
Evaluation of a Combinatorial Immunotherapy Regimen That Can Cure Mice Bearing MYCN-Driven High-Risk Neuroblastoma That Resists Current Clinical Therapy

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Cells
2.2. Murine Tumor Models
2.3. Radiotherapy
2.4. Antibodies and Immunocytokine
2.5. Chemotherapy
2.6. Statistical Analyses
3. Results
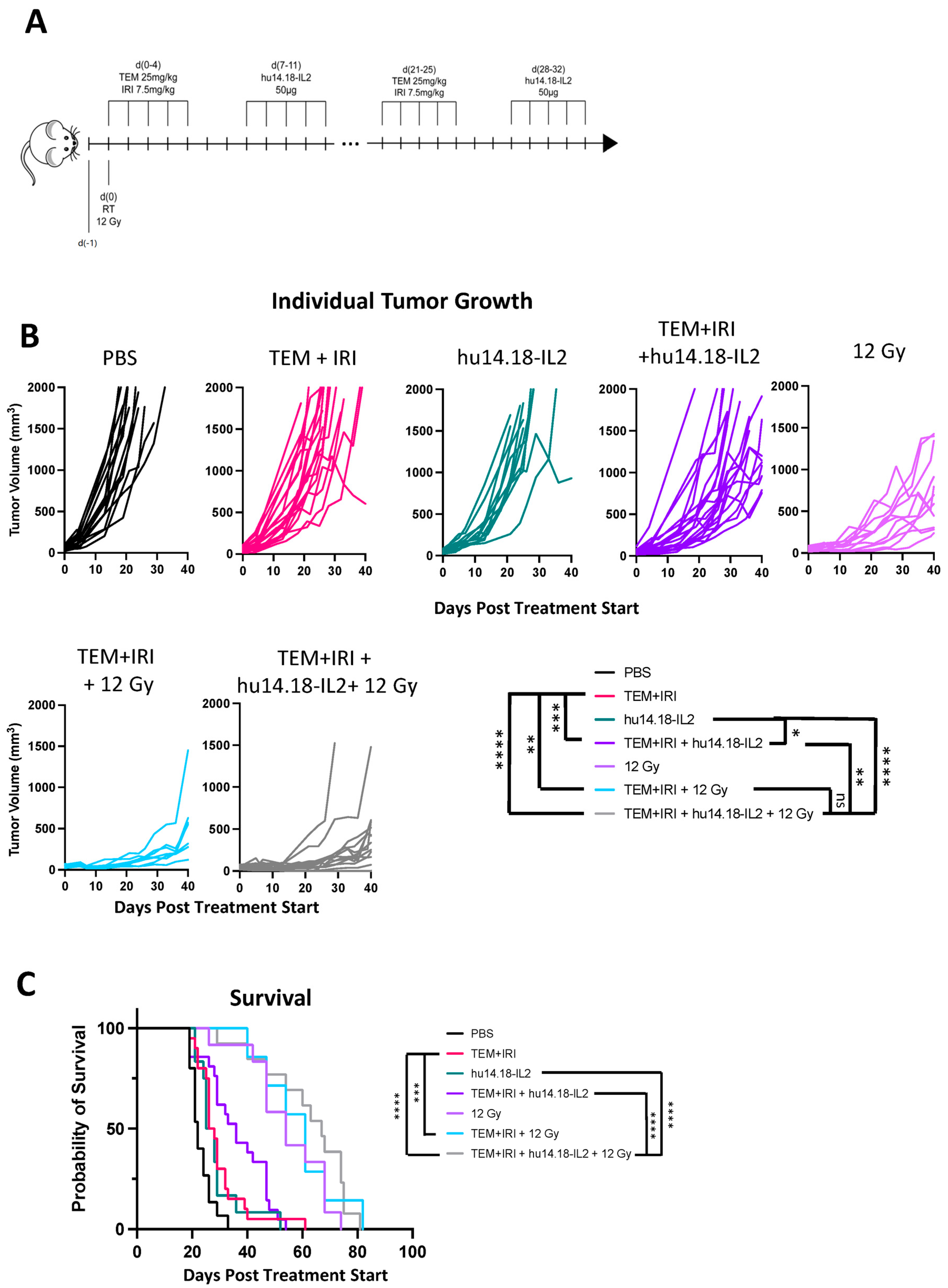
3.1. 9464D-GD2 Does Not Respond to the Combination of Temozolomide + Irinotecan + Anti-GD2
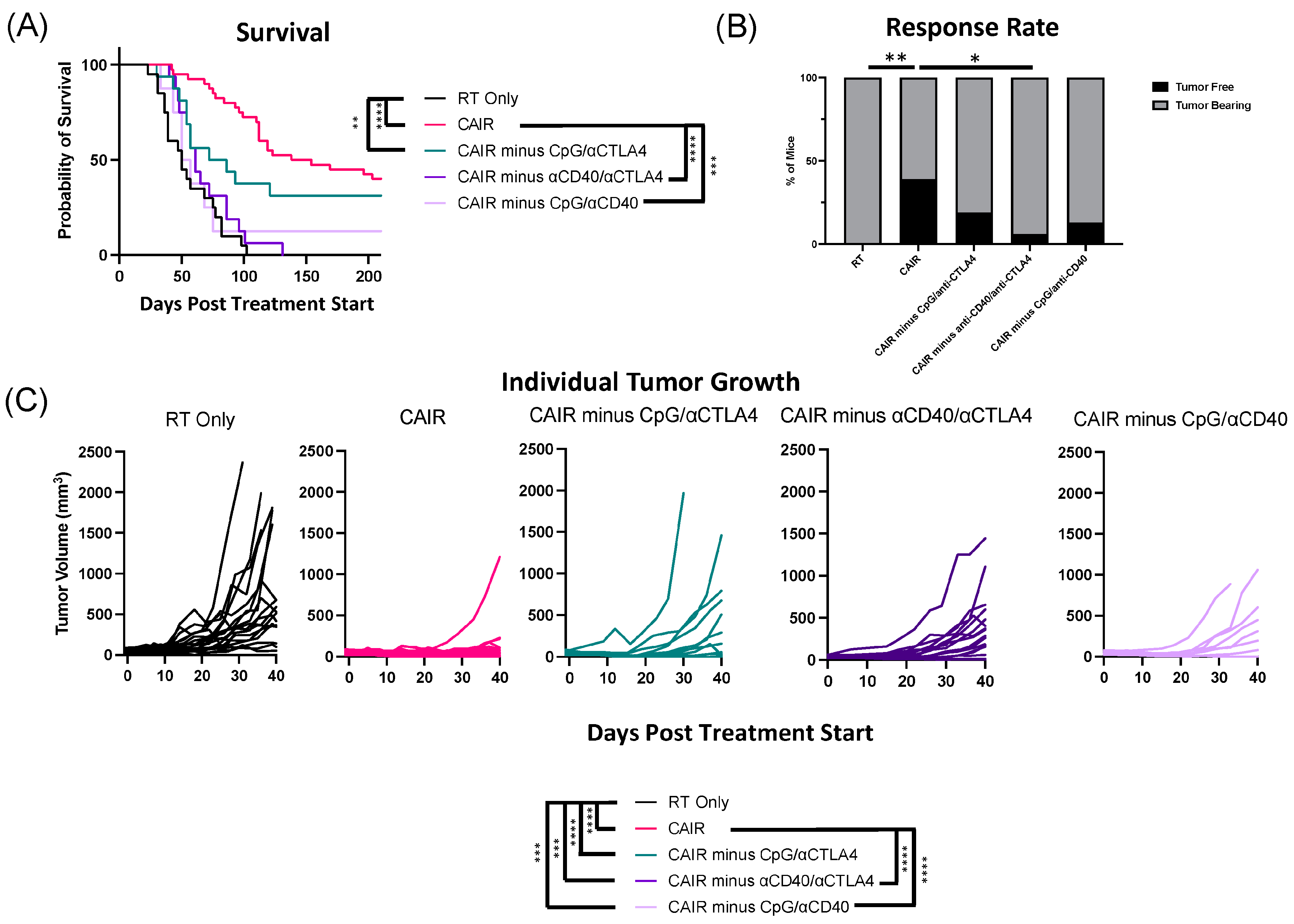
3.2. The Radiotherapy and the IT-IC Components Are Each Essential to Maintaining the Anti-Tumor Potency of CAIR
3.3. The Potency of the CAIR Regimen Is Abrogated by Eliminating CD40 with Either CpG or Anti-CTLA-4
3.4. Efficacy of the CAIR Regimen on a Much Smaller Distant Tumor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; London, W.B.; Kreissman, S.G.; Chen, H.X.; Smith, M.; Anderson, B.; Villablanca, J.G.; Matthay, K.K.; et al. Anti-GD2 antibody with GM-CSF, interleukin-2, and isotretinoin for neuroblastoma. N. Engl. J. Med. 2010, 363, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; Naranjo, A.; Diccianni, M.B.; Gan, J.; Hank, J.A.; Batova, A.; London, W.B.; Tenney, S.C.; et al. Long-Term Follow-up of a Phase III Study of ch14.18 (Dinutuximab) + Cytokine Immunotherapy in Children with High-Risk Neuroblastoma: COG Study ANBL0032. Clin. Cancer Res. 2021, 27, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.V.; Gilman, A.L.; Ozkaynak, M.F.; Naranjo, A.; London, W.B.; Tenney, S.C.; Diccianni, M.; Hank, J.A.; Parisi, M.T.; Shulkin, B.L.; et al. Outcomes Following GD2-Directed Postconsolidation Therapy for Neuroblastoma after Cessation of Random Assignment on ANBL0032: A Report from the Children’s Oncology Group. J. Clin. Oncol. 2022, 40, 4107–4118. [Google Scholar] [CrossRef] [PubMed]
- Ladenstein, R.; Potschger, U.; Valteau-Couanet, D.; Luksch, R.; Castel, V.; Yaniv, I.; Laureys, G.; Brock, P.; Michon, J.M.; Owens, C.; et al. Interleukin 2 with anti-GD2 antibody ch14.18/CHO (dinutuximab beta) in patients with high-risk neuroblastoma (HR-NBL1/SIOPEN): A multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Mody, R.; Yu, A.L.; Naranjo, A.; Zhang, F.F.; London, W.B.; Shulkin, B.L.; Parisi, M.T.; Servaes, S.E.; Diccianni, M.B.; Hank, J.A.; et al. Irinotecan, Temozolomide, and Dinutuximab with GM-CSF in Children with Refractory or Relapsed Neuroblastoma: A Report from the Children’s Oncology Group. J. Clin. Oncol. 2020, 38, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Lode, H.N.; Ladenstein, R.; Troschke-Meurer, S.; Struppe, L.; Siebert, N.; Zumpe, M.; Ehlert, K.; Huber, S.; Glogova, E.; Hundsdoerfer, P.; et al. Effect and Tolerance of N5 and N6 Chemotherapy Cycles in Combination with Dinutuximab Beta in Relapsed High-Risk Neuroblastoma Patients Who Failed at Least One Second-Line Therapy. Cancers 2023, 15, 3364. [Google Scholar] [CrossRef]
- Flaadt, T.; Ladenstein, R.L.; Ebinger, M.; Lode, H.N.; Arnardottir, H.B.; Poetschger, U.; Schwinger, W.; Meisel, R.; Schuster, F.R.; Doring, M.; et al. Anti-GD2 Antibody Dinutuximab Beta and Low-Dose Interleukin 2 after Haploidentical Stem-Cell Transplantation in Patients with Relapsed Neuroblastoma: A Multicenter, Phase I/II Trial. J. Clin. Oncol. 2023, 41, 3135–3148. [Google Scholar] [CrossRef]
- Del Bufalo, F.; De Angelis, B.; Caruana, I.; Del Baldo, G.; De Ioris, M.A.; Serra, A.; Mastronuzzi, A.; Cefalo, M.G.; Pagliara, D.; Amicucci, M.; et al. GD2-CART01 for Relapsed or Refractory High-Risk Neuroblastoma. N. Engl. J. Med. 2023, 388, 1284–1295. [Google Scholar] [CrossRef]
- Yankelevich, M.; Thakur, A.; Modak, S.; Chu, R.; Taub, J.; Martin, A.; Schalk, D.L.; Schienshang, A.; Whitaker, S.; Rea, K.; et al. Targeting GD2-positive Refractory/Resistant Neuroblastoma and Osteosarcoma with Anti-CD3× Anti-GD2 Bispecific Antibody Armed T cells. Res. Sq. 2023. preprint. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Majzner, R.G.; Sondel, P.M. Immunotherapy of Neuroblastoma: Facts and Hopes. Clin. Cancer Res. 2022, 28, 3196–3206. [Google Scholar] [CrossRef]
- Morris, Z.S.; Guy, E.I.; Francis, D.M.; Gressett, M.M.; Werner, L.R.; Carmichael, L.L.; Yang, R.K.; Armstrong, E.A.; Huang, S.; Navid, F.; et al. In Situ Tumor Vaccination by Combining Local Radiation and Tumor-Specific Antibody or Immunocytokine Treatments. Cancer Res. 2016, 76, 3929–3941. [Google Scholar] [CrossRef] [PubMed]
- Morris, Z.S.; Guy, E.I.; Werner, L.R.; Carlson, P.M.; Heinze, C.M.; Kler, J.S.; Busche, S.M.; Jaquish, A.A.; Sriramaneni, R.N.; Carmichael, L.L.; et al. Tumor-Specific Inhibition of In Situ Vaccination by Distant Untreated Tumor Sites. Cancer Immunol. Res. 2018, 6, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Voeller, J.; Erbe, A.K.; Slowinski, J.; Rasmussen, K.; Carlson, P.M.; Hoefges, A.; VandenHeuvel, S.; Stuckwisch, A.; Wang, X.; Gillies, S.D.; et al. Combined innate and adaptive immunotherapy overcomes resistance of immunologically cold syngeneic murine neuroblastoma to checkpoint inhibition. J. Immunother. Cancer 2019, 7, 344. [Google Scholar] [CrossRef] [PubMed]
- Kroesen, M.; Nierkens, S.; Ansems, M.; Wassink, M.; Orentas, R.J.; Boon, L.; den Brok, M.H.; Hoogerbrugge, P.M.; Adema, G.J. A transplantable TH-MYCN transgenic tumor model in C57Bl/6 mice for preclinical immunological studies in neuroblastoma. Int. J. Cancer 2014, 134, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Das, S.; Crespo, A.C.; Cornel, A.M.; Patel, A.G.; Mahadevan, N.R.; Campisi, M.; Ali, A.K.; Sharma, B.; Rowe, J.H.; et al. Mesenchymal and adrenergic cell lineage states in neuroblastoma possess distinct immunogenic phenotypes. Nat. Cancer 2022, 3, 1228–1246. [Google Scholar] [CrossRef]
- Stip, M.C.; Evers, M.; Nederend, M.; Chan, C.; Reiding, K.R.; Damen, M.J.; Heck, A.J.R.; Koustoulidou, S.; Ramakers, R.; Krijger, G.C.; et al. IgA antibody immunotherapy targeting GD2 is effective in preclinical neuroblastoma models. J. Immunother. Cancer 2023, 11, e006948. [Google Scholar] [CrossRef]
- Lucarini, V.; Melaiu, O.; D’Amico, S.; Pastorino, F.; Tempora, P.; Scarsella, M.; Pezzullo, M.; De Ninno, A.; D’Oria, V.; Cilli, M.; et al. Combined mitoxantrone and anti-TGFbeta treatment with PD-1 blockade enhances antitumor immunity by remodelling the tumor immune landscape in neuroblastoma. J. Exp. Clin. Cancer Res. 2022, 41, 326. [Google Scholar] [CrossRef]
- Hoogenboom, M.; Eikelenboom, D.C.; van den Bijgaart, R.J.E.; Heerschap, A.; Wesseling, P.; den Brok, M.H.; Futterer, J.J.; Adema, G.J. Impact of MR-guided boiling histotripsy in distinct murine tumor models. Ultrason. Sonochem. 2017, 38, 1–8. [Google Scholar] [CrossRef]
- Liu, X.; Wills, C.A.; Chen, L.; Zhang, J.; Zhao, Y.; Zhou, M.; Sundstrom, J.M.; Schell, T.; Spiegelman, V.S.; Young, M.M.; et al. Small extracellular vesicles induce resistance to anti-GD2 immunotherapy unveiling tipifarnib as an adjunct to neuroblastoma immunotherapy. J. Immunother. Cancer 2022, 10, e004399. [Google Scholar] [CrossRef]
- Boboila, S.; Okochi, S.; Banerjee, D.; Barton, S.; Street, C.; Zenilman, A.L.; Wang, Q.; Gartrell, R.D.; Saenger, Y.M.; Welch, D.; et al. Combining immunotherapy with high-dose radiation therapy (HDRT) significantly inhibits tumor growth in a syngeneic mouse model of high-risk neuroblastoma. Heliyon 2023, 9, e17399. [Google Scholar] [CrossRef]
- Cui, C.; Barberi, T.; Suresh, R.; Friedman, A.D. Adoptive transfer of immature myeloid cells lacking NF-kappaB p50 (p50-IMC) impedes the growth of MHC-matched high-risk neuroblastoma. Mol. Oncol. 2021, 15, 1783–1796. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Vicente, J.; Willoughby, J.E.; Taylor, M.C.; Booth, S.G.; English, V.L.; Williams, E.L.; Penfold, C.A.; Mockridge, C.I.; Inzhelevskaya, T.; Kim, J.; et al. Fc-null anti-PD-1 monoclonal antibodies deliver optimal checkpoint blockade in diverse immune environments. J. Immunother. Cancer 2022, 10, e003735. [Google Scholar] [CrossRef] [PubMed]
- Zeki, J.; Yavuz, B.; Wood, L.; Shimada, H.; Kaplan, D.L.; Chiu, B. Concurrent application of interferon-gamma and vincristine inhibits tumor growth in an orthotopic neuroblastoma mouse model. Pediatr. Surg. Int. 2023, 39, 241. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.R.; Lanati, S.; Wareham, C.; Easton, A.; Dunn, S.N.; Inzhelevskaya, T.; Sadler, F.M.; James, S.; Ashton-Key, M.; Cragg, M.S.; et al. Immune characterization of pre-clinical murine models of neuroblastoma. Sci. Rep. 2020, 10, 16695. [Google Scholar] [CrossRef] [PubMed]
- Buhtoiarov, I.N.; Sondel, P.M.; Wigginton, J.M.; Buhtoiarova, T.N.; Yanke, E.M.; Mahvi, D.A.; Rakhmilevich, A.L. Anti-tumour synergy of cytotoxic chemotherapy and anti-CD40 plus CpG-ODN immunotherapy through repolarization of tumour-associated macrophages. Immunology 2011, 132, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Aiken, T.J.; Erbe, A.K.; Zebertavage, L.; Komjathy, D.; Feils, A.S.; Rodriguez, M.; Stuckwisch, A.; Gillies, S.D.; Morris, Z.S.; Birstler, J.; et al. Mechanism of effective combination radio-immunotherapy against 9464D-GD2, an immunologically cold murine neuroblastoma. J. Immunother. Cancer 2022, 10, e004834. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.D.; Burkhart, C.A.; Marshall, G.M.; Weiss, W.A.; Haber, M. Expression of N-myc and MRP genes and their relationship to N-myc gene dosage and tumor formation in a murine neuroblastoma model. Med. Pediatr. Oncol. 2000, 35, 585–589. [Google Scholar] [CrossRef]
- Wienke, J.; Visser, L.L.; Kholosy, W.M.; Keller, K.M.; Barisa, M.; Poon, E.; Munnings-Tomes, S.; Himsworth, C.; Calton, E.; Rodriguez, A.; et al. Integrative analysis of neuroblastoma by single-cell RNA sequencing identifies the NECTIN2-TIGIT axis as a target for immunotherapy. Cancer Cell 2024, 42, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.K.; Kalogriopoulos, N.A.; Rakhmilevich, A.L.; Ranheim, E.A.; Seo, S.; Kim, K.; Alderson, K.L.; Gan, J.; Reisfeld, R.A.; Gillies, S.D.; et al. Intratumoral hu14.18-IL-2 (IC) induces local and systemic antitumor effects that involve both activated T and NK cells as well as enhanced IC retention. J. Immunol. 2012, 189, 2656–2664. [Google Scholar] [CrossRef]
- Moreno, V.; Perets, R.; Peretz-Yablonski, T.; Fourneau, N.; Girgis, S.; Guo, Y.; Hellemans, P.; Verona, R.; Pendas, N.; Xia, Q.; et al. A phase 1 study of intravenous mitazalimab, a CD40 agonistic monoclonal antibody, in patients with advanced solid tumors. Investig. New Drugs 2023, 41, 93–104. [Google Scholar] [CrossRef]
- Blake, S.J.; James, J.; Ryan, F.J.; Caparros-Martin, J.; Eden, G.L.; Tee, Y.C.; Salamon, J.R.; Benson, S.C.; Tumes, D.J.; Sribnaia, A.; et al. The immunotoxicity, but not anti-tumor efficacy, of anti-CD40 and anti-CD137 immunotherapies is dependent on the gut microbiota. Cell Rep. Med. 2021, 2, 100464. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Bamford, R.N.; Edmondson, E.F.; Waldmann, T.A. IL15 and Anti-PD-1 Augment the Efficacy of Agonistic Intratumoral Anti-CD40 in a Mouse Model with Multiple TRAMP-C2 Tumors. Clin. Cancer Res. 2022, 28, 2082–2093. [Google Scholar] [CrossRef] [PubMed]
- Pfefferle, M.; Dubach, I.L.; Buzzi, R.M.; Durst, E.; Schulthess-Lutz, N.; Baselgia, L.; Hansen, K.; Imhof, L.; Koernig, S.; Le Roy, D.; et al. Antibody-induced erythrophagocyte reprogramming of Kupffer cells prevents anti-CD40 cancer immunotherapy-associated liver toxicity. J. Immunother. Cancer 2023, 11, e005718. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.K.; Kalogriopoulos, N.A.; Rakhmilevich, A.L.; Ranheim, E.A.; Seo, S.; Kim, K.; Alderson, K.L.; Gan, J.; Reisfeld, R.A.; Gillies, S.D.; et al. Intratumoral treatment of smaller mouse neuroblastoma tumors with a recombinant protein consisting of IL-2 linked to the hu14.18 antibody increases intratumoral CD8+ T and NK cells and improves survival. Cancer Immunol. Immunother. 2013, 62, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Moreno, L.; Weston, R.; Barone, G.; Rubio, A.; Makin, G.; Vaidya, S.; Ng, A.; Castel, V.; Nysom, K.; et al. BEACON-Immuno: Results of the dinutuximab beta (dB) randomization of the BEACON-Neuroblastoma phase 2 trial—A European Innovative Therapies for Children with Cancer (ITCC–International Society of Paediatric Oncology Europe Neuroblastoma Group (SIOPEN) trial. J. Clin. Oncol. 2022, 40 (Suppl. S16), 10002. [Google Scholar] [CrossRef]
- Kroesen, M.; Brok, I.C.; Reijnen, D.; van Hout-Kuijer, M.A.; Zeelenberg, I.S.; Den Brok, M.H.; Hoogerbrugge, P.M.; Adema, G.J. Intra-adrenal murine TH-MYCN neuroblastoma tumors grow more aggressive and exhibit a distinct tumor microenvironment relative to their subcutaneous equivalents. Cancer Immunol. Immunother. 2015, 64, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.W.; Sheard, M.A.; Malvar, J.; Fernandez, G.E.; DeClerck, Y.A.; Blavier, L.; Shimada, H.; Theuer, C.P.; Sposto, R.; Seeger, R.C. Anti-CD105 Antibody Eliminates Tumor Microenvironment Cells and Enhances Anti-GD2 Antibody Immunotherapy of Neuroblastoma with Activated Natural Killer Cells. Clin. Cancer Res. 2019, 25, 4761–4774. [Google Scholar] [CrossRef]
- Otsuka, T.; Nishida, S.; Shibahara, T.; Temizoz, B.; Hamaguchi, M.; Shiroyama, T.; Kimura, K.; Miyake, K.; Hirata, H.; Mizuno, Y.; et al. CpG ODN (K3)-toll-like receptor 9 agonist-induces Th1-type immune response and enhances cytotoxic activity in advanced lung cancer patients: A phase I study. BMC Cancer 2022, 22, 744. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Cano-Mejia, J.; Andricovich, J.; Burga, R.A.; Sweeney, E.E.; Fernandes, R. An Engineered Prussian Blue Nanoparticles-based Nanoimmunotherapy Elicits Robust and Persistent Immunological Memory in a TH-MYCN Neuroblastoma Model. Adv. Nanobiomed. Res. 2021, 1, 2100021. [Google Scholar] [CrossRef]
- Pascual-Pasto, G.; McIntyre, B.; Shraim, R.; Buongervino, S.N.; Erbe, A.K.; Zhelev, D.V.; Sadirova, S.; Giudice, A.M.; Martinez, D.; Garcia-Gerique, L.; et al. GPC2 antibody-drug conjugate reprograms the neuroblastoma immune milieu to enhance macrophage-driven therapies. J. Immunother. Cancer 2022, 10, e004704. [Google Scholar] [CrossRef]
- Williams, E.L.; Dunn, S.N.; James, S.; Johnson, P.W.; Cragg, M.S.; Glennie, M.J.; Gray, J.C. Immunomodulatory monoclonal antibodies combined with peptide vaccination provide potent immunotherapy in an aggressive murine neuroblastoma model. Clin. Cancer Res. 2013, 19, 3545–3555. [Google Scholar] [CrossRef]
- Airoldi, I.; Lualdi, S.; Bruno, S.; Raffaghello, L.; Occhino, M.; Gambini, C.; Pistoia, V.; Corrias, M.V. Expression of costimulatory molecules in human neuroblastoma. Evidence that CD40+ neuroblastoma cells undergo apoptosis following interaction with CD40L. Br. J. Cancer 2003, 88, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Vonderheide, R.H. CD40 Agonist Antibodies in Cancer Immunotherapy. Annu. Rev. Med. 2020, 71, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Padron, L.J.; Maurer, D.M.; O’Hara, M.H.; O’Reilly, E.M.; Wolff, R.A.; Wainberg, Z.A.; Ko, A.H.; Fisher, G.; Rahma, O.; Lyman, J.P.; et al. Sotigalimab and/or nivolumab with chemotherapy in first-line metastatic pancreatic cancer: Clinical and immunologic analyses from the randomized phase 2 PRINCE trial. Nat. Med. 2022, 28, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Andersson, H.; Sobti, A.; Jimenez, D.G.; de Coana, Y.P.; Ambarkhane, S.V.; Hagerbrand, K.; Smith, K.E.; Lindstedt, M.; Ellmark, P. Early Pharmacodynamic Changes Measured Using RNA Sequencing of Peripheral Blood from Patients in a Phase I Study with Mitazalimab, a Potent CD40 Agonistic Monoclonal Antibody. Cells 2023, 12, 2365. [Google Scholar] [CrossRef]
- Coveler, A.L.; Smith, D.C.; Phillips, T.; Curti, B.D.; Goel, S.; Mehta, A.N.; Kuzel, T.M.; Markovic, S.N.; Rixe, O.; Bajor, D.L.; et al. Phase 1 dose-escalation study of SEA-CD40: A non-fucosylated CD40 agonist, in advanced solid tumors and lymphomas. J. Immunother. Cancer 2023, 11, e005584. [Google Scholar] [CrossRef]
- Weiss, S.A.; Djureinovic, D.; Jessel, S.; Krykbaeva, I.; Zhang, L.; Jilaveanu, L.; Ralabate, A.; Johnson, B.; Levit, N.S.; Anderson, G.; et al. A Phase I Study of APX005M and Cabiralizumab with or without Nivolumab in Patients with Melanoma, Kidney Cancer, or Non-Small Cell Lung Cancer Resistant to Anti-PD-1/PD-L1. Clin. Cancer Res. 2021, 27, 4757–4767. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PBS | TEM+IRI | hu14.18-IL2 | TEM+IRI + hu14.18-IL2 | 12 Gy | TEM+IRI + 12 Gy | TEM+IRI + hu14.18-IL2 + 12 Gy | |
|---|---|---|---|---|---|---|---|
| PBS | 0.0046 | 0.0303 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| TEM+IRI | 0.5347 | 0.0536 | <0.0001 | 0.0002 | <0.0001 | ||
| hu14.18-IL2 | 0.0603 | <0.0001 | 0.0002 | <0.0001 | |||
| TEM+IRI + hu14.18-IL2 | 0.0003 | 0.0006 | <0.0001 | ||||
| 12 Gy | 0.5663 | 0.1003 | |||||
| TEM+IRI + 12 Gy | 0.8681 | ||||||
| TEM+IRI + hu14.18-IL2 + 12 Gy |
| Treatment | Independent Experiments | Cure Rates |
|---|---|---|
| CAIR | 4 | 15/38 = 39% |
| CAIR minus hu14.18-IL2 | 2 | 0/16 = 0% |
| CAIR minus RT | 2 | 1/16 = 6% |
| CAIR minus CpG | 2 | 7/16 = 44% |
| CAIR minus αCD40 | 2 | 8/16 = 50% |
| CAIR minus αCTLA4 | 3 | 9/21 = 43% |
| CAIR minus CpG/αCTLA4 | 2 | 3/16 = 19% |
| CAIR minus αCD40/αCTLA4 | 2 | 1/16 = 6% |
| CAIR minus CpG/αCD40 | 1 | 1/8 = 13% |
| RT | 3 | 0/19 = 0% |
| RT Only | CAIR | CAIR Minus CpG | CAIR Minus αCD40 | CAIR Minus αCTLA4 | CAIR Minus RT | CAIR Minus hu14.18-IL2 | CAIR Minus CpG/αCTLA4 | CAIR Minus αCD40/αCTLA4 | CAIR Minus CpG/αCD40 | |
|---|---|---|---|---|---|---|---|---|---|---|
| RT Only | <0.0001 | 0.0002 | <0.0001 | <0.0001 | 0.2563 | 0.0621 | 0.0046 | 0.1730 | 0.5602 | |
| CAIR | 0.7724 | 0.7204 | 0.8719 | <0.0001 | <0.0001 | 0.1254 | <0.0001 | 0.0003 | ||
| CAIR minus CpG | 0.6140 | 0.7443 | 0.0033 | 0.0036 | 0.3285 | 0.0017 | 0.0125 | |||
| CAIR minus αCD40 | 0.7684 | 0.0007 | 0.0002 | 0.1898 | 0.0002 | 0.0070 | ||||
| CAIR minus αCTLA4 | 0.0008 | <0.0001 | 0.1939 | <0.0001 | 0.0020 | |||||
| CAIR minus RT | 0.9635 | 0.0422 | 0.8181 | 0.8105 | ||||||
| CAIR minus hu14.18-IL2 | 0.0586 | 0.8580 | 0.6308 | |||||||
| CAIR minus CpG/αCTLA4 | 0.0653 | 0.1639 | ||||||||
| CAIR minus αCD40/αCTLA4 | 0.8717 | |||||||||
| CAIR minus CpG/αCD40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zebertavage, L.; Schopf, A.; Nielsen, M.; Matthews, J.; Erbe, A.K.; Aiken, T.J.; Katz, S.; Sun, C.; Witt, C.M.; Rakhmilevich, A.L.; et al. Evaluation of a Combinatorial Immunotherapy Regimen That Can Cure Mice Bearing MYCN-Driven High-Risk Neuroblastoma That Resists Current Clinical Therapy. J. Clin. Med. 2024, 13, 2561. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092561
Zebertavage L, Schopf A, Nielsen M, Matthews J, Erbe AK, Aiken TJ, Katz S, Sun C, Witt CM, Rakhmilevich AL, et al. Evaluation of a Combinatorial Immunotherapy Regimen That Can Cure Mice Bearing MYCN-Driven High-Risk Neuroblastoma That Resists Current Clinical Therapy. Journal of Clinical Medicine. 2024; 13(9):2561. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092561
Chicago/Turabian StyleZebertavage, Lauren, Allison Schopf, Megan Nielsen, Joel Matthews, Amy K. Erbe, Taylor J. Aiken, Sydney Katz, Claire Sun, Cole M. Witt, Alexander L. Rakhmilevich, and et al. 2024. "Evaluation of a Combinatorial Immunotherapy Regimen That Can Cure Mice Bearing MYCN-Driven High-Risk Neuroblastoma That Resists Current Clinical Therapy" Journal of Clinical Medicine 13, no. 9: 2561. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm13092561