
Immunogenicity and Protective Efficacy of Psoralen-Inactivated SARS-CoV-2 Vaccine in Nonhuman Primates
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of SARS-CoV-2 PsIV
2.3. Purification and Characterization of the SARS-CoV-2 PsIV
2.4. SARS-CoV-2 DNA Vaccine
2.5. Evaluation of Immunogenicity in Nonhuman Primates
2.6. SARS-CoV-2 Microneutralization Assay
2.7. ELISA
2.8. Memory B Cell Assay
2.9. T Cell Recall Assay
2.10. Quantitative ELISA for Domain-Specific IgG
2.11. Digital Droplet RT-PCR
2.12. Data Analysis
3. Results
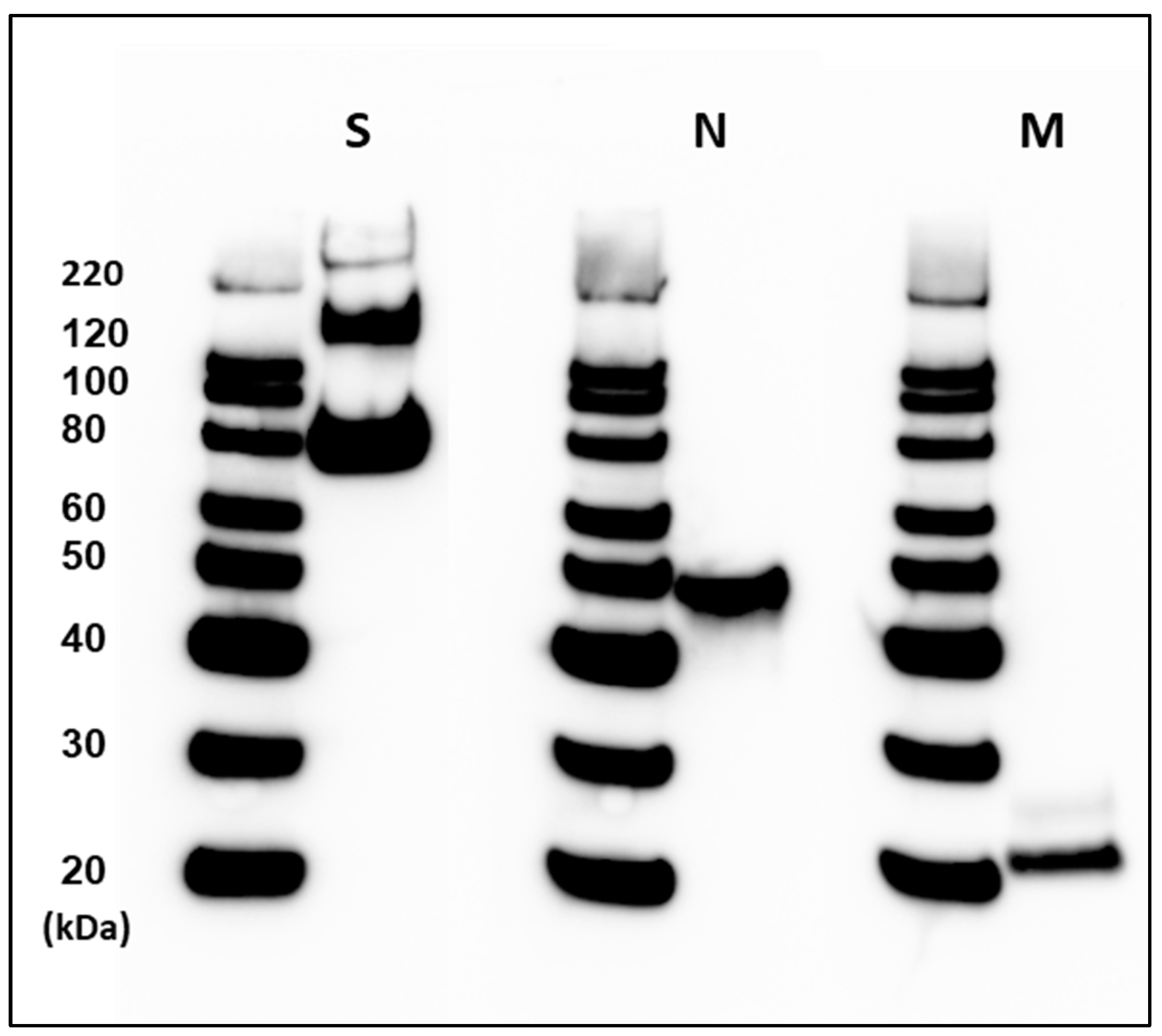
3.1. Preparation and Purification of the SARS-CoV-2 PsIV
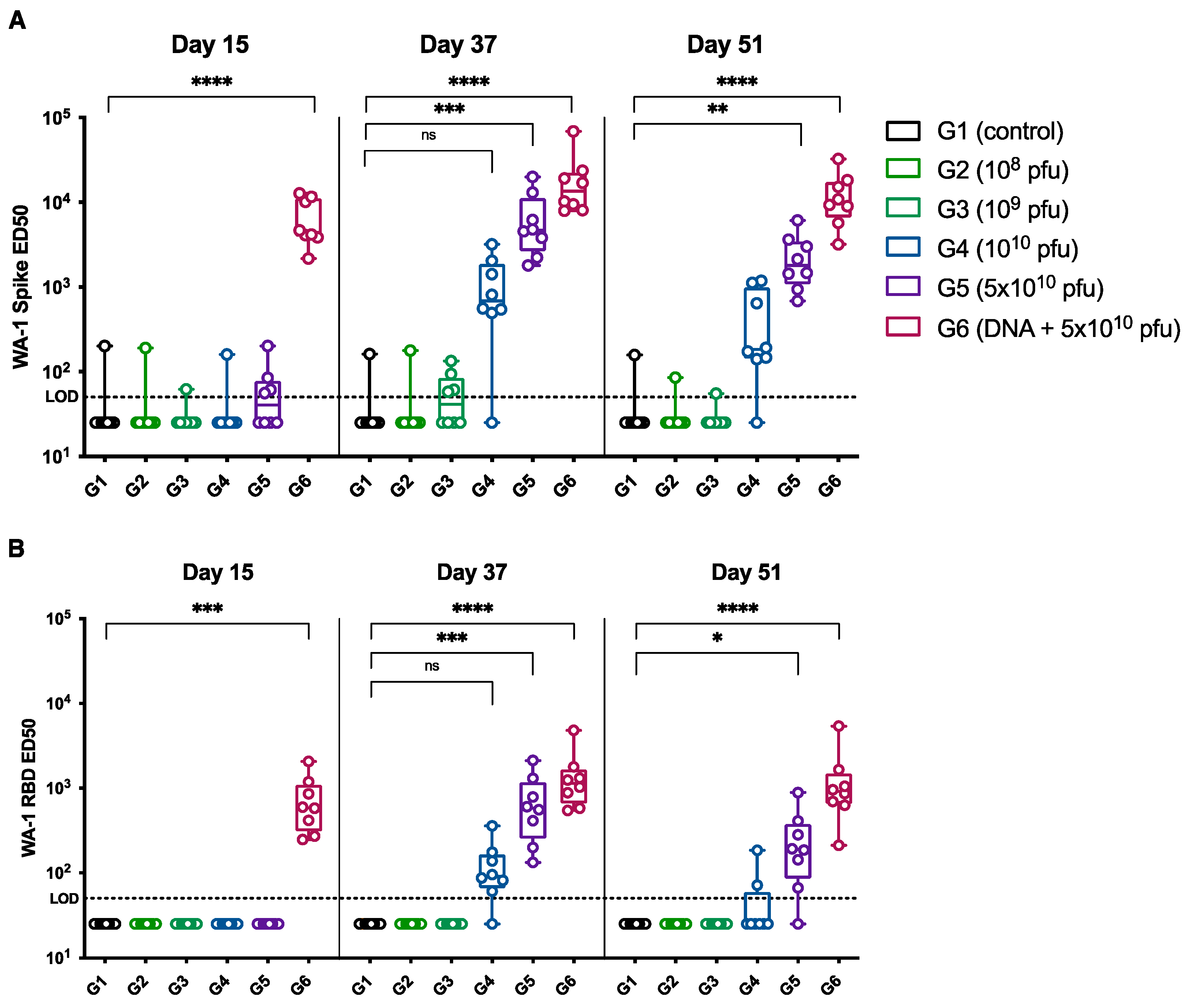
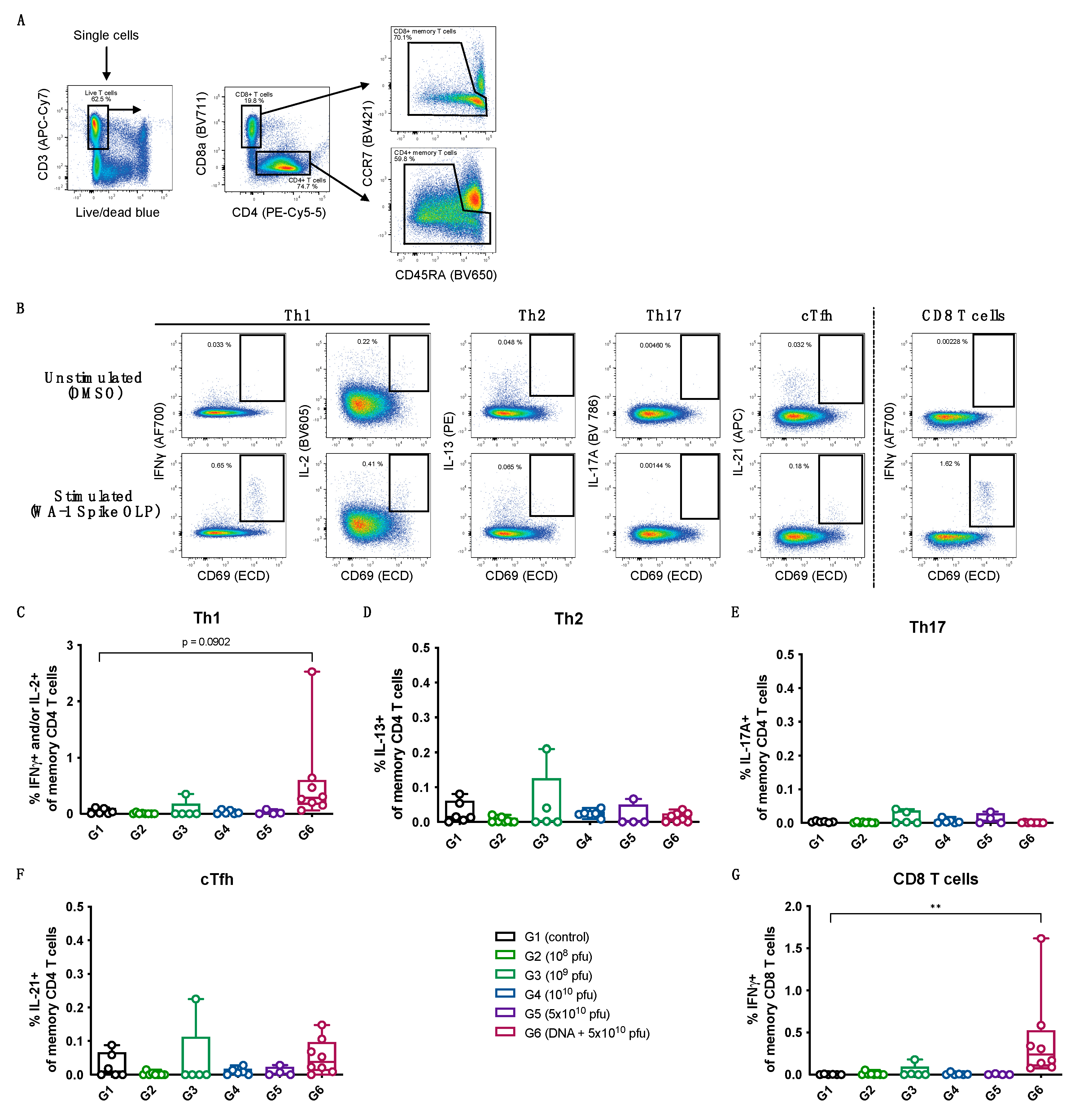
3.2. Immunogenicity of the SARS-CoV-2 PsIV in Nonhuman Primates
3.3. Protection against Infection by the SARS-CoV-2 Delta Variant
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19); StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, A.K.; Ewing, D.; Liang, Z.; Jani, V.; Cheng, Y.; Sun, P.; Raviprakash, K.; Wu, S.J.; Petrovsky, N.; Defang, G.; et al. Immunogenicity of Adjuvanted Psoralen-Inactivated SARS-CoV-2 Vaccines and SARS-CoV-2 Spike Protein DNA Vaccines in BALB/c Mice. Pathogens 2021, 10, 626. [Google Scholar] [CrossRef]
- WHO COVID-19 Dashboard. World Health Organization: Geneva, Switzerland. Available online: https://covid19.who.int/ (accessed on 8 June 2023).
- Harker, S.; James, S.E.; Murphy, J.; Davies, B.; Moore, C.; Tennant, B.P.; Geen, J.; Thomas, D. Serosurveillance of SARS-CoV-2 in Welsh Blood Donors: Establishment of the surveillance system and results up to November 2022. Eurosurveillance 2023, 28, 2200473. [Google Scholar] [CrossRef]
- Marta, R.A.; Nakamura, G.E.K.; de Matos Aquino, B.; Bignardi, P.R. COVID-19 vaccines: Update of the vaccines in use and under development. Vacunas 2022, 23, S88–S102. [Google Scholar] [CrossRef]
- Hadj, H.I. COVID-19 vaccines and variants of concern: A review. Rev. Med. Virol. 2022, 32, e2313. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R.A. Evolution of the SARS-CoV-2 omicron variants BA.1 to BA.5: Implications for immune escape and transmission. Rev. Med. Virol. 2022, 32, e2381. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Guthmiller, J.J.; Stovicek, O.; Wang, J.; Changrob, S.; Li, L.; Halfmann, P.; Zheng, N.Y.; Utset, H.; Stamper, C.T.; Dugan, H.L.; et al. SARS-CoV-2 Infection Severity Is Linked to Superior Humoral Immunity against the Spike. mBio 2021, 12, e02940-20. [Google Scholar] [CrossRef] [PubMed]
- Hartley, G.E.; Edwards, E.S.J.; Aui, P.M.; Varese, N.; Stojanovic, S.; McMahon, J.; Peleg, A.Y.; Boo, I.; Drummer, H.E.; Hogarth, P.M.; et al. Rapid generation of durable B cell memory to SARS-CoV-2 spike and nucleocapsid proteins in COVID-19 and convalescence. Sci. Immunol. 2020, 5, eabf8891. [Google Scholar] [CrossRef]
- Dugan, H.L.; Stamper, C.T.; Li, L.; Changrob, S.; Asby, N.W.; Halfmann, P.J.; Zheng, N.Y.; Huang, M.; Shaw, D.G.; Cobb, M.S.; et al. Profiling B cell immunodominance after SARS-CoV-2 infection reveals antibody evolution to non-neutralizing viral targets. Immunity 2021, 54, 1290–1303.e7. [Google Scholar] [CrossRef]
- Galhaut, M.; Lundberg, U.; Marlin, R.; Schlegl, R.; Seidel, S.; Bartuschka, U.; Heindl-Wruss, J.; Relouzat, F.; Langlois, S.; Dereuddre-Bosquet, N.; et al. Immunogenicity and efficacy of VLA2001 vaccine against SARS-CoV-2 infection in male cynomolgus macaques. Commun. Med. 2024, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721 e719. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Wollowitz, S. Fundamentals of the psoralen-based Helinx technology for inactivation of infectious pathogens and leukocytes in platelets and plasma. Semin. Hematol. 2001, 38 (Suppl. S11), 4–11. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.V.; Riggs, J.L.; Lennette, E.H. Photochemical inactivation of DNA and RNA viruses by psoralen derivatives. J. Gen. Virol. 1978, 40, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.V. Photochemical inactivation of viruses with psoralens: An overview. Blood Cells 1992, 18, 7–25. [Google Scholar] [PubMed]
- Sundaram, A.K.; Ewing, D.; Blevins, M.; Liang, Z.; Sink, S.; Lassan, J.; Raviprakash, K.; Defang, G.; Williams, M.; Porter, K.R.; et al. Comparison of purified psoralen-inactivated and formalin-inactivated dengue vaccines in mice and nonhuman primates. Vaccine 2020, 38, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Lenart, K.; Hellgren, F.; Ols, S.; Yan, X.; Cagigi, A.; Cerveira, R.A.; Winge, I.; Hanczak, J.; Mueller, S.O.; Jasny, E.; et al. A third dose of the unmodified COVID-19 mRNA vaccine CVnCoV enhances quality and quantity of immune responses. Mol. Ther. Methods Clin. Dev. 2022, 27, 309–323. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Kanagal-Shamanna, R. Digital PCR: Principles and Applications. Methods Mol. Biol. 2016, 1392, 43–50. [Google Scholar]
- Vasudevan, H.N.; Xu, P.; Servellita, V.; Miller, S.; Liu, L.; Gopez, A.; Chiu, C.Y.; Abate, A.R. Digital droplet PCR accurately quantifies SARS-CoV-2 viral load from crude lysate without nucleic acid purification. Sci. Rep. 2021, 11, 780. [Google Scholar] [CrossRef] [PubMed]
- Rando, H.M.; Lordan, R.; Lee, A.J.; Naik, A.; Wellhausen, N.; Sell, E.; Kolla, L.; Consortium, C.-R.; Gitter, A.; Greene, C.S. Application of Traditional Vaccine Development Strategies to SARS-CoV-2. mSystems 2023, 8, e0092722. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.L.; Sajkov, D.; Honda-Okubo, Y.; Wilks, S.H.; Aban, M.; Barr, I.G.; Petrovsky, N. Human Phase 1 trial of low-dose inactivated seasonal influenza vaccine formulated with Advax delta inulin adjuvant. Vaccine 2016, 34, 3780–3786. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Gagne, M.; Wagner, D.A.; O’Connell, S.; Narpala, S.R.; Flebbe, D.R.; Andrew, S.F.; Davis, R.L.; Flynn, B.; Johnston, T.S.; et al. Protection against SARS-CoV-2 Beta variant in mRNA-1273 vaccine-boosted nonhuman primates. Science 2021, 374, 1343–1353. [Google Scholar] [CrossRef]
- Wolz, O.O.; Kays, S.K.; Junker, H.; Koch, S.D.; Mann, P.; Quintini, G.; von Eisenhart-Rothe, P.; Oostvogels, L. A Third Dose of the COVID-19 Vaccine, CVnCoV, Increased the Neutralizing Activity against the SARS-CoV-2 Wild-Type and Delta Variant. Vaccines 2022, 10, 508. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Vaccine Formulation | SARS-CoV-2 PsIV Dose (Expressed as Spike Protein Equivalent) Given |
|---|---|---|
| Group 1 | Advax-CpG + PBS (Controls) (Days 0 and 30) | not applicable |
| Group 2 | SARS-CoV-2 PsIV + Advax-CpG (Days 0 and 30) | 0.0075 µg of spike protein/dose |
| Group 3 | SARS-CoV-2 PsIV + Advax-CpG (Days 0 and 30) | 0.075 µg of spike protein/dose |
| Group 4 | SARS-CoV-2 PsIV + Advax-CpG days 0 and 30) | 0.750 µg of spike protein/dose |
| Group 5 | SARS-CoV-2 PsIV + Advax-CpG (Days 0 and 30) | 3.75 µg of spike protein/dose |
| Group 6 | Plasmid DNA (Days −30 and 0) SARS-CoV-2 PsIV + Advax-CpG (Day 30) | 5 mg DNA/dose and 3.75 µg of spike protein/dose |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanders, J.W.; Ewing, D.; Sundaram, A.K.; Gamble, C.S.; Blevins, M.; Liang, Z.; Sanders, L.A.; Ornelles, D.A.; Sun, P.; Lenart, K.; et al. Immunogenicity and Protective Efficacy of Psoralen-Inactivated SARS-CoV-2 Vaccine in Nonhuman Primates. Vaccines 2024, 12, 451. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12050451
Sanders JW, Ewing D, Sundaram AK, Gamble CS, Blevins M, Liang Z, Sanders LA, Ornelles DA, Sun P, Lenart K, et al. Immunogenicity and Protective Efficacy of Psoralen-Inactivated SARS-CoV-2 Vaccine in Nonhuman Primates. Vaccines. 2024; 12(5):451. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12050451
Chicago/Turabian StyleSanders, John W., Daniel Ewing, Appavu K. Sundaram, Christopher Scott Gamble, Maria Blevins, Zhaodong Liang, Leigh Ann Sanders, David A. Ornelles, Peifang Sun, Klara Lenart, and et al. 2024. "Immunogenicity and Protective Efficacy of Psoralen-Inactivated SARS-CoV-2 Vaccine in Nonhuman Primates" Vaccines 12, no. 5: 451. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines12050451