
Oral Supplementation of Ozonated Sunflower Oil Augments Plasma Antioxidant and Anti-Inflammatory Abilities with Enhancement of High-Density Lipoproteins Functionality in Rats


Abstract
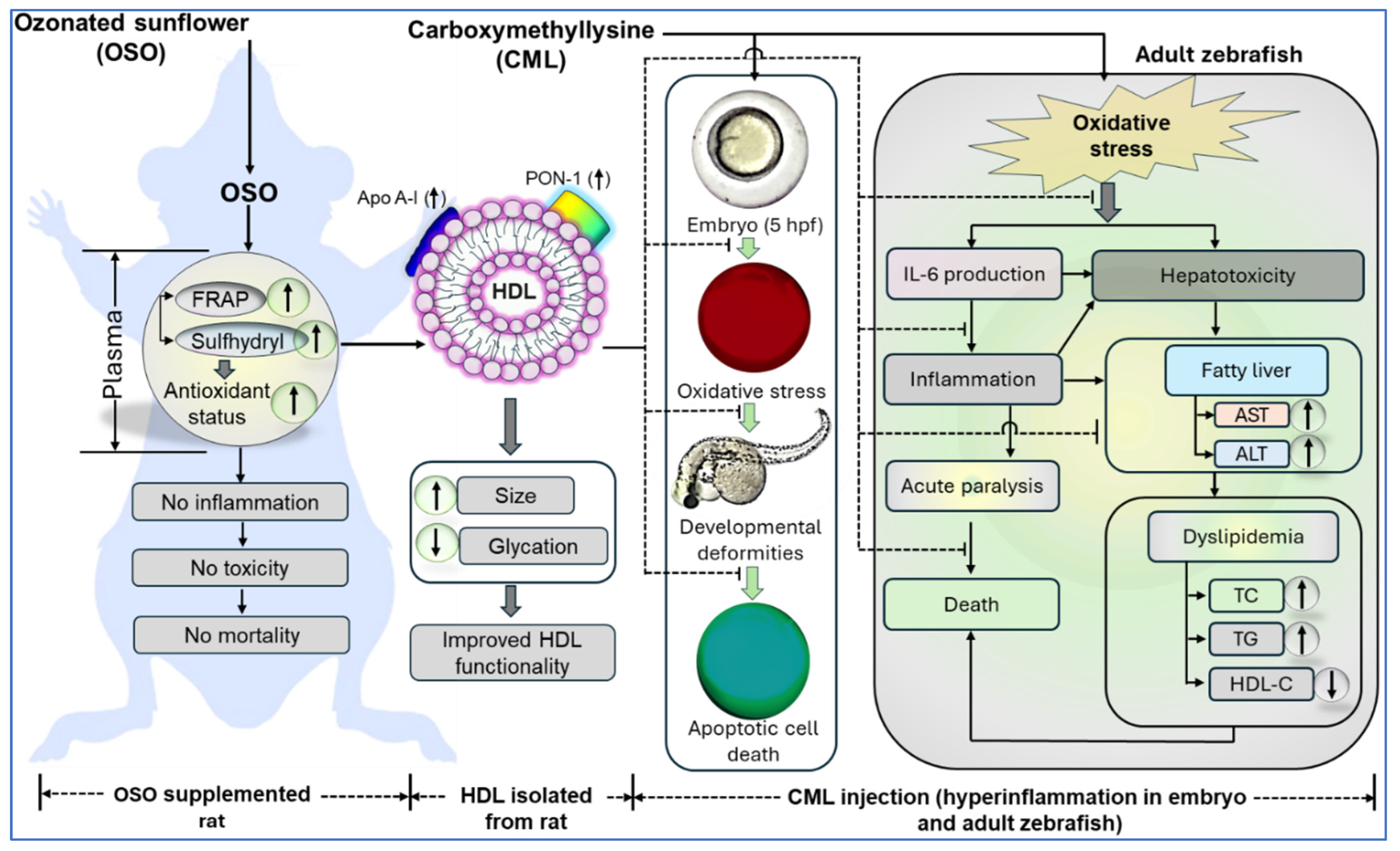
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Rat Maintenance, Experimental Design, and Hematological and Biochemical Analysis
2.3. Ferric Ion Reduction Ability and Sulfhydryl Group Estimation
2.4. Isolation of Lipoproteins and Protein Quantification
2.5. Evaluation of Glycation and Oxidation Extent
2.6. Morphological Analysis of LDL and HDL
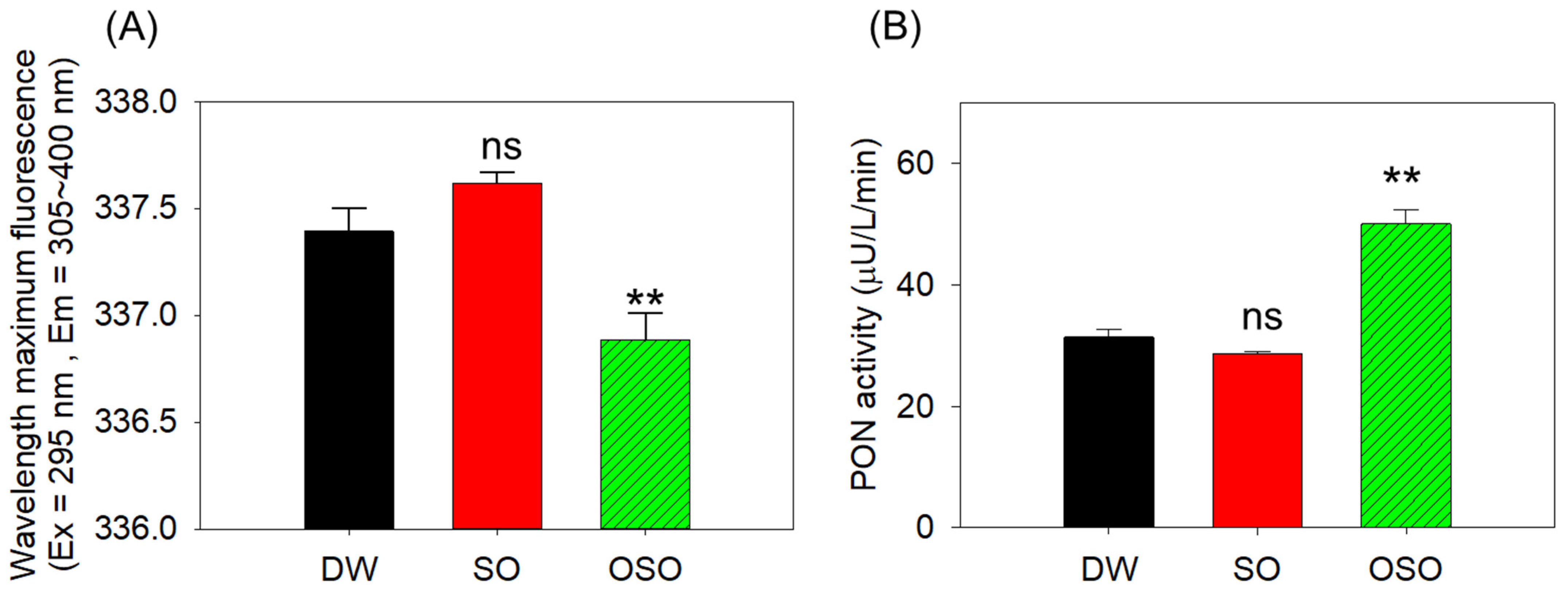
2.7. Paraoxonase (PON)-1 Assay
2.8. Zebrafish Husbandry and Embryo Collection
2.9. Effect of Rat Plasma on Survivability of Zebrafish Embryos
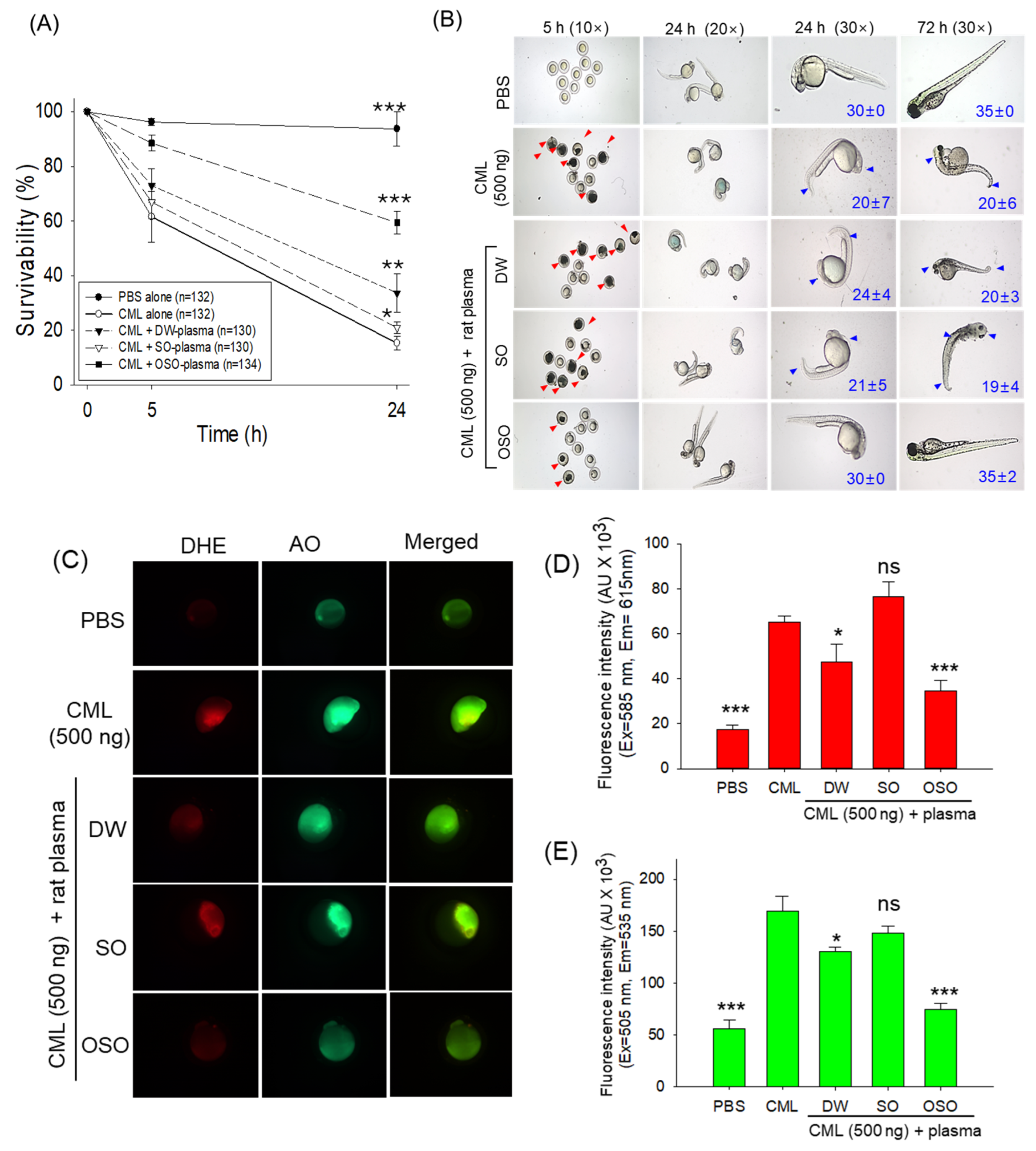
2.10. Determination of Reactive Oxygen Species (ROS) and Apoptosis in Zebrafish Embryos
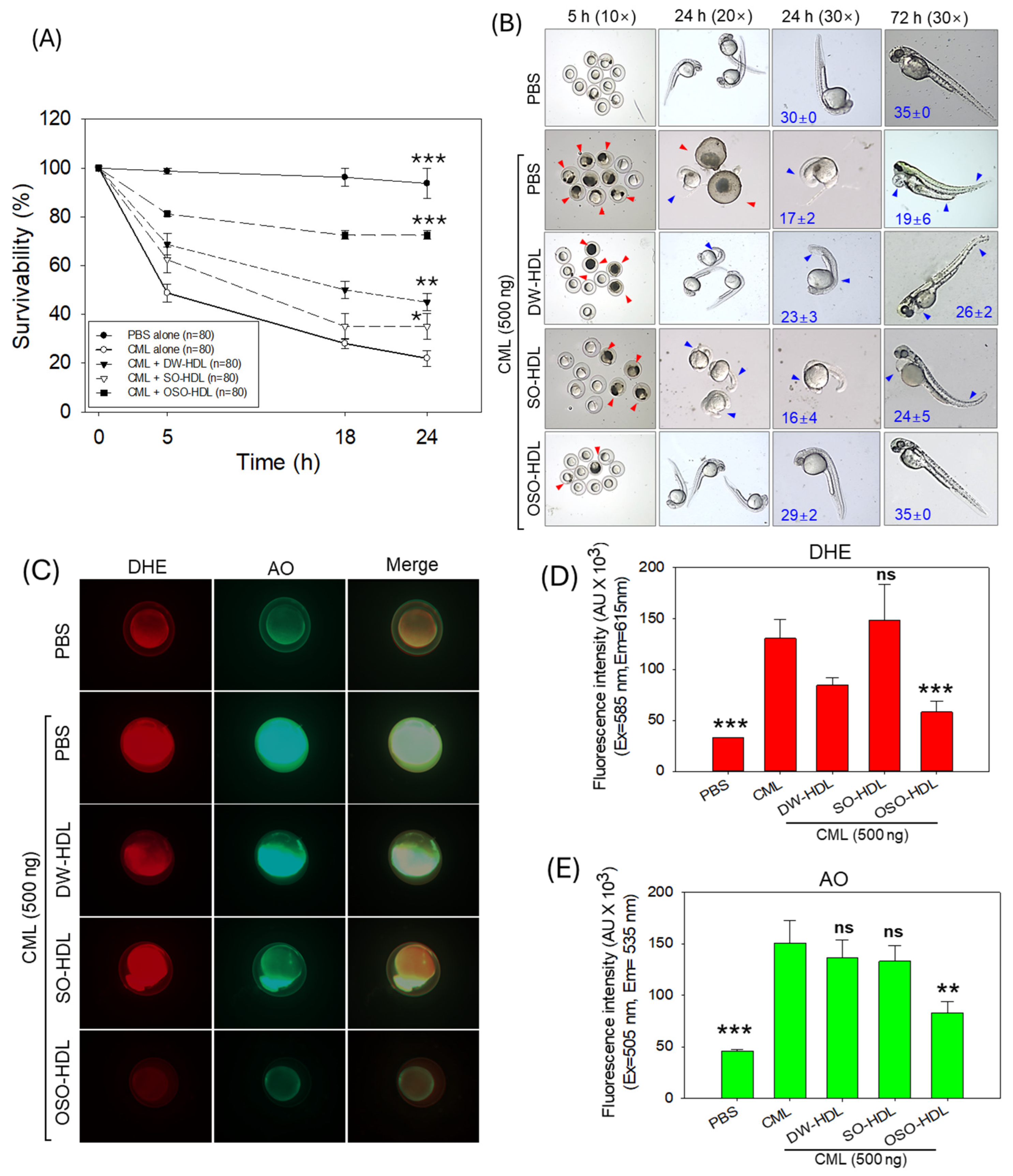
2.11. Effect of Rat HDL on Survivability of Zebrafish Embryos
2.12. Effect of Rat HDL on Carboxymethyllysine Induced Toxicity in Adult Zebrafish
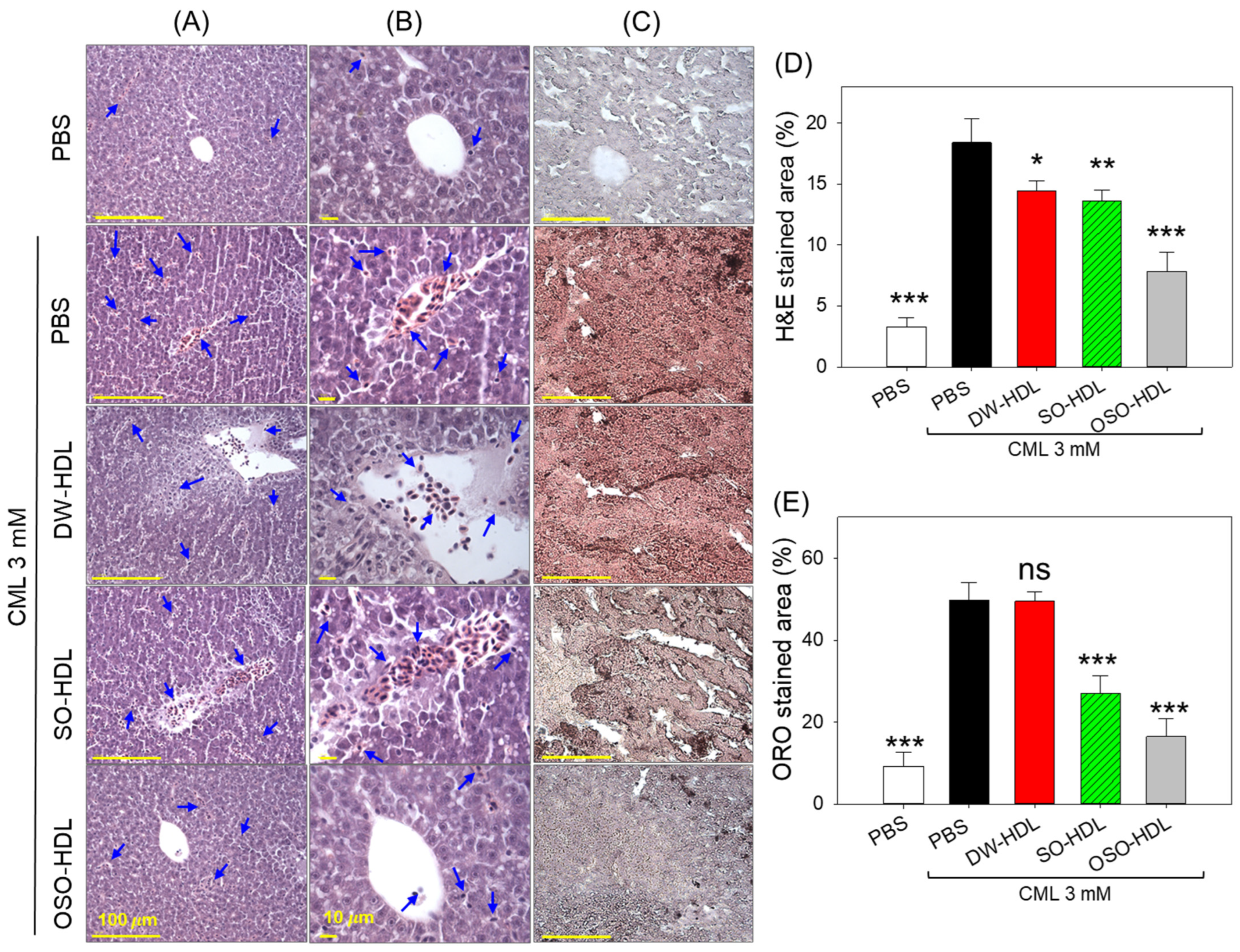
2.13. Liver Histology and Immunohistochemistry (IHC)
2.14. Blood Lipid Profile and Hepatic Function Biomarker Analysis
2.15. Statistical Analysis
3. Results
3.1. OSO Supplementation Posed Nontoxic Effects in Rats
3.2. OSO Supplementation Improved the Antioxidant Status
3.3. Plasma from OSO Protects Zebrafish Embryos against CML-Toxicity
3.4. Impact of OSO on the Attribute of LDL and HDL
3.5. Transmission Electron Microscopic (TEM) Imaging of LDL and HDL
3.6. Enhanced apoA-I Stability and Paraoxonase (PON)-1 Activity in HDL in OSO Group
3.7. HDL from OSO-Fed Rats Attenuated CML-Induced Embryo Toxicity
3.8. Effect of HDL from OSO Fed Rats on CML-Induced Acute Paralysis and Mortality of Adult Zebrafish
3.9. OSO from Rats Prevents CML-Induced Hepatic Damage
3.10. Effect of OSO on CML-Induced Hepatic Inflammation, ROS and Apoptosis
3.11. Hepatic Function Biomarkers
3.12. Influence of OSO-HDL on Dyslipidemia Induced CML
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clavo, B.; Rodríguez-Esparragón, F.; Rodríguez-Abreu, D.; Martínez-Sánchez, G.; Llontop, P.; Aguiar-Bujanda, D.; Fernández-Pérez, L.; Santana-Rodríguez, N. Modulation of oxidative stress by ozone therapy in the prevention and treatment of chemotherapy-induced toxicity: Review and prospects. Antioxidants 2019, 8, 588. [Google Scholar] [CrossRef]
- Galie, M.; Covi, V.; Tabaracci, G.; Malatesta, M. The role of Nrf2 in the antioxidant cellular response to medical ozone exposure. Int. J. Mol. Sci. 2019, 20, 4009. [Google Scholar] [CrossRef] [PubMed]
- Re, L.; Martínez-Sánchez, G.; Bordicchia, M.; Malcangi, G.; Pocognoli, A.; Angel Morales-Segura, M.; Rothchild, J.; Rojas, A. Is ozone pre-conditioning effect linked to Nrf2/EpRE activation pathway in vivo? A preliminary result. Eur. J. Pharmacol. 2014, 742, 158–162. [Google Scholar] [CrossRef]
- Al Faraj, I. Ozonetherapy in hypertension and ischemic diseases. In MedEspera: The 9th International Medical Congress for Students and Young Doctors, 12–14 May 2022, Chisinau, Republic of Moldova; Nicolae Testemitanu State University of Medicine and Pharmacy of the Republic of Moldova, Association of Medical Students and Residents: Chisinau, Republic of Moldova, 2022. [Google Scholar]
- Hernández, F.; Hernández, D.; Zamora, Z.; Díaz, M.; Ancheta, O.; Rodriguez, S.; Torres, D. Giardia duodenalis: Effects of an ozonized sunflower oil product (Oleozon®) on in vitro trophozoites. Exp. Parasitol. 2009, 121, 208–212. [Google Scholar] [CrossRef]
- Zeng, J.; Tang, Z.; Zhang, Y.; Tong, X.; Dou, J.; Gao, L.; Ding, S.; Lu, J. Ozonated autohemotherapy elevates PPAR-γ expression in CD4+ T cells and serum HDL-C levels, a potential immunomodulatory mechanism for treatment of psoriasis. Am. J. Transl. Res. 2021, 13, 349–359. [Google Scholar] [PubMed]
- Ugazio, E.; Tullio, V.; Binello, A.; Tagliapietra, S.; Dosio, F. Ozonated oils as antimicrobial systems in topical applications. their characterization, current applications, and advances in improved delivery techniques. Molecules 2020, 25, 334. [Google Scholar] [CrossRef]
- Díaz, M.F.; Sánchez, Y.; Gómez, M.; Hernández, F.; Da, C.; Veloso, M.C.; De, P.; Pereira, P.A.; Mangrich, A.S.; De Andrade, J.B. Physicochemical characteristics of ozonated sunflower oils obtained by different procedures. Grasas Aceites 2012, 63, 466–474. [Google Scholar] [CrossRef]
- Moureu, S.; Violleau, F.; Haimoud-Lekhal, D.A.; Calmon, A. Ozonation of sunflower oils: Impact of experimental conditions on the composition and the antibacterial activity of ozonized oils. Chem. Phys. Lipids. 2015, 186, 79–85. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kang, D.-J.; Nam, H.-S.; Kim, J.-H.; Kim, S.-Y.; Lee, J.-O.; Kim, B.-J. Ozonated sunflower oil exerted protective effect for embryo and cell survival via potent reduction power and antioxidant activity in HDL with strong antimicrobial activity. Antioxidants 2021, 10, 1651. [Google Scholar] [CrossRef]
- Hernández, F.; Menéndez, S.; Wong, R. Decrease of blood cholesterol and stimulation of antioxidative response in cardiopathy patients treated with endovenous ozone therapy. Free Radic. Biol. Med. 1995, 19, 115–119. [Google Scholar] [CrossRef]
- Oliveira, N.F.; Ferreira, C.S.; Silva, L.F.; Santos, I.P.; Jacob, J.C.; Penna, B.A.; Pinna, A.E. Use of ozonated sunflower oil in the treatment of infectious endometritis in mares. Equine Vet. Sci. 2023, 125, 104735. [Google Scholar] [CrossRef]
- Rodriguez, Z.Z.; Lemus, M.; González, E.F.; Lozano, O.E.L. Efficacy of ozonized sunflower oil as treatment of canine generalized demodicosis. Insights Vet. Sci. 2021, 5, 15–21. [Google Scholar]
- Menéndez, S.; Falcón, L.; Maqueira, Y. Therapeutic efficacy of topical OLEOZON® in patients suffering from onychomycosis. Mycoses 2011, 54, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Menéndez, S.; Falcón, L.; Simón, D.R.; Landa, N. Efficacy of ozonized sunflower oil in the treatment of tinea pedis. Mycoses 2002, 45, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Silva Filho, A.B.; Sobral, G.G.; Freire, L.Q.; Silva, E.R.; Vazquez, J.J.; Serres, C.; Lorenzo, P.L.; Gutiérrez-Cepeda, L.; Carneiro, G.F. Anti-biofilm action of ozonized sunflower oil against bacteria isolated from the uterus of mares susceptible to endometritis. J. Equine Vet. Sci. 2023, 125, 104746. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Lee, J.O.; Lee, S.; Heo, J.; Cho, K.-H.; Bahuguna, A.; Yoo, K.-H.; Kim, B.J. Ozonated sunflower oil (OSO) alleviates inflammatory responses in oxazolone-induced atopic dermatitis (AD)-like mice and LPS- treated RAW 264.7 cells. J. Microbiol. Biotechnol. 2024, 34, 1–10. [Google Scholar] [CrossRef]
- Cho, K.-H.; Bahuguna, A.; Kang, D.-J.; Kim, J.-E. Prolonged supplementation of ozonated sunflower oil Bestows an antiaging effect, improves blood lipid profile and spinal deformities, and protects vital organs of zebrafish (Danio rerio) against age-related degeneration: Two-years consumption study. Antioxidants 2024, 13, 123. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-E.; Bahuguna, A.; Kang, D.-J. Long-term supplementation of ozonated sunflower oil improves dyslipidemia and hepatic inflammation in hyperlipidemic zebrafish: Suppression of oxidative stress and inflammation against carboxymethyllysine toxicity. Antioxidants 2023, 12, 1240. [Google Scholar] [CrossRef]
- Zamora Rodríguez, Z.B.; González Alvarez, R.; Guanche, D.; Merino, N.; Hernández Rosales, F.; Menéndez Cepero, S.; Alonso González, Y.; Schulz, S. Antioxidant mechanism is involved in the gastroprotective effects of ozonized sunflower oil in ethanol-induced ulcers in rats. Mediat. Inflamm. 2007, 2007, 65873. [Google Scholar] [CrossRef]
- Machholz, E.; Mulder, G.; Ruiz, C.; Corning, B.F.; Pritchett-Corning, K.R. Manual restraint and common compound administration routes in mice and rats. J. Vis. Exp. 2012, 67, e2771. [Google Scholar]
- Cho, K.-H.; Kim, S.-J.; Yadav, D.; Kim, J.-Y.; Kim, J.-R. Consumption of Cuban policosanol improves blood pressure and lipid profile via enhancement of HDL functionality in healthy women subjects: Randomized, double-blinded, and placebo-controlled study. Oxid. Med. Cell. Longev. 2018, 2018, 4809525. [Google Scholar] [CrossRef]
- Cho, K.-H.; Baek, S.-H.; Nam, H.-S.; Bahuguna, A. Enhancement of antioxidant and anti-glycation properties of beeswax alcohol in reconstituted high-density lipoprotein: Safeguarding against carboxymethyllysine toxicity in zebrafish. Antioxidants 2023, 12, 2116. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Bieber, L.; Tolbert, N. A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-R.; Lee, I.-C.; Kwon, H.-J. Native high-density lipoproteins (HDL) with higher paraoxonase exert a potent antiviral effect against SARS-CoV-2 (COVID-19), while glycated HDL lost the antiviral activity. Antioxidants 2021, 10, 209. [Google Scholar] [CrossRef]
- OECD. Test No. 203: Fish, Acute Toxicity Testing. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2019. [Google Scholar]
- Cho, K.-H.; Kim, J.-E.; Nam, H.-S.; Kang, D.-J.; Na, H.-J. Anti-inflammatory activity of CIGB-258 against acute toxicity of carboxymethyllysine in paralyzed zebrafish via enhancement of high-density lipoproteins stability and functionality. Int. J. Mol. Sci. 2022, 23, 10130. [Google Scholar] [CrossRef]
- Hsieh, C.; Rajashekaraiah, V. Ferric Reducing Ability of Plasma: A Potential Oxidative Stress Marker in Stored Plasma. Acta Haematol. Pol. 2021, 52, 61–67. [Google Scholar] [CrossRef]
- Inayama, T.; Kumagai, Y.; Sakane, M.; Saito, M.; Matsuda, M. Plasma protein-bound sulfhydryl group oxidation in humans following a full marathon race. Life Sci. 1996, 59, 573–578. [Google Scholar] [CrossRef]
- Başkol, M.; Dolbun Seçkin, K.; Başkol, G. Advanced oxidation protein products, total thiol levels and total oxidant/antioxidant status in patients with nash. Turk. J. Gastroenterol. 2014, 25, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.-L. Measurement of protein thiol groups and glutathione in plasma. Methods Enzym. 1994, 233, 380–385. [Google Scholar]
- Lee, B.H.; Hsu, W.H.; Hsu, Y.W.; Pan, T.M. Suppression of dimerumic acid on hepatic fibrosis caused from carboxymethyllysine (CML) by attenuating oxidative stress depends on Nrf2 activation in hepatic stellate cells (HSCs). Food Chem. Toxicol. 2013, 62, 413–419. [Google Scholar] [CrossRef]
- Boesten, D.M.P.H.J.; Elie, A.G.I.M.; Drittij-Reijnders, M.-J.; den Hartog, G.J.M.; Bast, A. Effect of Nε-carboxymethyllysine on oxidative stress and the glutathione system in beta cells. Toxicol. Rep. 2014, 1, 973–980. [Google Scholar] [CrossRef]
- Kim, J.; Kim, C.S.; Sohn, E.; Kim, J.S. Elevated N ε-(Carboxymethyl) lysine is associated with apoptosis of retinal pericytes in streptozotocin-induced diabetic rats. Ophthalmic Res. 2011, 46, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Hoang, A.; Murphy, A.J.; Coughlan, M.T.; Thomas, M.C.; Forbes, J.M.; O’Brien, R.; Cooper, M.E.; Chin-Dusting, J.P.F.; Sviridov, D. Advanced glycation of apolipoprotein A-I impairs its anti-atherogenic properties. Diabetologia 2007, 50, 1770–1779. [Google Scholar] [CrossRef]
- Kashyap, S.R.; Osme, A.; Ilchenko, S.; Golizeh, M.; Lee, K.; Wang, S.; Bena, J.; Previs, S.F.; Smith, J.D.; Kasumov, T. Glycation reduces the stability of apoAI and increases HDL dysfunction in diet-controlled type 2 diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 388–396. [Google Scholar] [CrossRef]
- Pu, L.J.; Lu, L.; Zhang, R.Y.; Du, R.; Shen, Y.; Zhang, Q.; Yang, Z.K.; Chen, Q.J.; Shen, W.F. Glycation of apoprotein A-I is associated with coronary artery plaque progression in type 2 diabetic patients. Diabetes Care 2013, 36, 1312–1320. [Google Scholar] [CrossRef]
- Cho, K.H. The Current Status of Research on High-Density Lipoproteins (HDL): A paradigm shift from HDL quantity to HDL quality and HDL functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [PubMed]
- López-Olmos, V.; Luna-Luna, M.; Carreón-Torres, E.; González-Pacheco, H.; Bautista-Pérez, R.; Posadas-Sánchez, R.; Fragoso, J.M.; Vargas-Alarcón, G.; Pérez-Méndez, Ó. Trp fluorescence redshift during HDL apolipoprotein denaturation is increased in patients with coronary cyndrome in acute phase: A new assay to evaluate HDL stability. Int. J. Mol. Sci. 2021, 22, 7819. [Google Scholar] [CrossRef]
- Shokri, Y.; Variji, A.; Nosrati, M.; Khonakdar-Tarsi, A.; Kianmehr, A.; Kashi, Z.; Bahar, A.; Bagheri, A.; Mahrooz, A. Importance of paraoxonase 1 (PON1) as an antioxidant and antiatherogenic enzyme in the cardiovascular complications of type 2 diabetes: Genotypic and phenotypic evaluation. Diabetes Res. Clin. Pract. 2020, 161, 108067. [Google Scholar] [CrossRef]
- Márquez, A.B.; Nazir, S.; van der Vorst, E.P.C. High-density lipoprotein modifications: A pathological consequence or cause of disease progression? Biomedicines 2020, 8, 549. [Google Scholar] [CrossRef]
- Mackness, M.I.; Arrol, S.; Durrington, P.N. Paraoxonase prevents accumulation of lipoperoxides in low-density lipoprotein. FEBS Lett. 1991, 286, 152–154. [Google Scholar] [CrossRef]
- Brites, F.; Martin, M.; Guillas, I.; Kontush, A. Antioxidative activity of high-density lipoprotein (HDL): Mechanistic insights into potential clinical benefit. BBA Clin. 2017, 8, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Mastorikou, M.; Mackness, B.; Liu, Y.; Mackness, M. Glycation of paraoxonase-1 inhibits its activity and impairs the ability of high-density lipoprotein to metabolize membrane lipid hydroperoxides. Diabet. Med. 2008, 25, 1049–1055. [Google Scholar] [CrossRef]
- Gordts, S.C.; Singh, N.; Muthuramu, I.; Geest, B. Pleiotropic effects of HDL: Towards new therapeutic areas for HDL-targeted interventions. Curr. Mol. Med. 2014, 14, 481–503. [Google Scholar] [CrossRef]
- Boyce, G.; Button, E.; Soo, S.; Wellington, C. The pleiotropic vasoprotective functions of high-density lipoproteins (HDL). J. Biomed. Res. 2017, 32, 164. [Google Scholar]
- Denimal, D. Antioxidant and Anti-inflammatory functions of high-density lipoprotein in type 1 and type 2 diabetes. Antioxidants 2024, 13, 57. [Google Scholar] [CrossRef]
- Barter, P.J.; Nicholls, S.; Rye, K.-A.; Anantharamaiah, G.M.; Navab, M.; Fogelman, A.M. Antiinflammatory properties of HDL. Circ. Res. 2004, 95, 764–772. [Google Scholar] [CrossRef]
- Rader, D.J.; Hovingh, G.K. HDL and cardiovascular disease. Lancet 2014, 384, 618–625. [Google Scholar] [CrossRef]
- Marsillach, J.; Adorni, M.P.; Zimetti, F.; Papotti, B.; Zuliani, G.; Cervellati, C. HDL Proteome and alzheimer’s disease: Evidence of a link. Antioxidants 2020, 9, 1224. [Google Scholar] [CrossRef]
- Jomard, A.; Osto, E. High density lipoproteins: Metabolism, function, and therapeutic potential. Front. Cardiovasc. Med. 2020, 7, 39. [Google Scholar] [CrossRef]
- Delli Bovi, A.P.; Marciano, F.; Mandato, C.; Siano, M.A.; Savoia, M.; Vajro, P. Oxidative Stress in non-alcoholic fatty liver disease. An updated mini review. Front. Med. 2021, 8, 595371. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Najeeb, Q.; Sameer, A.S.; Aziz, R.; Hamid, S. Association of lipid profile and liver enzymes among non-alcoholic fatty liver disease patients attending a tertiary care hospital in northern Indian. Int. J. Curr. Res. 2015, 7, 14348–14352. [Google Scholar]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Bouwman, F.G.; van Baak, M.A.; Roumans, N.J.T.; Vink, R.G.; Mariman, E.C.M. Plasma levels of triglycerides and IL-6 are associated with weight regain and fat mass expansion. J. Clin. Endocrinol. Metab. 2022, 107, 1920–1929. [Google Scholar] [CrossRef]
- Nonogaki, K.; Fuller, G.M.; Fuentes, N.L.; Moser, A.H.; Staprans, I.; Grunfeld, C.; Feingold, K.R. Interleukin-6 stimulates hepatic triglyceride secretion in rats. Endocrinology 1995, 136, 2143–2149. [Google Scholar] [CrossRef]
- Feingold, K.R.; Grunfeld, C. The effect of inflammation and infection on lipids and lipoproteins. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | Group 3 | |
|---|---|---|---|
| Oral supplementation | DW | SO | OSO |
| Intake amount (g/kg BW/day) | 3.0 | 3.0 | 3.0 |
| Number of rats (week 0) | (n = 10) | (n = 10) | (n = 10) |
| Number of rats (week 4) | (n = 10) | (n = 10) | (n = 10) |
| Body weight (BW) week 0 (g) | 153.1 ± 6.3 ns,1) | 153.8 ± 6.5 ns,1) | 153.2 ± 6.4 ns,1) |
| Body weight (BW) week 4 (g) | 299.4 ± 22.0 ns,1), ***,2) | 300.4 ± 15.6 ns,1), ***,2) | 297.2 ± 13.5 ns,1), ***,2) |
| Body weight (BW) change (g) 3) | 146.3 ± 26.9 ns,1) | 146.6 ± 21.1 ns,1) | 144.0 ± 19.7 ns,1) |
| Parameters | Group 1 DW (n = 10) | Group 2 SO (n = 10) | Group 3 OSO (n = 10) | Statistical Difference (p Value) | ||
|---|---|---|---|---|---|---|
| DW vs. SO | SO vs. OSO | DW vs. OSO | ||||
| WBC (103/μL) | 8.1 ± 1.8 | 6.9 ± 1.8 | 8.7 ± 2.5 | 0.431 | 0.155 | 0.791 |
| RBC (106/μL) | 7.5 ± 0.2 | 7.6 ± 0.2 | 7.2 ± 0.3 | 0.914 | 0.006 | 0.017 |
| Hematocrit (%) | 44.2 ± 1.5 | 44.6 ± 1.1 | 42.8 ± 1.9 | 0.831 | 0.034 | 0.113 |
| Platelet (103/μL) | 1045 ± 82 | 1099 ± 113 | 1108 ± 106 | 0.468 | 0.978 | 0.359 |
| Neutrophil (%) | 9.8 ± 2.1 | 10.9 ± 3.1 | 10.7 ± 2.6 | 0.620 | 0.994 | 0.683 |
| Lymphocyte (%) | 83.6 ± 3.1 | 83.2 ± 3.4 | 83.5 ± 2.3 | 0.949 | 0.961 | 0.999 |
| Monocyte (%) | 5.2 ± 1.5 | 4.4 ± 0.9 | 4.6 ± 1.3 | 0.342 | 0.934 | 0.538 |
| Eosinophil (%) | 0.7 ± 0.2 | 0.9 ± 0.5 | 0.5 ± 0.1 | 0.322 | 0.009 | 0.202 |
| Large unstained cells (%) | 0.6 ± 0.1 | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.450 | 1.000 | 0.450 |
| Basophil (%) | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.936 | 1.000 | 0.936 |
| Parameters | Reference Range | Group 1 DW (n = 10) | Group 2 SO (n = 10) | Group 3 OSO (n = 10) | Statistical Difference (p Value) | ||
|---|---|---|---|---|---|---|---|
| DW vs. SO | SO vs. OSO | DW vs. OSO | |||||
| Albumin (g/dL) | 2.9–4.8 | 3.50 ± 0.08 | 3.49 ± 0.08 | 3.64 ± 0.12 | 0.973 | 0.012 | 0.007 |
| ALT (U/L) | 6–114 | 69.25 ± 15.7 | 68.38 ± 7.9 | 58.18 ± 4.1 | 0.998 | 0.740 | 0.774 |
| AST (U/L) | 37–205 | 91.21 ± 2.9 | 86.91 ± 8 | 76.08 ± 2.5 | 0.826 | 0.146 | 0.335 |
| Creatinine (mg/dL) | 0.4–1.5 | 0.57 ± 0.03 | 0.57 ± 0.02 | 0.58 ± 0.04 | 1.000 | 0.565 | 0.565 |
| CRP (mg/L) | 2.8 ± 0.1 | 2.8 ± 0.2 | 2.7 ± 0.2 | 0.890 | 0.890 | 0.631 | |
| γ-GTP (U/L) | 0.5–5.3 | 3.1 ± 0.4 | 3.1 ± 0.5 | 3.1 ± 0.5 | 0.979 | 0.898 | 0.967 |
| Glucose (mg/dL) | 44–131 | 109.3 ± 6.0 | 100.2 ± 7.0 | 102.3 ± 13.7 | 0.104 | 0.877 | 0.250 |
| Interleukin-6 (ng/mL) | - | 0.020 ± 0.001 | 0.024 ± 0.004 | 0.016 ± 0.001 | 0.418 | 0.044 | 0.414 |
| TC (mg/dL) | 40–281 | 100.7 ± 10.2 | 96.3 ± 1.8 | 83.7 ± 2.3 | 0.873 | 0.370 | 0.201 |
| TG (mg/dL) | 30–409 | 42.0 ± 4.0 | 42.3 ± 1.5 | 40.3 ± 4.4 | 0.998 | 0.918 | 0.942 |
| HDL-C (mg/dL) | 61.3 ± 7.9 | 55.3 ± 3.2 | 53.0 ± 2.6 | 0.702 | 0.945 | 0.523 | |
| LDL-C (mg/dL) | 19.0 ± 2.0 | 20.0 ± 1.0 | 12.7 ± 0.3 | 0.854 | 0.017 | 0.032 | |
| % HDL (HDL/TC) | 60.6 ± 1.7 | 57.4 ± 2.4 | 63.3 ± 1.4 | 0.485 | 0.142 | 0.592 | |
| TG/HDL | 0.7 ± 0.0 | 0.8 ± 0.0 | 0.8 ± 0.1 | 0.483 | 1.000 | 0.483 | |
| LDL/HDL | 0.3 ± 0.0 | 0.4 ± 0.0 | 0.2 ± 0.0 | 0.269 | 0.031 | 0.269 | |
| RC (mg/dL) | 20.3 ± 0.7 | 21.0 ± 2.0 | 18.0 ± 0.6 | 0.927 | 0.287 | 0.442 | |
| Group 1 DW (n = 10) | Group 2 SO (n = 10) | Group 3 OSO (n = 10) | Statistical Difference (p Value) | ||||
|---|---|---|---|---|---|---|---|
| DW vs. SO | SO vs. OSO | DW vs. OSO | |||||
| LDL | TC (mg/mL) | 75.9 ± 4.8 | 84.5 ± 3.5 | 87.2 ± 4.3 | 0.300 | 0.760 | 0.146 |
| TG (mg/mL) | 23.9 ± 2.8 | 23.3 ± 3.6 | 23.3 ± 1.6 | 0.932 | 1.000 | 0.789 | |
| MDA (μM) | 6.1 ± 0.4 | 6.2 ± 1.0 | 5.2 ± 0.8 | 0.969 | 0.242 | 0.343 | |
| FI (Glycated) | 5583 ± 298 | 5452 ± 214 | 5140 ± 145 | 1.000 | 0.238 | 0.047 | |
| Size (nm2) | 360.1 ± 132.7 | 387.2 ± 121.9 | 363.3 ± 155.1 | 0.651 | 0.716 | 0.994 | |
| HDL | TC (mg/mL) | 107.4 ± 3.2 | 98.3 ± 6.5 | 99.0 ± 3.9 | 0.442 | 0.952 | 0.067 |
| TG (mg/mL) | 4.0 ± 0.2 | 4.1 ± 0.2 | 3.5 ± 0.2 | 0.724 | 0.157 | 0.314 | |
| MDA (μM) | 4.1 ± 1.0 | 4.8 ± 0.4 | 3.2 ± 0.4 | 0.548 | 0.018 | 0.434 | |
| FI (Glycated) | 1941 ± 73 | 1832 ± 41 | 1752 ± 64 | 0.098 | 0.318 | 0.002 | |
| Size (nm2) | 199.6 ± 67.1 | 166.9 ± 44.7 | 260.1 ± 88.7 | 0.092 | <0.001 | <0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.-H.; Kim, J.-E.; Lee, M.-S.; Bahuguna, A. Oral Supplementation of Ozonated Sunflower Oil Augments Plasma Antioxidant and Anti-Inflammatory Abilities with Enhancement of High-Density Lipoproteins Functionality in Rats. Antioxidants 2024, 13, 529. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050529
Cho K-H, Kim J-E, Lee M-S, Bahuguna A. Oral Supplementation of Ozonated Sunflower Oil Augments Plasma Antioxidant and Anti-Inflammatory Abilities with Enhancement of High-Density Lipoproteins Functionality in Rats. Antioxidants. 2024; 13(5):529. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050529
Chicago/Turabian StyleCho, Kyung-Hyun, Ji-Eun Kim, Myeong-Sung Lee, and Ashutosh Bahuguna. 2024. "Oral Supplementation of Ozonated Sunflower Oil Augments Plasma Antioxidant and Anti-Inflammatory Abilities with Enhancement of High-Density Lipoproteins Functionality in Rats" Antioxidants 13, no. 5: 529. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050529