potential for a new generation of waste to human life-support system for long duration space travel and planetary human habitation. Algal Res. 2021, 55, 102258. [Google Scholar] [CrossRef]
- Williamson, E.; Ross, I.L.; Wall, B.T.; Hankamer, B. Microalgae: Potential novel protein for sustainable human nutrition. Trends Plant Sci. 2024, 29, 370–382. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef]
- Kandilian, R.; Lee, E.; Pilon, L. Radiation and optical properties of Nannochloropsis oculata grown under different irradiances and spectra. Bioresour. Technol. 2013, 137, 63–73. [Google Scholar] [CrossRef]
- Janouskovec, J.; Horák, A.; Oborník, M.; Lukes, J.; Keeling, P.J. A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids. Proc. Natl. Acad. Sci. USA 2010, 107, 10949–10954. [Google Scholar] [CrossRef]
- Guo, L.; Liang, S.; Zhang, Z.; Liu, H.; Wang, S.; Pan, K.; Xu, J.; Ren, X.; Pei, S.; Yang, G. Genome assembly of Nannochloropsis oceanica provides evidence of host nucleus overthrow by the symbiont nucleus during speciation. Commun. Biol. 2019, 2, 249. [Google Scholar] [CrossRef]
- Basso, S.; Simionato, D.; Gerotto, C.; Segalla, A.; Giacometti, G.M.; Morosinotto, T. Characterization of the photosynthetic apparatus of the Eustigmatophycean Nannochloropsis gaditana: Evidence of convergent evolution in the supramolecular organization of photosystem I. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2014, 1837, 306–314. [Google Scholar] [CrossRef]
- Lubián, L.M. Nannochloropsis gaditana spec. nov., una nueva Eustigmatophyceae marina. Lezama 1982, 4, 287–293. [Google Scholar]
- Karlson, B.; Potter, D.; Kuylenstierna, M.; Andersen, R.A. Ultrastructure, pigment composition, and 18S rRNA gene sequence for Nannochloropsis granulata sp. nov. (Monodopsidaceae, Eustigmatophyceae), a marine ultraplankter isolated from the Skagerrak, northeast Atlantic Ocean. Phycologia 1996, 35, 253–260. [Google Scholar] [CrossRef]
- Suda, S.; Atsumi, M.; Miyashita, H. Taxonomic characterization of a marine Nannochloropsis species, N. oceanica sp. nov. (Eustigmatophyceae). Phycologia 2002, 41, 273–279. [Google Scholar] [CrossRef]
- Krienitz, L.; Hepperle, D.; Stich, H.-B.; Weiler, W. Nannochloropsis limnetica (Eustigmatophyceae), a new species of picoplankton from freshwater. Phycologia 2000, 39, 219–227. [Google Scholar] [CrossRef]
- Hibberd, D.J. Notes on the taxonomy and nomenclature of the algal classes Eustigmatophyceae and Tribophyceae (synonym Xanthophyceae). Bot. J. Linn. Soc. 2008, 82, 93–119. [Google Scholar] [CrossRef]
- Fawley, M.W.; Jameson, I.; Fawley, K.P. The phylogeny of the genus Nannochloropsis (Monodopsidaceae, Eustigmatophyceae), with descriptions of N. australis sp. nov. and Microchloropsis gen. nov. Phycologia 2015, 54, 545–552. [Google Scholar] [CrossRef]
- Starkenburg, S.R.; Kwon, K.J.; Jha, R.K.; McKay, C.; Jacobs, M.; Chertkov, O.; Twary, S.; Rocap, G.; Cattolico, R.A. A pangenomic analysis of the Nannochloropsisorganellar genomes reveals novel genetic variations in key metabolic genes. BMC Genom. 2014, 15, 212. [Google Scholar] [CrossRef]
- Jinkerson, R.E.; Radakovits, R.; Posewitz, M.C. Genomic insights from the oleaginous model alga Nannochloropsis gaditana. Bioengineered 2013, 4, 37–43. [Google Scholar] [CrossRef]
- Sukenik, A.; Carmeli, Y.; Berner, T. Regulation of Fatty Acid Composition by Irradiance Level in the Eustigmatophyte Nannochloropsis sp. J. Phycol. 1989, 25, 686–692. [Google Scholar] [CrossRef]
- Boussiba, S.; Vonshak, A.; Cohen, Z.; Avissar, Y.J.; Richmond, A.E. Lipid and biomass production by the halotolerant microalga Nannochloropsis salina. Biomass 1987, 12, 37–47. [Google Scholar] [CrossRef]
- Chini Zittelli, G.; Lavista, F.; Bastianini, A.; Rodolfi, L.; Vincenzini, M.; Tredici, M.R. Production of eicosapentaenoic acid by Nannochloropsis sp. cultures in outdoor tubular photobioreactors. J. Biotechnol. 1999, 70, 299–312. [Google Scholar] [CrossRef]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry 2002, 61, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, J.; Cui, J.; Feng, Y.; Cui, Q. Metabolic profiles of Nannochloropsis oceanica IMET1 under nitrogen-deficiency stress. Bioresour. Technol. 2013, 130, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Han, D.; Gerken, H.G.; Li, Y.; Sommerfeld, M.; Hu, Q.; Xu, J. Molecular mechanisms for photosynthetic carbon partitioning into storage neutral lipids in Nannochloropsis oceanica under nitrogen-depletion conditions. Algal Res. 2015, 7, 66–77. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.; Lukyanov, A.; Solovchenko, O.; Didi-Cohen, S.; Boussiba, S.; Khozin-Goldberg, I. Interactive effects of salinity, high light, and nitrogen starvation on fatty acid and carotenoid profiles in Nannochloropsis oceanica CCALA 804. Eur. J. Lipid Sci. Technol. 2014, 116, 635–644. [Google Scholar] [CrossRef]
- Cecchin, M.; Berteotti, S.; Paltrinieri, S.; Vigliante, I.; Iadarola, B.; Giovannone, B.; Maffei, M.E.; Delledonne, M.; Ballottari, M. Improved lipid productivity in Nannochloropsis gaditana in nitrogen-replete conditions by selection of pale green mutants. Biotechnol. Biofuels 2020, 13, 78. [Google Scholar] [CrossRef]
- Fakhry, E.M.; El Maghraby, D.M. Lipid accumulation in response to nitrogen limitation and variation of temperature in Nannochloropsis salina. Bot. Stud. 2015, 56, 6. [Google Scholar] [CrossRef]
- Barber, J.; Andersson, B. Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem. Sci. 1992, 17, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Sung, M.-G.; Nam, K.; Moon, M.; Kwon, J.-H.; Yang, J.-W. Effect of monochromatic illumination on lipid accumulation of Nannochloropsis gaditana under continuous cultivation. Bioresour. Technol. 2014, 159, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Van Wagenen, J.; Miller, T.W.; Hobbs, S.; Hook, P.; Crowe, B.; Huesemann, M. Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis salina. Energies 2012, 5, 731–740. [Google Scholar] [CrossRef]
- Li, J.; Han, D.; Wang, D.; Ning, K.; Jia, J.; Wei, L.; Jing, X.; Huang, S.; Chen, J.; Li, Y.; et al. Choreography of Transcriptomes and Lipidomes of Nannochloropsis Reveals the Mechanisms of Oil Synthesis in Microalgae. Plant Cell 2014, 26, 1645–1665. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Cao, X.; Yao, C.; Xue, S.; Yang, Q. Identification of the role of polar glycerolipids in lipid metabolism and their acyl attribution for TAG accumulation in Nannochloropsis oceanica. Algal Res. 2017, 24, 122–129. [Google Scholar] [CrossRef]
- Han, D.; Jia, J.; Li, J.; Sommerfeld, M.; Xu, J.; Hu, Q. Metabolic Remodeling of Membrane Glycerolipids in the Microalga Nannochloropsis oceanica under Nitrogen Deprivation. Front. Mar. Sci. 2017, 4, 242. [Google Scholar] [CrossRef]
- Simionato, D.; Block, M.A.; La Rocca, N.; Jouhet, J.; Maréchal, E.; Finazzi, G.; Morosinotto, T. The response of Nannochloropsis gaditana to nitrogen starvation includes de novo biosynthesis of triacylglycerols, a decrease of chloroplast galactolipids, and reorganization of the photosynthetic apparatus. Eukaryot Cell 2013, 12, 665–676. [Google Scholar] [CrossRef]
- Liang, J.; Wen, F.; Liu, J. Transcriptomic and lipidomic analysis of an EPA-containing Nannochloropsis sp. PJ12 in response to nitrogen deprivation. Sci. Rep. 2019, 9, 4540. [Google Scholar] [CrossRef]
- Lubian, L.M.; Montero, O.; Moreno-Garrido, I.; Huertas, I.E.; Sobrino, C.; Gonzalez-del Valle, M.; Pares, G. Nannochloropsis (Eustigmatophyceae) as source of commercially valuable pigments. J. Appl. Phycol. 2000, 12, 249–255. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; McKuin, B.; Fitzgerald, D.S.; Nash, H.M.; Greenwood, C. Microalgae-blend tilapia feed eliminates fishmeal and fish oil, improves growth, and is cost viable. Sci. Rep. 2020, 10, 19328. [Google Scholar] [CrossRef]
- Kumar, P.; Jain, K.; Munilkumar, S.; Chalal, R.S. Beta glucan: Avaluable nutraceutical for promoting health in aquaculture (short review). Res. Rev. BioSci. RRBS 2014, 5, 220–227. [Google Scholar]
- Vieler, A.; Wu, G.; Tsai, C.H.; Bullard, B.; Cornish, A.J.; Harvey, C.; Reca, I.B.; Thornburg, C.; Achawanantakun, R.; Buehl, C.J.; et al. Genome, functional gene annotation, and nuclear transformation of the heterokont oleaginous alga Nannochloropsis oceanica CCMP1779. PLoS Genet. 2012, 8, e1003064. [Google Scholar] [CrossRef]
- Liang, C.; Cao, S.; Zhang, X.; Zhu, B.; Su, Z.; Xu, D.; Guang, X.; Ye, N. De Novo Sequencing and Global Transcriptome Analysis of Nannochloropsis sp. (Eustigmatophyceae) Following Nitrogen Starvation. BioEnergy Res. 2013, 6, 494–505. [Google Scholar] [CrossRef]
- Ohan, J.A.; Hovde, B.T.; Zhang, X.L.; Davenport, K.W.; Chertkov, O.; Han, C.; Twary, S.N.; Starkenburg, S.R. Nuclear Genome Assembly of the Microalga Nannochloropsis salina CCMP1776. Microbiol. Resour. Announc. 2019, 8, e00750-19. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ning, K.; Li, J.; Hu, J.; Han, D.; Wang, H.; Zeng, X.; Jing, X.; Zhou, Q.; Su, X.; et al. Nannochloropsis Genomes Reveal Evolution of Microalgal Oleaginous Traits. PLOS Genet. 2014, 10, e1004094. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Jiang, Z.; Liu, B. A CRISPR/dCas9-based transcription activated system developed in marine microalga Nannochloropsis oceanica. Aquaculture 2022, 546, 737064. [Google Scholar] [CrossRef]
- Naduthodi, M.I.S.; Mohanraju, P.; Südfeld, C.; D’Adamo, S.; Barbosa, M.J.; van der Oost, J. CRISPR–Cas ribonucleoprotein mediated homology-directed repair for efficient targeted genome editing in microalgae Nannochloropsis oceanica IMET1. Biotechnol. Biofuels 2019, 12, 66. [Google Scholar] [CrossRef]
- Naduthodi, M.I.S.; Südfeld, C.; Avitzigiannis, E.K.; Trevisan, N.; van Lith, E.; Alcaide Sancho, J.; D’Adamo, S.; Barbosa, M.; van der Oost, J. Comprehensive Genome Engineering Toolbox for Microalgae Nannochloropsis oceanica Based on CRISPR-Cas Systems. ACS Synth. Biol. 2021, 10, 3369–3378. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lu, Y.; Xin, Y.; Wei, L.; Huang, S.; Xu, J. Genome editing of model oleaginous microalgae Nannochloropsis spp. by CRISPR/Cas9. Plant J. 2016, 88, 1071–1081. [Google Scholar] [CrossRef]
- Wang, Q.; Gong, Y.; He, Y.; Xin, Y.; Lv, N.; Du, X.; Li, Y.; Jeong, B.-R.; Xu, J. Genome engineering of Nannochloropsis with hundred-kilobase fragment deletions by Cas9 cleavages. Plant J. 2021, 106, 1148–1162. [Google Scholar] [CrossRef] [PubMed]
- Poliner, E.; Takeuchi, T.; Du, Z.-Y.; Benning, C.; Farré, E.M. Nontransgenic Marker-Free Gene Disruption by an Episomal CRISPR System in the Oleaginous Microalga, Nannochloropsis oceanica CCMP1779. ACS Synth. Biol. 2018, 7, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Südfeld, C.; Pozo-Rodríguez, A.; Manjavacas Díez, S.A.; Wijffels, R.H.; Barbosa, M.J.; D’Adamo, S. The nucleolus as a genomic safe harbor for strong gene expression in Nannochloropsis oceanica. Mol. Plant 2022, 15, 340–353. [Google Scholar] [CrossRef]
- Schwartz Ariel, S.; Brown, R.; Ajjawi, I.; McCarren, J.; Atilla, S.; Bauman, N.; Richardson Toby, H. Complete Genome Sequence of the Model Oleaginous Alga Nannochloropsis gaditana CCMP1894. Genome Announc. 2018, 6, e01448-17. [Google Scholar] [CrossRef]
- Corteggiani Carpinelli, E.; Telatin, A.; Vitulo, N.; Forcato, C.; D’Angelo, M.; Schiavon, R.; Vezzi, A.; Giacometti, G.M.; Morosinotto, T.; Valle, G. Chromosome Scale Genome Assembly and Transcriptome Profiling of Nannochloropsis gaditana in Nitrogen Depletion. Mol. Plant 2014, 7, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; El Hajjami, M.; Shen, C.; You, W.; Lu, Y.; Li, J.; Jing, X.; Hu, Q.; Zhou, W.; Poetsch, A.; et al. Transcriptomic and proteomic responses to very low CO2 suggest multiple carbon concentrating mechanisms in Nannochloropsis oceanica. Biotechnol. Biofuels 2019, 12, 168. [Google Scholar] [CrossRef] [PubMed]
- Hulatt, C.J.; Smolina, I.; Dowle, A.; Kopp, M.; Vasanth, G.K.; Hoarau, G.G.; Wijffels, R.H.; Kiron, V. Proteomic and Transcriptomic Patterns during Lipid Remodeling in Nannochloropsis gaditana. Int. J. Mol. Sci. 2020, 21, 6946. [Google Scholar] [CrossRef]
- Wei, L.; You, W.; Gong, Y.; El Hajjami, M.; Liang, W.; Xu, J.; Poetsch, A. Transcriptomic and proteomic choreography in response to light quality variation reveals key adaption mechanisms in marine Nannochloropsis oceanica. Sci. Total Environ. 2020, 720, 137667. [Google Scholar] [CrossRef] [PubMed]
- You, W.; Wei, L.; Gong, Y.; Hajjami, M.E.; Xu, J.; Poetsch, A. Integration of proteome and transcriptome refines key molecular processes underlying oil production in Nannochloropsis oceanica. Biotechnol. Biofuels 2020, 13, 109. [Google Scholar] [CrossRef]
- Poliner, E.; Farré, E.M.; Benning, C. Advanced genetic tools enable synthetic biology in the oleaginous microalgae Nannochloropsis sp. Plant Cell Rep. 2018, 37, 1383–1399. [Google Scholar] [CrossRef]
- Chen, H.L.; Li, S.S.; Huang, R.; Tsai, H.J. Conditional production of a functional fish growth hormone in the transgenic line of Nannochloropsis oculata (Eustigmatophyceae). J. Phycol. 2008, 44, 768–776. [Google Scholar] [CrossRef]
- Kilian, O.; Benemann, C.S.; Niyogi, K.K.; Vick, B. High-efficiency homologous recombination in the oil-producing alga Nannochloropsis sp. Proc. Natl. Acad. Sci. USA 2011, 108, 21265–21269. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gao, D.; Hu, H. High-efficiency nuclear transformation of the oleaginous marine Nannochloropsis species using PCR product. Biosci. Biotechnol. Biochem. 2014, 78, 812–817. [Google Scholar] [CrossRef]
- Cha, T.-S.; Chen, C.-F.; Yee, W.; Aziz, A.; Loh, S.-H. Cinnamic acid, coumarin and vanillin: Alternative phenolic compounds for efficient Agrobacterium-mediated transformation of the unicellular green alga, Nannochloropsis sp. J. Microbiol. Methods 2011, 84, 430–434. [Google Scholar] [CrossRef]
- Kang, N.K.; Choi, G.-G.; Kim, E.K.; Shin, S.-E.; Jeon, S.; Park, M.S.; Jeong, K.J.; Jeong, B.-r.; Chang, Y.K.; Yang, J.-W.; et al. Heterologous overexpression of sfCherry fluorescent protein in Nannochloropsis salina. Biotechnol. Rep. 2015, 8, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.K.; Jeon, S.; Kwon, S.; Koh, H.G.; Shin, S.-E.; Lee, B.; Choi, G.-G.; Yang, J.-W.; Jeong, B.-r.; Chang, Y.K. Effects of overexpression of a bHLH transcription factor on biomass and lipid production in Nannochloropsis salina. Biotechnol. Biofuels 2015, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wang, Q.T.; Xin, Y.; Lu, Y.D.; Xu, J. Enhancing photosynthetic biomass productivity of industrial oleaginous microalgae by overexpression of RuBisCO activase. Algal Res.-Biomass Biofuels Bioprod. 2017, 27, 366–375. [Google Scholar] [CrossRef]
- Kang, N.K.; Kim, E.K.; Kim, Y.U.; Lee, B.; Jeong, W.-J.; Jeong, B.-R.; Chang, Y.K. Increased lipid production by heterologous expression of AtWRI1 transcription factor in Nannochloropsis salina. Biotechnol. Biofuels 2017, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Zienkiewicz, K.; Zienkiewicz, A.; Poliner, E.; Du, Z.-Y.; Vollheyde, K.; Herrfurth, C.; Marmon, S.; Farré, E.M.; Feussner, I.; Benning, C. Nannochloropsis, a rich source of diacylglycerol acyltransferases for engineering of triacylglycerol content in different hosts. Biotechnol. Biofuels 2017, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Moog, D.; Stork, S.; Reislöhner, S.; Grosche, C.; Maier, U.G. In vivo localization studies in the stramenopile alga Nannochloropsis oceanica. Protist 2015, 166, 161–171. [Google Scholar] [CrossRef]
- Poliner, E.; Panchy, N.; Newton, L.; Wu, G.; Lapinsky, A.; Bullard, B.; Zienkiewicz, A.; Benning, C.; Shiu, S.-H.; Farré, E.M. Transcriptional coordination of physiological responses in Nannochloropsis oceanica CCMP1779 under light/dark cycles. Plant J. 2015, 83, 1097–1113. [Google Scholar] [CrossRef] [PubMed]
- Poliner, E.; Clark, E.; Cummings, C.; Benning, C.; Farre, E.M. A high-capacity gene stacking toolkit for the oleaginous microalga, Nannochloropsis oceanica CCMP1779. Algal Res. 2020, 45, 101664. [Google Scholar] [CrossRef]
- Iwai, M.; Hori, K.; Sasaki-Sekimoto, Y.; Shimojima, M.; Ohta, H. Manipulation of oil synthesis in Nannochloropsis strain NIES-2145 with a phosphorus starvation–inducible promoter from Chlamydomonas reinhardtii. Front. Microbiol. 2015, 6, 912. [Google Scholar] [CrossRef]
- Jackson, H.O.; Berepiki, A.; Baylay, A.J.; Terry, M.J.; Moore, C.M.; Bibby, T.S. An inducible expression system in the alga Nannochloropsis gaditana controlled by the nitrate reductase promoter. J. Appl. Phycol. 2019, 31, 269–279. [Google Scholar] [CrossRef]
- de Grahl, I.; Rout, S.S.; Maple-Grødem, J.; Reumann, S. Development of a constitutive and an auto-inducible high-yield expression system for recombinant protein production in the microalga Nannochloropsis oceanica. Appl. Microbiol. Biotechnol. 2020, 104, 8747–8760. [Google Scholar] [CrossRef]
- Kaye, Y.; Grundman, O.; Leu, S.; Zarka, A.; Zorin, B.; Didi-Cohen, S.; Khozin-Goldberg, I.; Boussiba, S. Metabolic engineering toward enhanced LC-PUFA biosynthesis in Nannochloropsis oceanica: Overexpression of endogenous Δ12 desaturase driven by stress-inducible promoter leads to enhanced deposition of polyunsaturated fatty acids in TAG. Algal Res. 2015, 11, 387–398. [Google Scholar] [CrossRef]
- Radakovits, R.; Jinkerson, R.E.; Fuerstenberg, S.I.; Tae, H.; Settlage, R.E.; Boore, J.L.; Posewitz, M.C. Draft genome sequence and genetic transformation of the oleaginous alga Nannochloropsis gaditana. Nat. Commun. 2012, 3, 686. [Google Scholar] [CrossRef]
- Kwon, S.; Kang, N.K.; Koh, H.G.; Shin, S.E.; Lee, B.; Jeong, B.R.; Chang, Y.K. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2018, 115, 331–340. [Google Scholar] [CrossRef]
- Ma, X.; Yao, L.; Yang, B.; Lee, Y.K.; Chen, F.; Liu, J. RNAi-mediated silencing of a pyruvate dehydrogenase kinase enhances triacylglycerol biosynthesis in the oleaginous marine alga Nannochloropsis salina. Sci. Rep. 2017, 7, 11485. [Google Scholar] [CrossRef]
- Südfeld, C.; Hubáček, M.; Figueiredo, D.; Naduthodi, M.I.S.; van der Oost, J.; Wijffels, R.H.; Barbosa, M.J.; D’Adamo, S. High-throughput insertional mutagenesis reveals novel targets for enhancing lipid accumulation in Nannochloropsis oceanica. Metab. Eng. 2021, 66, 239–258. [Google Scholar] [CrossRef]
- Ramarajan, M.; Fabris, M.; Abbriano, R.M.; Pernice, M.; Ralph, P.J. Novel endogenous promoters for genetic engineering of the marine microalga Nannochloropsis gaditana CCMP526. Algal Res. 2019, 44, 101708. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, L.; Liu, H.; Pan, X.; Liu, L.; Zong, Y.; Yang, G. Zeocin treatment significantly elevated transformation efficiency of Nannochloropsis oceanica. J. Appl. Phycol. 2022, 34, 1587–1594. [Google Scholar] [CrossRef]
- Tsien, R.Y. THE GREEN FLUORESCENT PROTEIN. Annu. Rev. Biochem. 1998, 67, 509–544. [Google Scholar] [CrossRef]
- Poliner, E.; Pulman, J.A.; Zienkiewicz, K.; Childs, K.; Benning, C.; Farré, E.M. A toolkit for Nannochloropsis oceanica CCMP1779 enables gene stacking and genetic engineering of the eicosapentaenoic acid pathway for enhanced long-chain polyunsaturated fatty acid production. Plant Biotechnol. J. 2018, 16, 298–309. [Google Scholar] [CrossRef]
- Shih, C.H.; Chen, H.Y.; Lee, H.C.; Tsai, H.J. Purple chromoprotein gene serves as a new selection marker for transgenesis of the microalga Nannochloropsis oculata. PLoS ONE 2015, 10, e0120780. [Google Scholar] [CrossRef]
- Wei, H.; Shi, Y.; Ma, X.; Pan, Y.; Hu, H.; Li, Y.; Luo, M.; Gerken, H.; Liu, J. A type-I diacylglycerol acyltransferase modulates triacylglycerol biosynthesis and fatty acid composition in the oleaginous microalga, Nannochloropsis oceanica. Biotechnol. Biofuels 2017, 10, 174. [Google Scholar] [CrossRef]
- Poliner, E.; Busch, A.W.U.; Newton, L.; Kim, Y.U.; Clark, R.; Gonzalez-Martinez, S.C.; Jeong, B.-R.; Montgomery, B.L.; Farré, E.M. Aureochromes maintain polyunsaturated fatty acid content in Nannochloropsis oceanica. Plant Physiol. 2022, 189, 906–921. [Google Scholar] [CrossRef]
- Jiang, W.; Brueggeman, A.J.; Horken, K.M.; Plucinak, T.M.; Weeks, D.P. Successful Transient Expression of Cas9 and Single Guide RNA Genes in Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 1465–1469. [Google Scholar] [CrossRef]
- Baek, K.; Kim, D.H.; Jeong, J.; Sim, S.J.; Melis, A.; Kim, J.S.; Jin, E.; Bae, S. DNA-free two-gene knockout in Chlamydomonas reinhardtii via CRISPR-Cas9 ribonucleoproteins. Sci. Rep. 2016, 6, 30620. [Google Scholar] [CrossRef]
- Kim, S.; Kim, D.; Cho, S.W.; Kim, J.; Kim, J.-S. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome Res. 2014, 24, 1012–1019. [Google Scholar] [CrossRef]
- Zuris, J.A.; Thompson, D.B.; Shu, Y.; Guilinger, J.P.; Bessen, J.L.; Hu, J.H.; Maeder, M.L.; Joung, J.K.; Chen, Z.-Y.; Liu, D.R. Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo. Nat. Biotechnol. 2015, 33, 73–80. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Subburaj, S.; Chung, S.J.; Lee, C.; Ryu, S.-M.; Kim, D.H.; Kim, J.-S.; Bae, S.; Lee, G.-J. Site-directed mutagenesis in Petunia × hybrida protoplast system using direct delivery of purified recombinant Cas9 ribonucleoproteins. Plant Cell Rep. 2016, 35, 1535–1544. [Google Scholar] [CrossRef]
- Murakami, H.; Kakutani, N.; Kuroyanagi, Y.; Iwai, M.; Hori, K.; Shimojima, M.; Ohta, H. MYB-like transcription factor NoPSR1 is crucial for membrane lipid remodeling under phosphate starvation in the oleaginous microalga Nannochloropsis oceanica. FEBS Lett. 2020, 594, 3384–3394. [Google Scholar] [CrossRef]
- Murakami, H.; Nobusawa, T.; Hori, K.; Shimojima, M.; Ohta, H. Betaine Lipid Is Crucial for Adapting to Low Temperature and Phosphate Deficiency in Nannochloropsis. Plant Physiol. 2018, 177, 181–193. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Plucinak, T.M.; Horken, K.M.; Jiang, W.; Fostvedt, J.; Nguyen, S.T.; Weeks, D.P. Improved and versatile viral 2A platforms for dependable and inducible high-level expression of dicistronic nuclear genes in Chlamydomonas reinhardtii. Plant J. 2015, 82, 717–729. [Google Scholar] [CrossRef]
- Koh, H.G.; Kang, N.K.; Kim, E.K.; Jeon, S.; Shin, S.-E.; Lee, B.; Chang, Y.K. Advanced multigene expression system for Nannochloropsis salina using 2A self-cleaving peptides. J. Biotechnol. 2018, 278, 39–47. [Google Scholar] [CrossRef]
- Vafaee, Y.; Staniek, A.; Mancheno-Solano, M.; Warzecha, H. A Modular Cloning Toolbox for the Generation of Chloroplast Transformation Vectors. PLoS ONE 2014, 9, e110222. [Google Scholar] [CrossRef]
- Daniell, H.; Jin, S.; Zhu, X.-G.; Gitzendanner, M.A.; Soltis, D.E.; Soltis, P.S. Green giant—A tiny chloroplast genome with mighty power to produce high-value proteins: History and phylogeny. Plant Biotechnol. J. 2021, 19, 430–447. [Google Scholar] [CrossRef]
- Scranton, M.A.; Ostrand, J.T.; Fields, F.J.; Mayfield, S.P. Chlamydomonas as a model for biofuels and bio-products production. Plant J. 2015, 82, 523–531. [Google Scholar] [CrossRef]
- Larrea-Alvarez, M.; Purton, S. Multigenic engineering of the chloroplast genome in the green alga Chlamydomonas reinhardtii. Microbiology 2020, 166, 510–515. [Google Scholar] [CrossRef]
- Munjal, N.; Garzon-Sanabria, A.J.; Quinones, K.W.; Gregory, J.; Nikolov, Z.L. Light-Induced Production of an Antibody Fragment and Malaria Vaccine Antigen from Chlamydomonas reinhardtii. Processes 2014, 2, 625–638. [Google Scholar] [CrossRef]
- Gan, Q.; Jiang, J.; Han, X.; Wang, S.; Lu, Y. Engineering the Chloroplast Genome of Oleaginous Marine Microalga Nannochloropsis oceanica. Front. Plant Sci. 2018, 9, 439. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, K.; Xu, W.; Wang, Y.; Gao, Z.; Cui, H.; Meng, C.; Qin, S. Plastid Engineering of a Marine Alga, Nannochloropsis gaditana, for Co-Expression of Two Recombinant Peptides. J. Phycol. 2021, 57, 569–576. [Google Scholar] [CrossRef]
- Perozeni, F.; Baier, T. Current Nuclear Engineering Strategies in the Green Microalga Chlamydomonas reinhardtii. Life 2023, 13, 1566. [Google Scholar] [CrossRef]
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of Triacylglycerol Accumulation in Plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef]
- Li, D.-W.; Balamurugan, S.; Yang, Y.-F.; Zheng, J.-W.; Huang, D.; Zou, L.-G.; Yang, W.-D.; Liu, J.-S.; Guan, Y.; Li, H.-Y. Transcriptional regulation of microalgae for concurrent lipid overproduction and secretion. Sci. Adv. 2019, 5, eaau3795. [Google Scholar] [CrossRef]
- Chen, J.W.; Liu, W.J.; Hu, D.X.; Wang, X.; Balamurugan, S.; Alimujiang, A.; Yang, W.D.; Liu, J.S.; Li, H.Y. Identification of a malonyl CoA-acyl carrier protein transacylase and its regulatory role in fatty acid biosynthesis in oleaginous microalga Nannochloropsis oceanica. Biotechnol. Appl. Biochem. 2017, 64, 620–626. [Google Scholar] [CrossRef]
- Han, X.; Song, X.; Li, F.; Lu, Y. Improving lipid productivity by engineering a control-knob gene in the oleaginous microalga Nannochloropsis oceanica. Metab. Eng. Commun. 2020, 11, e00142. [Google Scholar] [CrossRef]
- Jeon, S.; Koh, H.G.; Cho, J.M.; Kang, N.K.; Chang, Y.K. Enhancement of lipid production in Nannochloropsis salina by overexpression of endogenous NADP-dependent malic enzyme. Algal Res. 2021, 54, 102218. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Pan, Y.; Shi, Y.; Hu, H. Metabolic engineering of the oleaginous alga Nannochloropsis for enriching eicosapentaenoic acid in triacylglycerol by combined pulling and pushing strategies. Metab. Eng. 2022, 69, 163–174. [Google Scholar] [CrossRef]
- Liu, M.; Ding, W.; Yu, L.; Shi, Y.; Liu, J. Functional characterization of carotenogenic genes provides implications into carotenoid biosynthesis and engineering in the marine alga Nannochloropsis oceanica. Algal Res. 2022, 67, 102853. [Google Scholar] [CrossRef]
- Perin, G.; Bellan, A.; Michelberger, T.; Lyska, D.; Wakao, S.; Niyogi, K.K.; Morosinotto, T. Modulation of xanthophyll cycle impacts biomass productivity in the marine microalga Nannochloropsis. Proc. Natl. Acad. Sci. USA 2023, 120, e2214119120. [Google Scholar] [CrossRef]
- Dautermann, O.; Lyska, D.; Andersen-Ranberg, J.; Becker, M.; Fröhlich-Nowoisky, J.; Gartmann, H.; Krämer, L.C.; Mayr, K.; Pieper, D.; Rij, L.M.; et al. An algal enzyme required for biosynthesis of the most abundant marine carotenoids. Sci. Adv. 2020, 6, eaaw9183. [Google Scholar] [CrossRef]
- Liu, M.; Ding, W.; Pan, Y.; Hu, H.; Liu, J. Zeaxanthin epoxidase is involved in the carotenoid biosynthesis and light-dependent growth of the marine alga Nannochloropsis oceanica. Biotechnol. Biofuels Bioprod. 2023, 16, 74. [Google Scholar] [CrossRef]
- Park, S.-B.; Yun, J.-H.; Ryu, A.J.; Yun, J.; Kim, J.W.; Lee, S.; Choi, S.; Cho, D.-H.; Choi, D.-Y.; Lee, Y.J.; et al. Development of a novel nannochloropsis strain with enhanced violaxanthin yield for large-scale production. Microb. Cell Factories 2021, 20, 43. [Google Scholar] [CrossRef]
- Cecchin, M.; Cazzaniga, S.; Martini, F.; Paltrinieri, S.; Bossi, S.; Maffei, M.E.; Ballottari, M. Astaxanthin and eicosapentaenoic acid production by S4, a new mutant strain of Nannochloropsis gaditana. Microb. Cell Factories 2022, 21, 117. [Google Scholar] [CrossRef]
- Koh, H.G.; Kang, N.K.; Jeon, S.; Shin, S.-E.; Jeong, B.-r.; Chang, Y.K. Heterologous synthesis of chlorophyll b in Nannochloropsis salina enhances growth and lipid production by increasing photosynthetic efficiency. Biotechnol. Biofuels 2019, 12, 122. [Google Scholar] [CrossRef]
- Li, S.S.; Tsai, H.J. Transgenic microalgae as a non-antibiotic bactericide producer to defend against bacterial pathogen infection in the fish digestive tract. Fish Shellfish. Immunol. 2009, 26, 316–325. [Google Scholar] [CrossRef]
- Rout, S.S.; de Grahl, I.; Yu, X.; Reumann, S. Production of a viral surface protein in Nannochloropsis oceanica for fish vaccination against infectious pancreatic necrosis virus. Appl. Microbiol. Biotechnol. 2022, 106, 6535–6549. [Google Scholar] [CrossRef]
- Lauersen, K.J. Eukaryotic microalgae as hosts for light-driven heterologous isoprenoid production. Planta 2019, 249, 155–180. [Google Scholar] [CrossRef]
- Wichmann, J.; Eggert, A.; Elbourne, L.D.H.; Paulsen, I.T.; Lauersen, K.J.; Kruse, O. Farnesyl pyrophosphate compartmentalization in the green microalga Chlamydomonas reinhardtii during heterologous (E)-α-bisabolene production. Microb. Cell Factories 2022, 21, 190. [Google Scholar] [CrossRef]
- Einhaus, A.; Steube, J.; Freudenberg, R.A.; Barczyk, J.; Baier, T.; Kruse, O. Engineering a powerful green cell factory for robust photoautotrophic diterpenoid production. Metab. Eng. 2022, 73, 82–90. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Overmans, S.; Wellman, G.B.; Samaras, V.G.; Oviedo, C.; Gede, M.; Szekely, G.; Lauersen, K.J. A green alternative to fragrant agarwood sesquiterpenoid production. bioRxiv 2023. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Bhat, W.W.; Poliner, E.; Johnson, S.; Bertucci, C.; Farre, E.; Hamberger, B. Engineering Nannochloropsis oceanica for the production of diterpenoid compounds. mLife 2023, 2, 428–437. [Google Scholar] [CrossRef]
- Perin, G.; Bellan, A.; Segalla, A.; Meneghesso, A.; Alboresi, A.; Morosinotto, T. Generation of random mutants to improve light-use efficiency of Nannochloropsis gaditana cultures for biofuel production. Biotechnol. Biofuels 2015, 8, 161. [Google Scholar] [CrossRef]
- Arora, N.; Lo, E.; Philippidis, G.P. Dissecting Enhanced Carbohydrate and Pigment Productivity in Mutants of Nannochloropsis oculata Using Metabolomics and Lipidomics. J. Agric. Food Chem. 2022, 70, 8338–8350. [Google Scholar] [CrossRef]
- Kang, N.K.; Kim, E.K.; Sung, M.G.; Kim, Y.U.; Jeong, B.R.; Chang, Y.K. Increased biomass and lipid production by continuous cultivation of Nannochloropsis salina transformant overexpressing a bHLH transcription factor. Biotechnol. Bioeng. 2019, 116, 555–568. [Google Scholar] [CrossRef]
Figure 1.
Nannochloropsis phylogenetic tree. Phylogenetic tree showing the evolution of Nannochloropsis spp. with respect to the other algae. The phylogenetic tree is obtained from the multiple sequence alignment of 18S ribosomal RNA gene (18S rDNA) using Clustal omega and neighbor-joining distance matrix. The following sequences from NCBI were used: Physcomitrella patens (AF126289.1), Arabidopsis thaliana (X16077.1), Chlamydomonas reinhardtii (AB511835.1), Chlorella vulgaris (X13688.1), Cyanidioshyzon merolae (XR_002461616.1), Thalassiosira pseudonana (MH545685.1), Phaeodactylum tricornutum (AJ269501.1), Nannochloropsis CCMP505 (U41050.1), and Ectocarpus siliculosus (L43062.1).
Figure 1.
Nannochloropsis phylogenetic tree. Phylogenetic tree showing the evolution of Nannochloropsis spp. with respect to the other algae. The phylogenetic tree is obtained from the multiple sequence alignment of 18S ribosomal RNA gene (18S rDNA) using Clustal omega and neighbor-joining distance matrix. The following sequences from NCBI were used: Physcomitrella patens (AF126289.1), Arabidopsis thaliana (X16077.1), Chlamydomonas reinhardtii (AB511835.1), Chlorella vulgaris (X13688.1), Cyanidioshyzon merolae (XR_002461616.1), Thalassiosira pseudonana (MH545685.1), Phaeodactylum tricornutum (AJ269501.1), Nannochloropsis CCMP505 (U41050.1), and Ectocarpus siliculosus (L43062.1).

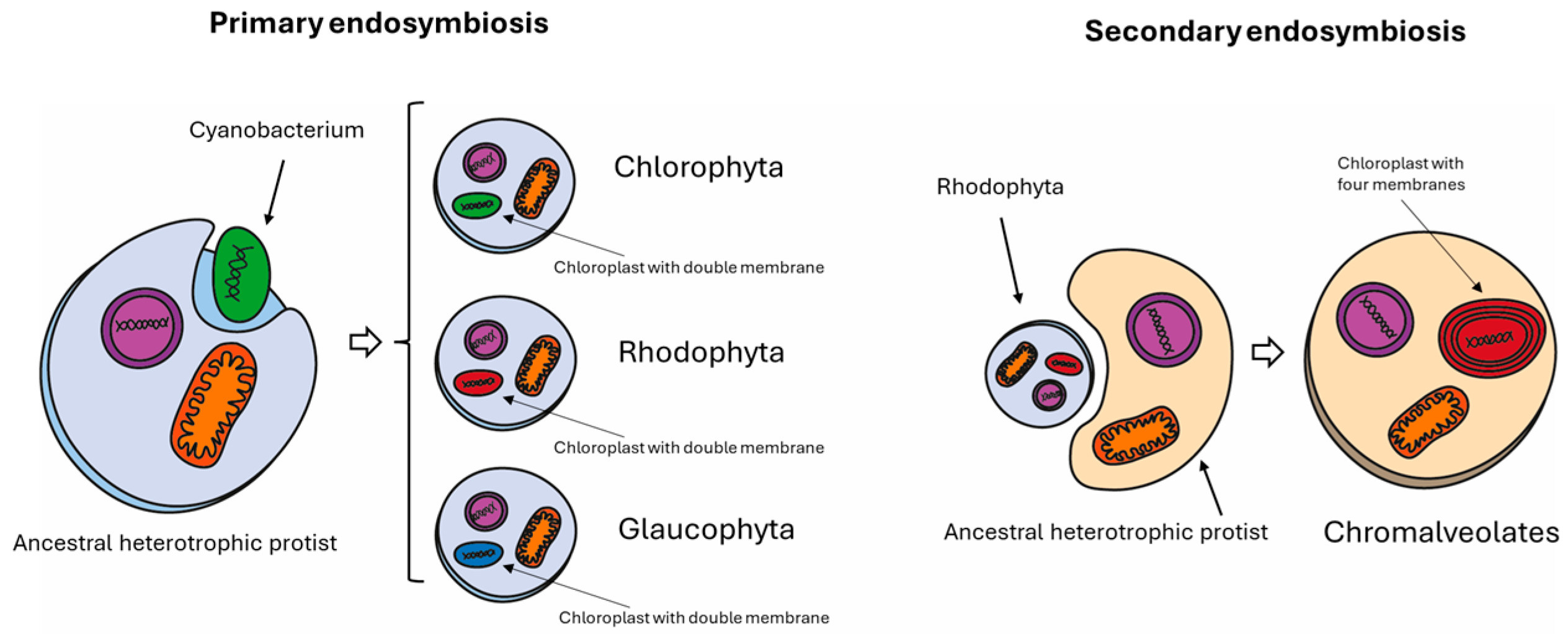
Figure 2.
Diagram showing primary and secondary endosymbiosis events at the origin of Nannocchloropsis spp. The left panel shows the primary endosymbiosis at the origin of green algae (Chlorophyta), red algae (Rhodophyta), and Glaucophytes. In this process, a eukaryotic cell engulfs a prokaryotic cell that can undergo photosynthesis. As a footprint, the chloroplast of cells belonging to primary endosymbiosis is surrounded by a double membrane. By contrast, Chromalveolates, which include Cryptophyta, Haptophyta, Stramenopiles (or Heterokontophyta), and Alveolata, were originated by a secondary endosymbiosis. In this process, an eukaryotic cell engulfs another eukaryotic cell which has already engulfed a prokaryotic cell in its past (a Rhodophyte in that case). The Chromalveolate chloroplast, as a result of secondary endosymbiosis, is surrounded by four membranes.
Figure 2.
Diagram showing primary and secondary endosymbiosis events at the origin of Nannocchloropsis spp. The left panel shows the primary endosymbiosis at the origin of green algae (Chlorophyta), red algae (Rhodophyta), and Glaucophytes. In this process, a eukaryotic cell engulfs a prokaryotic cell that can undergo photosynthesis. As a footprint, the chloroplast of cells belonging to primary endosymbiosis is surrounded by a double membrane. By contrast, Chromalveolates, which include Cryptophyta, Haptophyta, Stramenopiles (or Heterokontophyta), and Alveolata, were originated by a secondary endosymbiosis. In this process, an eukaryotic cell engulfs another eukaryotic cell which has already engulfed a prokaryotic cell in its past (a Rhodophyte in that case). The Chromalveolate chloroplast, as a result of secondary endosymbiosis, is surrounded by four membranes.

Figure 3.
CRISPR/Cas9 mechanism. The main components are an RNA-guided Cas9 endonuclease and a single guide RNA (sgRNA). The genome integration-based approach (left side) requires the construction of a vector carrying three different cassettes: one encoding for the Cas9 protein, the second for the sgRNA, and the third for antibiotic resistance. Once the vector is stably integrated into the host genome, Cas9 protein and sgRNA are synthesized through cell machinery. On the contrary, the in vitro ribonucleoprotein (RNP) assembly approach (right side) requires Cas9 purified protein, usually from E. coli, and in vitro synthesized sgRNA. Both in the integration and in the in vitro RNP assembly, Cas9 protein and sgRNA are assembled to form Cas9 (RNP) with the ability to bind and cleave target DNA [92]. The main difference is that in the former, the RNP complex is assembled by the cell itself, while in the latter, RNP is in vitro assembled and later inserted into the cell. Single guide RNA drives the RNP to the targeted site that must be close to the protospacer adjacent motif (PAM) [93]. The cleavage carried out by Cas9 is a double-strand blunt break (DSB). Once the DSB is introduced in the target gene by Cas9 nuclease activity, the cell has two alternatives for the repair: non-homologous end-joining (NHEJ) or homology-directed repair (HDR) [93]. The first can introduce insertion or deletion at cleavage sites, generating a frameshift into the gene open reading frame. The latter can introduce exogenous DNA at the target site by using a homologous DNA repair template. In the HDR, usually, as donor DNA, the resistant marker cassette is used to allow transformant screening.
Figure 3.
CRISPR/Cas9 mechanism. The main components are an RNA-guided Cas9 endonuclease and a single guide RNA (sgRNA). The genome integration-based approach (left side) requires the construction of a vector carrying three different cassettes: one encoding for the Cas9 protein, the second for the sgRNA, and the third for antibiotic resistance. Once the vector is stably integrated into the host genome, Cas9 protein and sgRNA are synthesized through cell machinery. On the contrary, the in vitro ribonucleoprotein (RNP) assembly approach (right side) requires Cas9 purified protein, usually from E. coli, and in vitro synthesized sgRNA. Both in the integration and in the in vitro RNP assembly, Cas9 protein and sgRNA are assembled to form Cas9 (RNP) with the ability to bind and cleave target DNA [92]. The main difference is that in the former, the RNP complex is assembled by the cell itself, while in the latter, RNP is in vitro assembled and later inserted into the cell. Single guide RNA drives the RNP to the targeted site that must be close to the protospacer adjacent motif (PAM) [93]. The cleavage carried out by Cas9 is a double-strand blunt break (DSB). Once the DSB is introduced in the target gene by Cas9 nuclease activity, the cell has two alternatives for the repair: non-homologous end-joining (NHEJ) or homology-directed repair (HDR) [93]. The first can introduce insertion or deletion at cleavage sites, generating a frameshift into the gene open reading frame. The latter can introduce exogenous DNA at the target site by using a homologous DNA repair template. In the HDR, usually, as donor DNA, the resistant marker cassette is used to allow transformant screening.

Figure 4.
Screening pipeline for transformants using the “safe harbor” system. Briefly, genes of interest are in silico designed, optimizing codon usage and adding homologous recombination sequences (1). Nannochloropsis landing pad strain nuclear transformation is performed by means of electroporation, and homologous recombination (HR) occurs in the safe harbor site (2). td Tomato fluorescence can be used to identify lines in which homologous recombination occurred. WT shows no fluorescence, background landing pad strains (BG) show maximum fluorescence, while the transformed lines have or do not have fluorescence according to the occurrence of HR (3). Protein quantification allows quantifying the protein of interest; lines showing no tomato fluorescence, in which HR occurs in the safe harbor site, show the highest protein accumulation; other transformed lines show no or lower protein accumulation due to random insertion into the genome (4).
Figure 4.
Screening pipeline for transformants using the “safe harbor” system. Briefly, genes of interest are in silico designed, optimizing codon usage and adding homologous recombination sequences (1). Nannochloropsis landing pad strain nuclear transformation is performed by means of electroporation, and homologous recombination (HR) occurs in the safe harbor site (2). td Tomato fluorescence can be used to identify lines in which homologous recombination occurred. WT shows no fluorescence, background landing pad strains (BG) show maximum fluorescence, while the transformed lines have or do not have fluorescence according to the occurrence of HR (3). Protein quantification allows quantifying the protein of interest; lines showing no tomato fluorescence, in which HR occurs in the safe harbor site, show the highest protein accumulation; other transformed lines show no or lower protein accumulation due to random insertion into the genome (4).

Figure 5.
Nannochloropsis lipid metabolism. The main characterized enzymes are written in dark blue. Abbreviations: ACCase, acetyl-CoA carboxylase; DGAT, diacylglycerol acyltransferase; DGD1, digalactosyldiacylglycerol synthase; FAD, fatty acid desaturase; FAS, fatty acid synthase; FAE, fatty acid elongase; GPAT, glycerol 3-phosphate acyltransferase; LPAT, lysophosphatidic acid acyltransferase; MGD1, monogalactosyldiacylglycerol synthase; PAP, phosphatidic acid phosphatase; PDAT, phospholipid:diacylglycerol acyltransferase; PDHC, pyruvate dehydrogenase complex; SQD1, UDP-sulfoquinovose synthase 1; CoA, coenzyme A; DGDG, digalactosyldiacylglycerol; G3P, glycerol 3-phosphate; MGDG, monogalactosyldiacylglycerol; SQDG, sulfoquinovosyldiacylglycerol; TAG, triacylglycerol; PUFA, polyunsaturated fatty acid.
Figure 5.
Nannochloropsis lipid metabolism. The main characterized enzymes are written in dark blue. Abbreviations: ACCase, acetyl-CoA carboxylase; DGAT, diacylglycerol acyltransferase; DGD1, digalactosyldiacylglycerol synthase; FAD, fatty acid desaturase; FAS, fatty acid synthase; FAE, fatty acid elongase; GPAT, glycerol 3-phosphate acyltransferase; LPAT, lysophosphatidic acid acyltransferase; MGD1, monogalactosyldiacylglycerol synthase; PAP, phosphatidic acid phosphatase; PDAT, phospholipid:diacylglycerol acyltransferase; PDHC, pyruvate dehydrogenase complex; SQD1, UDP-sulfoquinovose synthase 1; CoA, coenzyme A; DGDG, digalactosyldiacylglycerol; G3P, glycerol 3-phosphate; MGDG, monogalactosyldiacylglycerol; SQDG, sulfoquinovosyldiacylglycerol; TAG, triacylglycerol; PUFA, polyunsaturated fatty acid.

Table 2.
List of available selection markers.
| Protein | Conferring Resistance to | Reference |
|---|---|---|
| Bleomycin resistance protein (Sh ble) | Zeomycin | [58] |
| Hygromycin-B phosphotransferase (AphVII) | Hygromycin | [39] |
| Blasticidin-S deaminase (BSD) | Blasticidin | [68] |
| Nourseothricin acetyltransferase (NAT) | Nourseothricin | [68] |
| Aminoglycoside 3′ phosphotransferase | G418 | [68] |
Table 3.
Most commonly used fluorescent reporters in Nannochloropsis spp. for screening. Excitation and emission maximum wavelength and brightness are reported. Data from FPbase (www.FPbase.org) (accessed on 24 March 2024).
Table 3.
Most commonly used fluorescent reporters in Nannochloropsis spp. for screening. Excitation and emission maximum wavelength and brightness are reported. Data from FPbase (www.FPbase.org) (accessed on 24 March 2024).
| Name | Ex λ | Em λ | Brightness | Excitation/Emission Spectra |
|---|---|---|---|---|
| Clover (GFP) | 505 | 515 | 84.36 |  |
| mVenus (YFP) | 515 | 527 | 66.56 |  |
| mCerulean (CFP) | 433 | 475 | 34.8 |  |
| sfCherry | 589 | 610 | 17.47 |  |
| tdTomato | 554 | 581 | 95.22 |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Canini, D.; Ceschi, E.; Perozeni, F. Toward the Exploitation of Sustainable Green Factory: Biotechnology Use of Nannochloropsis spp. Biology 2024, 13, 292. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13050292
AMA Style
Canini D, Ceschi E, Perozeni F. Toward the Exploitation of Sustainable Green Factory: Biotechnology Use of Nannochloropsis spp. Biology. 2024; 13(5):292. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13050292
Chicago/Turabian StyleCanini, Davide, Edoardo Ceschi, and Federico Perozeni. 2024. "Toward the Exploitation of Sustainable Green Factory: Biotechnology Use of Nannochloropsis spp." Biology 13, no. 5: 292. https://0-doi-org.brum.beds.ac.uk/10.3390/biology13050292
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.