

Differential Expression of LMNA/C and Insulin Receptor Transcript Variants in Peripheral Blood Mononuclear Cells of Leukemia Patients
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Demographic Data of Subjects
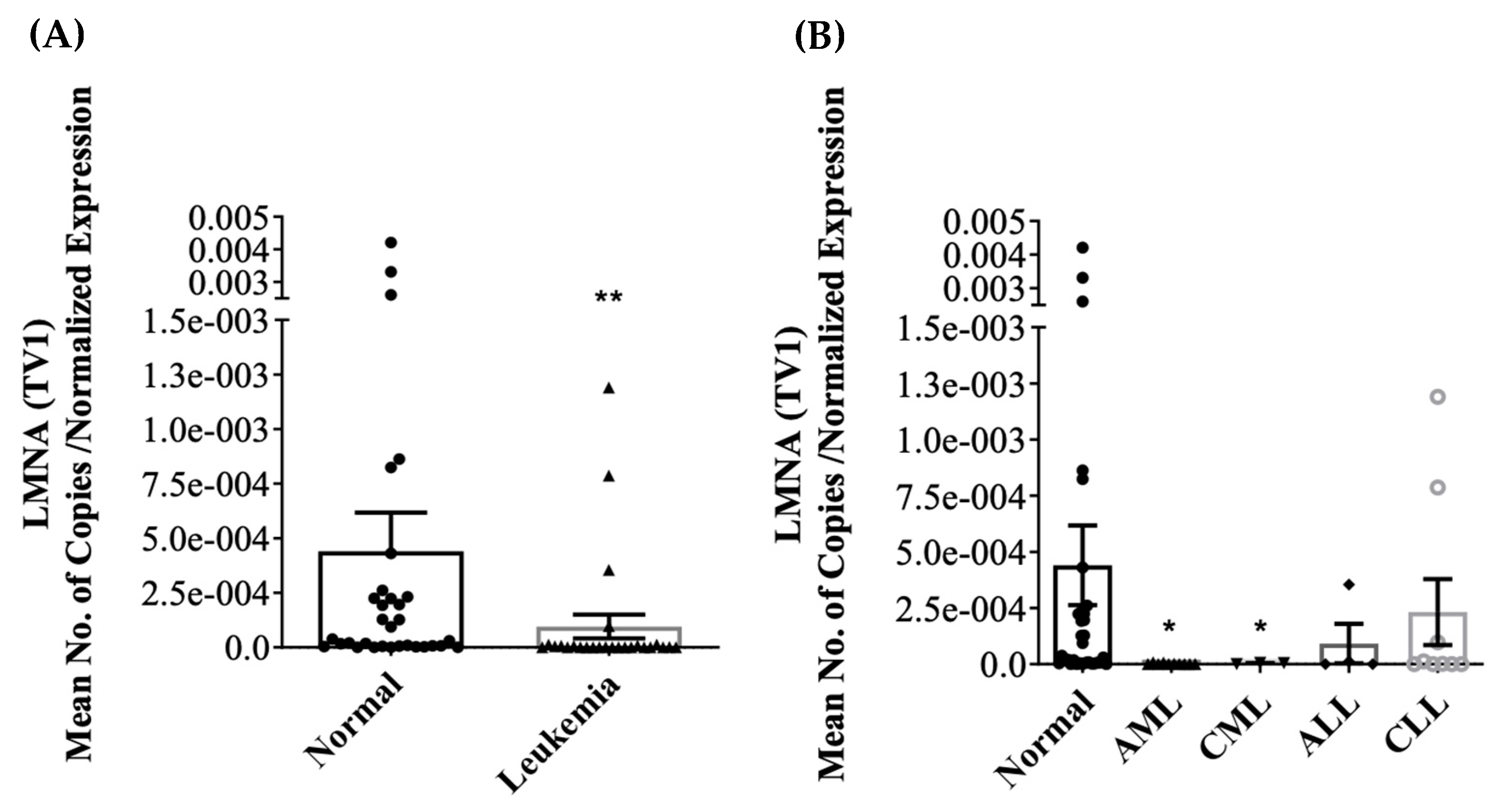
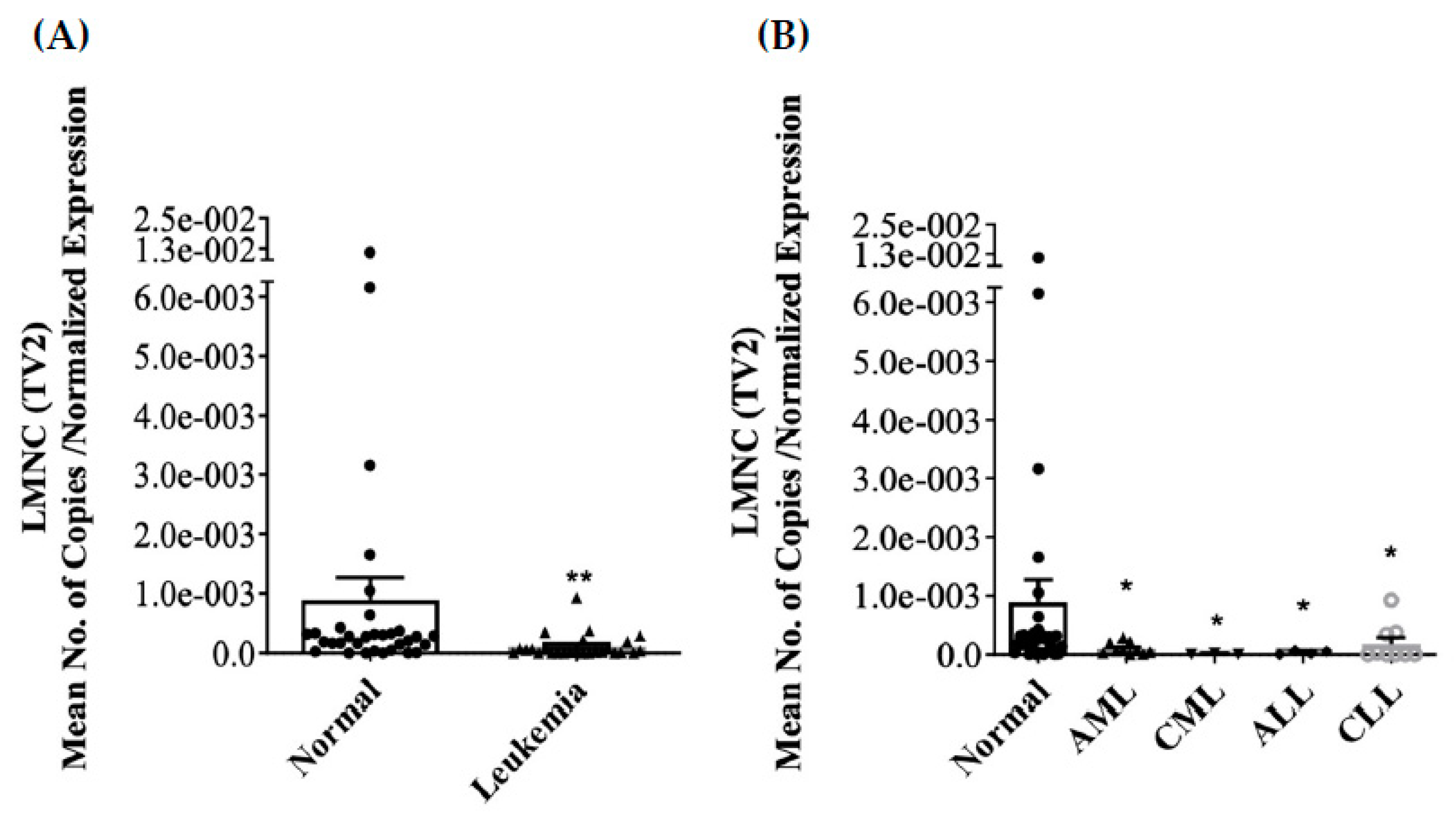
3.2. Differential Expression of LMNA/C mRNA Expression in PBMC
3.3. Inhibition of INSR (IR-A) mRNA Expression in PBMC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamid, G.A. Classification of acute leukemia. Ann. Intern. Med. 2011, 87, 740–753. [Google Scholar]
- Laosai, J.; Chamnongthai, K. Classification of acute leukemia using medical-knowledge-based morphology and CD marker. Biomed. Signal Process. Control 2018, 44, 127–137. [Google Scholar] [CrossRef]
- Mckenna, R.W. Multifaceted approach to the diagnosis and classification of acute leukemias. Clin. Chem. 2000, 46 Pt 2, 1252–1259. [Google Scholar] [CrossRef]
- Taylor, C.G.; Stasi, R.; Bastianelli, C.; Venditti, A.; del Poeta, G.; Amadori, S.; Sargent, J.M. Diagnosis and classification of the acute leukemias: Recent advances and controversial issues. Hematop. Mol. Hematol. 1996, 10, 1–38. [Google Scholar]
- Olsen, R.J.; Chang, C.-C.J.; Herrick, J.; Zu, Y.; Ehsan, A. Acute leukemia immunohistochemistry: A systematic diagnostic approach. Arch. Pathol. Lab. Med. 2008, 132, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Foon, K.A.; Gale, R.P.; Todd, R.F. Recent advances in the immunologic classification of leukemia. Semin. Hematol. 1986, 23, 257–283. [Google Scholar] [PubMed]
- Sobol, R.E.; Royston, I.; Lebien, T.W.; Minowada, J.; Anderson, K.; Davey, F.R.; Cuttner, J.; Schiffer, C.A.; Ellison, R.R.; Bloomfield, C.D. Adult acute lymphoblastic leukemia phenotypes defined by monoclonal antibodies. Blood 1985, 65, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Pui, C.-H.; Carroll, W.L.; Meshinchi, S.; Arceci, R.J. Biology, risk stratification, and therapy of pediatric acute leukemias: An update. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Yohe, S.L. Molecular Genetic Markers in Acute Myeloid Leukemia. J. Clin. Med. 2015, 4, 460–478. [Google Scholar] [CrossRef]
- Mullighan, C.G.; Flotho, C.; Downing, J.R. Genomic Assessment of Pediatric Acute Leukemia. Cancer J. 2005, 11, 268–282. [Google Scholar] [CrossRef]
- Tefferi, A.; Dewald, G.W.; Litzow, M.; Cortes, J.E.; Mauro, M.J.; Talpaz, M.; Kantarjian, H.M. Chronic myeloid leukemia: Current application of cytogenetics and molecular testing for diagnosis and treatment. Mayo Clin. Proc. 2005, 80, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Dechat, T.; Adam, S.A.; Taimen, P.; Shimi, T.; Goldman, R.D. Nuclear lamins. Cold Spring Harb. Perspect. Biol. 2010, 2, a000547. [Google Scholar] [CrossRef] [PubMed]
- Stuurman, N.; Heins, S.; Aebi, U. Nuclear lamins: Their structure, assembly, and interactions. J. Struct. Biol. 1998, 122, 42–66. [Google Scholar] [CrossRef] [PubMed]
- Butin-Israeli, V.; Adam, S.A.; Goldman, A.E.; Goldman, R.D. Nuclear lamin functions and disease. Trends Genet. TIG 2012, 28, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Prokocimer, M.; Margalit, A.; Gruenbaum, Y. The nuclear lamina and its proposed roles in tumorigenesis: Projection on the hematologic malignancies and future targeted therapy. J. Struct. Biol. 2006, 155, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Irianto, J.; Pfeifer, C.R.; Ivanovska, I.L.; Swift, J.; Discher, D.E. Nuclear Lamins in Cancer. Cell. Mol. Bioeng. 2016, 9, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Aljada, A.; Doria, J.; Saleh, A.M.; Al-Matar, S.H.; AlGabbani, S.; Shamsa, H.B.; Al-Bawab, A.; Ahmed, A.A. Altered Lamin A/C splice variant expression as a possible diagnostic marker in breast cancer. Cell. Oncol. 2016, 39, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef]
- Belfiore, A.; Malaguarnera, R. Insulin receptor and cancer. Endocr.-Relat. Cancer 2011, 18, R125–R147. [Google Scholar] [CrossRef]
- Vivekanandhan, S.; Mukhopadhyay, D. Genetic status of KRAS influences Transforming Growth Factor-beta (TGF-β) signaling: An insight into Neuropilin-1 (NRP1) mediated tumorigenesis. Semin. Cancer Biol. 2018, 54, 72–79. [Google Scholar] [CrossRef]
- Ghoshal Gupta, S.; Baumann, H.; Wetzler, M. Epigenetic regulation of signal transducer and activator of transcription 3 in acute myeloid leukemia. Leuk. Res. 2008, 32, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Benekli, M.; Baumann, H.; Wetzler, M. Targeting signal transducer and activator of transcription signaling pathway in leukemias. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 4422–4432. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Chen, Y.-G. Interplay between TGF-β signaling and receptor tyrosine kinases in tumor development. Sci. China Life Sci. 2017, 60, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Frasca, F.; Pandini, G.; Sciacca, L.; Pezzino, V.; Squatrito, S.; Belfiore, A.; Vigneri, R. The role of insulin receptors and IGF-I receptors in cancer and other diseases. Arch. Physiol. Biochem. 2008, 114, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Galal, M.A.; Alouch, S.S.; Alsultan, B.S.; Dahman, H.; Alyabis, N.A.; Alammar, S.A.; Aljada, A. Insulin Receptor Isoforms and Insulin Growth Factor-like Receptors: Implications in Cell Signaling, Carcinogenesis, and Chemoresistance. Int. J. Mol. Sci. 2023, 24, 15006. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, A. The role of insulin receptor isoforms and hybrid insulin/IGF-I receptors in human cancer. Curr. Pharm. Des. 2007, 13, 671–686. [Google Scholar] [CrossRef]
- Masson, K.; Rönnstrand, L. Oncogenic signaling from the hematopoietic growth factor receptors c-Kit and Flt3. Cell. Signal. 2009, 21, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- De Meyts, P. The insulin receptor isoform A: A mitogenic proinsulin receptor? Endocrinology 2012, 153, 2054–2056. [Google Scholar] [CrossRef] [PubMed]
- Vella, V.; Milluzzo, A.; Scalisi, N.M.; Vigneri, P.; Sciacca, L. Insulin Receptor Isoforms in Cancer. Int. J. Mol. Sci. 2018, 19, 3615. [Google Scholar] [CrossRef]
- Aljada, A.; Saleh, A.M.; Al-Aqeel, S.M.; Shamsa, H.B.; Al-Bawab, A.; Al Dubayee, M.; Ahmed, A.A. Quantification of insulin receptor mRNA splice variants as a diagnostic tumor marker in breast cancer. Cancer Biomark. Sect. A Dis. Markers 2015, 15, 653–661. [Google Scholar] [CrossRef]
- Sciacca, L.; Prisco, M.; Wu, A.; Belfiore, A.; Vigneri, R.; Baserga, R. Signaling differences from the A and B isoforms of the insulin receptor (IR) in 32D cells in the presence or absence of IR substrate-1. Endocrinology 2003, 144, 2650–2658. [Google Scholar] [CrossRef]
- Sesti, G.; Tullio, A.N.; D’Alfonso, R.; Napolitano, M.; Marini, M.A.; Borboni, P.; Longhi, R.; Albonici, L.; Fusco, A.; Aglianó, A.M.; et al. Tissue-specific expression of two alternatively spliced isoforms of the human insulin receptor protein. Acta Diabetol. 1994, 31, 59–65. [Google Scholar] [CrossRef]
- Huang, J.; Morehouse, C.; Streicher, K.; Higgs, B.W.; Gao, J.; Czapiga, M.; Boutrin, A.; Zhu, W.; Brohawn, P.; Chang, Y.; et al. Altered expression of insulin receptor isoforms in breast cancer. PLoS ONE 2011, 6, e26177. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhu, W.; Streicher, K.; Morehouse, C.; Brohawn, P.; Ge, X.; Dong, Z.; Yin, X.; Zhu, G.; Gu, Y.; et al. Increased IR-A/IR-B ratio in non-small cell lung cancers associates with lower epithelial-mesenchymal transition signature and longer survival in squamous cell lung carcinoma. BMC Cancer 2014, 14, 131. [Google Scholar] [CrossRef]
- Ziegler, A.; König, I.R.; Schulz-Knappe, P.D.M. Challenges in planning and conducting diagnostic studies with molecular biomarkers. Dtsch. Med. Wochenschr. 2013, 138, e14–e24. [Google Scholar] [CrossRef]
- Hajia, M. Limitations of Different PCR Protocols Used in Diagnostic Laboratories: A Short Review. Med. Lab. J. 2018, 1, 1–6. [Google Scholar] [CrossRef]
- Eggington, J.M.; Bowles, K.R.; Moyes, K.; Manley, S.; Esterling, L.E.; Sizemore, S.; Rosenthal, E.T.; Theisen, A.P.; Saam, J.; Arnell, C.; et al. A comprehensive laboratory-based program for classification of variants of uncertain significance in hereditary cancer genes. Clin. Genet. 2014, 86, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Zangar, R.C.; Daly, D.S.; White, A.M. ELISA microarray technology as a high-throughput system for cancer biomarker validation. Expert Rev. Proteom. 2006, 3, 37–44. [Google Scholar] [CrossRef]
- Horvatovich, P.L.; Bischoff, R. Current Technological Challenges in Biomarker Discovery and Validation. Eur. J. Mass. Spectrom. 2010, 16, 101–121. [Google Scholar] [CrossRef]
- Simon, P. Q-Gene: Processing Quantitative Real-time RT-PCR Data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef]
- Broers, J.L.; Machiels, B.M.; Kuijpers, H.J.; Smedts, F.; van den Kieboom, R.; Raymond, Y.; Ramaekers, F.C. A- and B-type lamins are differentially expressed in normal human tissues. Histochem. Cell Biol. 1997, 107, 505–517. [Google Scholar] [CrossRef]
- Hutchison, C.J.; Worman, H.J. A-type lamins: Guardians of the soma? Nat. Cell Biol. 2004, 6, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, B.; Khandjian, E.; Hirt, A.; Luthy, A.; Weil, R.; Wagner, H.P. Repression of nuclear lamin A and C gene expression in human acute lymphoblastic leukemia and non-Hodgkin’s lymphoma cells. Leuk. Res. 1990, 14, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Worman, H.J. Expression of nuclear lamins in human tissues and cancer cell lines and transcription from the promoters of the lamin A/C and B1 genes. Exp. Cell Res. 1997, 236, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Agrelo, R.; Setien, F.; Espada, J.; Artiga, M.J.; Rodriguez, M.; Perez-Rosado, A.; Sanchez-Aguilera, A.; Fraga, M.F.; Piris, M.A.; Esteller, M. Inactivation of the lamin A/C gene by CpG island promoter hypermethylation in hematologic malignancies, and its association with poor survival in nodal diffuse large B-cell lymphoma. J. Clin. Oncol. 2005, 23, 3940–3947. [Google Scholar] [CrossRef] [PubMed]
- Willis, N.D.; Cox, T.R.; Rahman-Casans, S.F.; Smits, K.; Przyborski, S.A.; van den Brandt, P.; van Engeland, M.; Weijenberg, M.; Wilson, R.G.; de Bruine, A.; et al. Lamin A/C is a risk biomarker in colorectal cancer. PLoS ONE 2008, 3, e2988. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T. Reversal of the cellular phenotype in the premature aging disease Hutchinson-Gilford progeria syndrome. Nat. Med. 2005, 11, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Toth, J.I.; Yang, S.H.; Qiao, X.; Beigneux, A.P.; Gelb, M.H.; Moulson, C.L.; Miner, J.H.; Young, S.G.; Fong, L.G. Blocking protein farnesyltransferase improves nuclear shape in fibroblasts from humans with progeroid syndromes. Proc. Natl. Acad. Sci. USA 2005, 102, 12873–12878. [Google Scholar] [CrossRef] [PubMed]
- Moulson, C.L.; Fong, L.G.; Gardner, J.M.; Farber, E.A.; Go, G.; Passariello, A.; Grange, D.K.; Young, S.G.; Miner, J.H. Increased progerin expression associated with unusual LMNA mutations causes severe progeroid syndromes. Hum. Mutat. 2007, 28, 882–889. [Google Scholar] [CrossRef]
- McClintock, D.; Ratner, D.; Lokuge, M.; Owens, D.M.; Gordon, L.B.; Collins, F.S.; Djabali, K. The mutant form of lamin A that causes Hutchinson-Gilford progeria is a biomarker of cellular aging in human skin. PLoS ONE 2007, 2, e1269. [Google Scholar] [CrossRef]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef] [PubMed]
- Perillie, P.E.; Finch, S.C. The local exudative cellular response in leukemia. J. Clin. Investig. 1960, 39, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Itälä, M.; Vainio, O.; Remes, K. Functional abnormalities in granulocytes predict susceptibility to bacterial infections in chronic lymphocytic leukaemia. Eur. J. Haematol. 1996, 57, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.O.; Chang, X.; Blaser, B.W.; Caligiuri, M.A.; Zheng, P.; Liu, Y. Tumor growth impedes natural-killer-cell maturation in the bone marrow. Blood 2006, 108, 246–252. [Google Scholar] [CrossRef]
- Carlsten, M.; Baumann, B.C.; Simonsson, M.; Jädersten, M.; Forsblom, A.-M.; Hammarstedt, C.; Bryceson, Y.T.; Ljunggren, H.-G.; Hellström-Lindberg, E.; Malmberg, K.-J. Reduced DNAM-1 expression on bone marrow NK cells associated with impaired killing of CD34+ blasts in myelodysplastic syndrome. Leukemia 2010, 24, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Riches, J.C.; Ramsay, A.G.; Gribben, J.G. Immune Reconstitution in Chronic Lymphocytic Leukemia. Curr. Hematol. Malig. Rep. 2012, 7, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Bayona, P.W.; Kim, M.; Chang, J.; Hong, S.; Park, Y.; Budiman, A.; Kim, Y.J.; Choi, C.Y.; Kim, W.S.; et al. Macrophage Lamin A/C Regulates Inflammation and the Development of Obesity-Induced Insulin Resistance. Front. Immunol. 2018, 9, 696. [Google Scholar] [CrossRef] [PubMed]
- Goalstone, M.; Carel, K.; Leitner, J.W.; Draznin, B. Insulin stimulates the phosphorylation and activity of farnesyltransferase via the Ras-mitogen-activated protein kinase pathway. Endocrinology 1997, 138, 5119–5124. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.S.; Fong, L.G.; Yang, S.H.; Coffinier, C.; Young, S.G. The posttranslational processing of prelamin A and disease. Annu. Rev. Genom. Hum. Genet. 2009, 10, 153–174. [Google Scholar] [CrossRef]
- Reagan, J.L.; Ingham, R.R.; Dalia, S.; Furman, M.S.; Merhi, B.; Nemr, S.; Zarrabi, A.; Mitri, J.; Castillo, J.J. Association Between Obesity/Overweight and Leukemia: A Meta-Analysis of Prospective Cohort Studies. Blood 2011, 118, 3588. [Google Scholar] [CrossRef]
- Larsson, S.C.; Wolk, A. Overweight and obesity and incidence of leukemia: A meta-analysis of cohort studies. Int. J. Cancer 2008, 122, 1418–1421. [Google Scholar] [CrossRef] [PubMed]
- Galati, P.C.; Ribeiro, C.M.; Pereira, L.T.G.; Amato, A.A. The association between excess body weight at diagnosis and pediatric leukemia prognosis: A systematic review and meta-analysis. Blood Rev. 2022, 51, 100870. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer. | Sense (5′→3′) | Anti Sense (5′→3′) | Probe | Accession Number |
|---|---|---|---|---|
| LMNA | TGACTGTGGTTGAGGACGAC | GACACTGGAGGCAGAAGAGC | CGCTGAGTACAACCT | NM_170707.3 |
| LMNC | GTGGAAGGCACAGAACACCT | GCGGCGGCTACCACTCAC | AGATGACCTGCTCCATCACC | NM_005572.3 |
| LMNAΔ10 | AACTCCACTGGGGAAGGCTCC | GCTCCTGAGCCGCTGGCAGA | AGTACAACCTGCGCTCGCGC | NM_170708.3 |
| LMNAΔ50 | GCGTCAGGAGCCCTGAGC | GACGCAGGAAGCCTCCAC | AGCATCATGTAATCTGGGACCT | NM_001282626.1 |
| INSR (IR-A) | TATCCGGAACAACCTCACTA | GGAAGAGCAGCAAGTAATCA | CTCTGTCATCGAAGGACACTTG | NM_001079817 |
| INSR (IR-B) | AGGAGTCCTCGTTTAGGAAG | AGGAAGTGTTGGGGAAAG | AGAAAAACCTCTTCAGGCACTG | NM_000208 |
| Ubiquitin C | ACTACAACATCCAGAAAGAGTCCA | CCAGTCAGGGTCTTCACGAAG | CCCACCTCTGAGACGGAGCACCAG | NM_021009.6 |
| RPL13 | AACAAGTTGAAGTACCTGGCTTTC | TGGTTTTGTGGGGCAGCATA | CGCAAGCGGATGAACACCAACCCT | NM_000977.3 |
| Gender | Age (years) | BMI kg/m2 | |
|---|---|---|---|
| Control | 14M, 18F | 31.9 ± 2.18 | 32.1 ± 2.0 |
| AML | 9M, 8F | 48.2 ± 5.18 * | 26.4 ± 1.28 |
| CML | 4M, 1F | 59.4 ± 6.75 *ɛ | 30.4 ± 1.78 |
| ALL | 5M, 3F | 34.4 ± 5.51 | 25.9 ± 2.38 |
| CLL | 7M, 8F | 66.5 ± 3.42 *ɛ | 28.1 ± 1.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshaalan, K.S.; Albawardi, T.K.; Zhra, M.; Bin Sulaiman, N.; Jnied, O.Y.; Saleem, R.A.; Aljada, A. Differential Expression of LMNA/C and Insulin Receptor Transcript Variants in Peripheral Blood Mononuclear Cells of Leukemia Patients. J. Clin. Med. 2024, 13, 2568. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092568
Alshaalan KS, Albawardi TK, Zhra M, Bin Sulaiman N, Jnied OY, Saleem RA, Aljada A. Differential Expression of LMNA/C and Insulin Receptor Transcript Variants in Peripheral Blood Mononuclear Cells of Leukemia Patients. Journal of Clinical Medicine. 2024; 13(9):2568. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092568
Chicago/Turabian StyleAlshaalan, Khalid Saud, Turki Khalid Albawardi, Mahmoud Zhra, Norah Bin Sulaiman, Osama Yaheia Jnied, Rimah Abdullah Saleem, and Ahmad Aljada. 2024. "Differential Expression of LMNA/C and Insulin Receptor Transcript Variants in Peripheral Blood Mononuclear Cells of Leukemia Patients" Journal of Clinical Medicine 13, no. 9: 2568. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092568