
Unravelling the Role of Candida albicans Prn1 in the Oxidative Stress Response through a Proteomics Approach
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Experimental Design
2.3. Cell Disruption and Protein Extract Quantification
2.4. Proteomics Assay
2.5. Protein Quantification
2.6. GO Enrichment Analysis and Protein Clustering
2.7. Viability Assays
2.8. ROS Detection
2.9. Cellular Apoptosis
2.10. Mitochondrial Function
2.11. Proteasome Activity
3. Results
3.1. Impact of PRN1 Deletion on Cell Death and Recovery under Oxidative Stress
3.2. Proteomic Response of SN250 and prn1∆ Strains to H2O2 Treatment
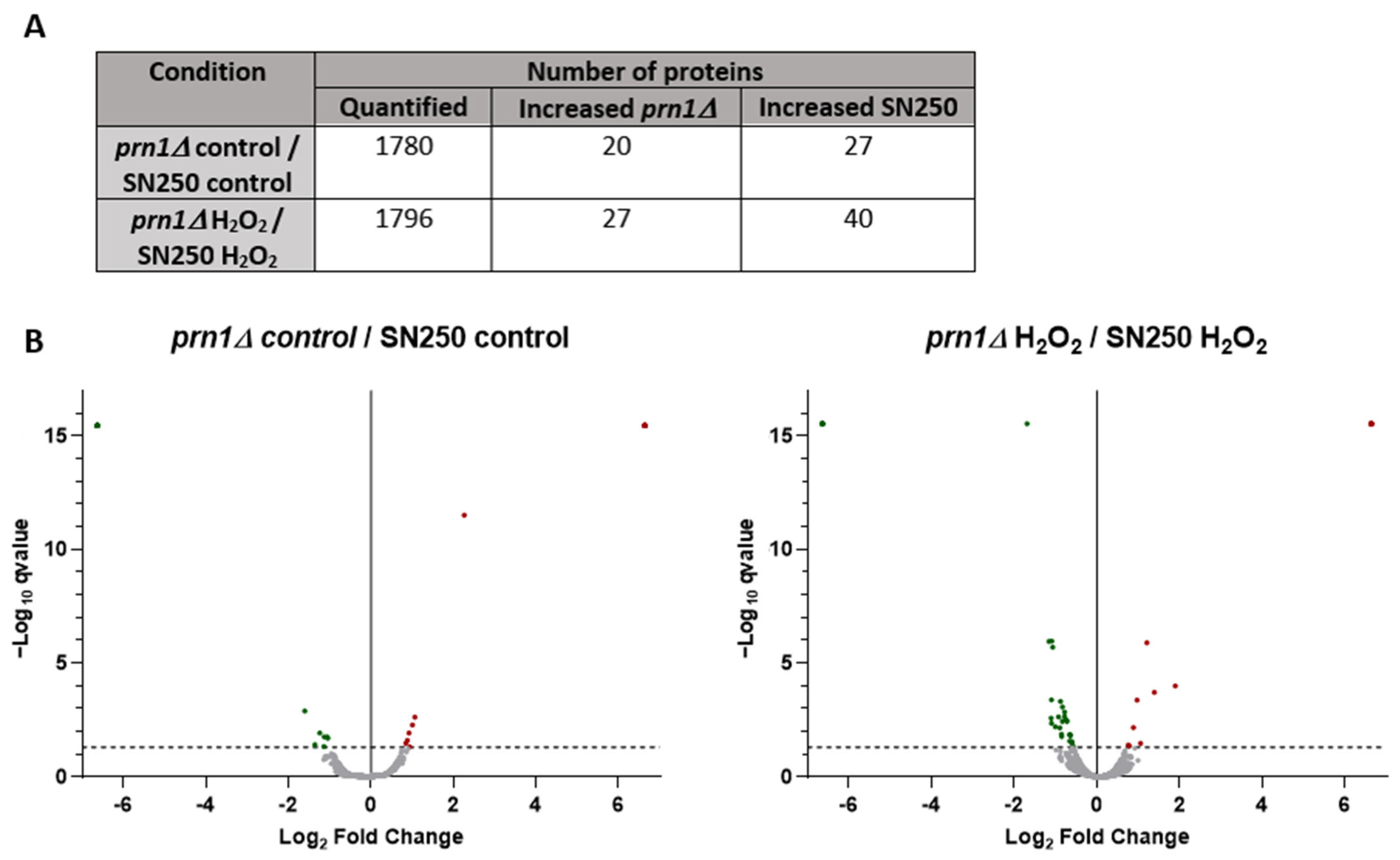
3.3. Comparative Proteomics Analysis between SN250 and prn1∆ Strains Either in the Presence or the Absence of Oxidative Stress
3.4. Influence of Prn1 Deletion on ROS Production and Proteasome Function
3.5. C. albicans orf19.4850 Mutant Response to Oxidative Stress
4. Discussion
4.1. Prn1 Has a Main Role in the Oxidative Response
4.2. Prn1 Absence Increases ROS Levels after H2O2 Treatment
4.3. Absence of Prn1 Increases Proteasome Activity and Decreases Proteins Related to Translation after H2O2 Treatment
4.4. Differential Expression of Transcription Factors between Strains
4.5. Prn1 May Be Related to Mitochondrial Oxidative Stress Detoxication in Basal Conditions
4.6. Involvement in the Oxidative Stress Response of orf19.4850, an S. cerevisiae CUB1 Orthologue
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans Biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans—The Virulence Factors and Clinical Manifestations of Infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W. Global Incidence and Mortality of Severe Fungal Disease. Lancet Infect. Dis. 2024, ahead of print. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: The World Health Organization (WHO) Fungal Priority Pathogens List in Response to Emerging Fungal Pathogens during the COVID-19 Pandemic. Med. Sci. Monit. 2022, 28, e939088-1–e939088-3. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Song, Y.G.; Li, R.Y. A Ten-Year Retrospective Study of Invasive Candidiasis in a Tertiary Hospital in Beijing. Biomed. Environ. Sci. 2021, 34, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, R.; Luan, Z.; Ma, X. Risk of Invasive Candidiasis with Prolonged Duration of ICU Stay: A Systematic Review and Meta-Analysis. BMJ Open 2020, 10, e036452. [Google Scholar] [CrossRef] [PubMed]
- Molero, G.; Guillén, M.V.; Martínez-Solano, L.; Gil, C.; Pla, J.; Nombela, C.; Sánchez-Pérez, M.; Diez-Orejas, R. The Importance of the Phagocytes’ Innate Response in Resolution of the Infection Induced by a Low Virulent Candida albicans Mutant. Scand. J. Immunol. 2005, 62, 224–233. [Google Scholar] [CrossRef]
- Diez-Orejas, R.; Molero, G.; Moro, M.A.; Gil, C.; Nombela, C.; Sánchez-Pérez, M. Two Different NO-Dependent Mechanisms Account for the Low Virulence of a Non-Mycelial Morphological Mutant of Candida albicans. Med. Microbiol. Immunol. 2001, 189, 153–160. [Google Scholar] [CrossRef]
- Cabezón, V.; Vialás, V.; Gil-Bona, A.; Reales-Calderón, J.A.; Martínez-Gomariz, M.; Gutiérrez-Blázquez, D.; Monteoliva, L.; Molero, G.; Ramsdale, M.; Gil, C. Apoptosis of Candida albicans during the Interaction with Murine Macrophages: Proteomics and Cell-Death Marker Monitoring. J. Proteome Res. 2016, 15, 1418–1434. [Google Scholar] [CrossRef]
- Fernández-Arenas, E.; Cabezón, V.; Bermejo, C.; Arroyo, J.; Nombela, C.; Diez-Orejas, R.; Gil, C. Integrated Proteomics and Genomics Strategies Bring New Insight into Candida albicans Response upon Macrophage Interaction. Mol. Cell Proteom. 2007, 6, 460–478. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Bender, J.A.; Fink, G.R. Transcriptional Response of Candida albicans upon Internalization by Macrophages. Eukaryot. Cell 2004, 3, 1076. [Google Scholar] [CrossRef] [PubMed]
- Alarco, A.M.; Raymond, M. The BZip Transcription Factor Cap1p Is Involved in Multidrug Resistance and Oxidative Stress Response in Candida albicans. J. Bacteriol. 1999, 181, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Yang, C.Y.; Lan, C.Y. Candida albicans Hap43 Is a Repressor Induced under Low-Iron Conditions and Is Essential for Iron-Responsive Transcriptional Regulation and Virulence. Eukaryot. Cell 2011, 10, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Dantas, A.D.S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative Stress Responses in the Human Fungal Pathogen, Candida albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Monge, R.; Navarro-García, F.; Román, E.; Negredo, A.I.; Eisman, B.; Nombela, C.; Pla, J. The Hog1 Mitogen-Activated Protein Kinase Is Essential in the Oxidative Stress Response and Chlamydospore Formation in Candida albicans. Eukaryot. Cell 2003, 2, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Amador-García, A.; Zapico, I.; Borrajo, A.; Malmström, J.; Monteoliva, L.; Gil, C. Extending the Proteomic Characterization of Candida albicans Exposed to Stress and Apoptotic Inducers through Data-Independent Acquisition Mass Spectrometry. mSystems 2021, 6, e0094621. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H:Quinone Acceptor Oxidoreductase 1 (NQO1), a Multifunctional Antioxidant Enzyme and Exceptionally Versatile Cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef]
- Hübner, R.H.; Schwartz, J.D.; De, B.P.; Ferris, B.; Omberg, L.; Mezey, J.G.; Hackett, N.R.; Crystal, R.G. Coordinate Control of Expression of Nrf2-Modulated Genes in the Human Small Airway Epithelium Is Highly Responsive to Cigarette Smoking. Mol. Med. 2009, 15, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Bartlam, M.; Zeng, Q.; Miyatake, H.; Hisano, T.; Miki, K.; Wong, L.L.; Gao, G.F.; Rao, Z. Crystal Structure of Human Pirin: An Iron-Binding Nuclear Protein and Transcription Cofactor. J. Biol. Chem. 2004, 279, 1491–1498. [Google Scholar] [CrossRef]
- Barman, A.; Hamelberg, D. Fe(II)/Fe(III) Redox Process Can Significantly Modulate the Conformational Dynamics and Electrostatics of Pirin in NF-κB Regulation. ACS Omega 2016, 1, 837–842. [Google Scholar] [CrossRef]
- Talà, A.; Damiano, F.; Gallo, G.; Pinatel, E.; Calcagnile, M.; Testini, M.; Fico, D.; Rizzo, D.; Sutera, A.; Renzone, G.; et al. Pirin: A Novel Redox-Sensitive Modulator of Primary and Secondary Metabolism in Streptomyces. Metab. Eng. 2018, 48, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; French, S.; Kohn, L.A.; Chen, V.; Johnson, A.D. Systematic Screens of a Candida albicans Homozygous Deletion Library Decouple Morphogenetic Switching and Pathogenicity. Nat. Genet. 2010, 42, 590. [Google Scholar] [CrossRef] [PubMed]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, Encapsulated Proteomic-Sample Processing Applied to Copy-Number Estimation in Eukaryotic Cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, M.B.; Costanzo, M.C.; Skrzypek, M.S.; Binkley, G.; Lane, C.; Miyasato, S.R.; Sherlock, G. The Candida Genome Database (CGD), a Community Resource for Candida albicans Gene and Protein Information. Nucleic Acids Res. 2005, 33, D358–D363. [Google Scholar] [CrossRef] [PubMed]
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of Assembly 22, Systematic Identifiers and Visualization of High Throughput Sequencing Data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef] [PubMed]
- Firestone, K.; Awonusi, D.; Panfair, D.; Roland, D.; Ramamurthy, A.; Kusmierczyk, A.R. YPL260W, a High-Copy Suppressor of a Copper-Sensitive Phenotype in Yeast, Is Linked to DNA Repair and Proteasome Function. Plant Gene 2016, 5, 38–48. [Google Scholar] [CrossRef]
- Hu, N.; Bai, L.; Dai, E.; Han, L.; Kang, R.; Li, H.; Tang, D. Pirin Is a Nuclear Redox-Sensitive Modulator of Autophagy-Dependent Ferroptosis. Biochem. Biophys. Res. Commun. 2021, 536, 100–106. [Google Scholar] [CrossRef]
- Adams, M.; Jia, Z. Structural and Biochemical Analysis Reveal Pirins to Possess Quercetinase Activity. J. Biol. Chem. 2005, 280, 28675–28682. [Google Scholar] [CrossRef]
- Gelbman, B.D.; Heguy, A.; O’Connor, T.P.; Zabner, J.; Crystal, R.G. Upregulation of Pirin Expression by Chronic Cigarette Smoking Is Associated with Bronchial Epithelial Cell Apoptosis. Respir. Res. 2007, 8, 10. [Google Scholar] [CrossRef]
- Franken, J.; Bauer, F.F. Carnitine Supplementation Has Protective and Detrimental Effects in Saccharomyces cerevisiae That Are Genetically Mediated. FEMS Yeast Res. 2010, 10, 270–281. [Google Scholar] [CrossRef]
- du Plessis, M.; Franken, J.; Bauer, F.F. Carnitine Requires Choline to Exert Physiological Effects in Saccharomyces cerevisiae. Front. Microbiol. 2018, 9, 360360. [Google Scholar] [CrossRef] [PubMed]
- Ashoori, M.; Saedisomeolia, A. Riboflavin (Vitamin B2) and Oxidative Stress: A Review. Br. J. Nutr. 2014, 111, 1985–1991. [Google Scholar] [CrossRef] [PubMed]
- Boretsky, Y.R.; Protchenko, O.V.; Prokopiv, T.M.; Mukalov, I.O.; Fedorovych, D.V.; Sibirny, A.A. Mutations and Environmental Factors Affecting Regulation of Riboflavin Synthesis and Iron Assimilation Also Cause Oxidative Stress in the Yeast Pichia Guilliermondii. J. Basic. Microbiol. 2007, 47, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Perez-Dominguez, F.; Carrillo-Beltrán, D.; Blanco, R.; Muñoz, J.P.; León-Cruz, G.; Corvalan, A.H.; Urzúa, U.; Calaf, G.M.; Aguayo, F. Role of Pirin, an Oxidative Stress Sensor Protein, in Epithelial Carcinogenesis. Biology 2021, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Rehmani, I.; Esaki, S.; Fu, R.; Chen, L.; De Serrano, V.; Liu, A. Pirin Is an Iron-Dependent Redox Regulator of NF-ΚB. Proc. Natl. Acad. Sci. USA 2013, 110, 9722–9727. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Kriete, A.; Sacan, A.; Michal Jazwinski, S. Comparing the Yeast Retrograde Response and NF-ΚB Stress Responses: Implications for Aging. Aging Cell 2010, 9, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, D.; Nobile, C.J.; Dong, D.; Ni, Q.; Su, T.; Jiang, C.; Peng, Y. Systematic Identification and Characterization of Five Transcription Factors Mediating the Oxidative Stress Response in Candida albicans. Microb. Pathog. 2023, 187, 106507. [Google Scholar] [CrossRef] [PubMed]
- Ramsdale, M.; Selway, L.; Stead, D.; Walker, J.; Yin, Z.; Nicholls, S.M.; Crowe, J.; Sheils, E.M.; Brown, A.J.P. MNL1 Regulates Weak Acid-Induced Stress Responses of the Fungal Pathogen Candida albicans. Mol. Biol. Cell 2008, 19, 4393–4403. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Bernardo, R.; Wang, C.; Schröder, M.S.; Pedro, N.A.; Butler, G.; Azeredo, J.; Henriques, M.; Pereira Mira, N.; Silva, S. Effect of Progesterone on Candida albicans Biofilm Formation under Acidic Conditions: A Transcriptomic Analysis. Int. J. Med. Microbiol. 2020, 310, 151414. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zaurin, R.; Beato, M.; Peterson, C.L. Swi3p Controls SWI/SNF Assembly and ATP-Dependent H2A-H2B Displacement. Nat. Struct. Mol. Biol. 2007, 14, 540–547. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Protein | Description | SN250 H2O2/ SN250 Control Ratio Log2 | prn1∆ H2O2/ prn1∆ Control Ratio Log2 |
|---|---|---|---|---|
| SN250 | Psa2 | Mannose-1-phosphate guanyltransferase | 5.36 | NS |
| Icl1 | Isocitrate lyase; glyoxylate cycle enzyme | 4.63 | 3.82 | |
| Prn1 | Protein with similarity to Pirin | 4.16 | ND | |
| Alt1 | Putative alanine transaminase | 4.13 | NS | |
| Oye22 | Putative NADPH dehydrogenase | 4.06 | 3.28 | |
| Acc1 | Putative acetyl-coenzyme-A carboxylases | 4.04 | 3.57 | |
| orf19.3477 | Putative pseudouridine synthase | 4 | 3.54 | |
| SN250 and prn1∆ | Cip1 | Possible oxidoreductase | 4.47 | 4.65 |
| Ahc1 | Ortholog(s) histone acetyltransferase activity | 4.13 | 4.23 | |
| Pim1 | ATP-dependent Lon protease | 3.96 | 4.17 | |
| prn1∆ | Mal2 | Alpha-glucosidase | 4.57 | + |
| Lys21 | Homocitrate synthase | 4.34 | 3.07 | |
| orf19.6035 | Unknown function | 4.27 | 3.89 | |
| Hsp21 | Small heat shock protein | 4.25 | 3.19 | |
| Qcr9 | Putative ubiquinol cytochrome c reductase | 4.19 | NS | |
| Nuc2 | Putative NADH-ubiquinone oxidoreductase | 4.15 | + | |
| orf19.7310 | Role in directing meiotic recombination | 4.09 | NS |
| Strain | Detected in Treated Condition | Detected in Basal Condition |
|---|---|---|
| SN250 H2O2/ SN250 control | Ald4, Bzz1, orf19.4430, orf19.4850, Rib2 | Hrr25, Mak11, Mcm3, orf19.1272, Ynd1 |
| prn1∆ H2O2/ prn1∆ control | Fmp28, Nit2, orf19.1150, orf19.2051, orf19.2452, Osh2, Phb2, Pr26, Rpn3, Rpt4, Rts1, Smp2, Snu66, Tad2, Tif33, Uso1, Yah1 | Afg2, Has1, Lig1, Msd1, Nrg1, orf19.5987, orf19.6445, Pnc1, Rpa43, Sec24, Trp1, Ubp12 |
| Protein | Description | SN250 H2O2/ SN250 Control Ratio Log2 | prn1∆ H2O2/ prn1∆ Control Ratio Log2 |
|---|---|---|---|
| Bas1 | Putative helix–loop–helix (HLH) transcription factor; role in filamentous growth | - | 3.61 |
| Cat5 | 2-octoprenyl-3-methyl-6-methoxy-1,4-benzoquinone hydroxylase activity; ubiquinone biosynthetic process | - | 2.75 |
| Dad4 | Subunit of the Dam1 (DASH) complex; chromosome segregation | −3.15 | 4.06 |
| Fmp53 | Ubiquinone-6 biosynthetic process | 3.69 | - |
| Ftr2 | High-affinity iron permease | −2.67 | 2.11 |
| Mfg1 | Regulator of filamentous growth; biofilm formation | - | 3.5 |
| Mnl1 | Transcription factor; induces transcripts of stress response genes via SLE (STRE-like) elements | - | + |
| orf19.764 | Ortholog(s)-negative regulation of TORC1 signaling | - | + |
| Rpo41 | Putative mitochondrial RNA polymerase; repressed in core stress response | - | 3.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arribas, V.; Monteoliva, L.; Hernáez, M.L.; Gil, C.; Molero, G. Unravelling the Role of Candida albicans Prn1 in the Oxidative Stress Response through a Proteomics Approach. Antioxidants 2024, 13, 527. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050527
Arribas V, Monteoliva L, Hernáez ML, Gil C, Molero G. Unravelling the Role of Candida albicans Prn1 in the Oxidative Stress Response through a Proteomics Approach. Antioxidants. 2024; 13(5):527. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050527
Chicago/Turabian StyleArribas, Victor, Lucia Monteoliva, María Luisa Hernáez, Concha Gil, and Gloria Molero. 2024. "Unravelling the Role of Candida albicans Prn1 in the Oxidative Stress Response through a Proteomics Approach" Antioxidants 13, no. 5: 527. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050527