

Lysyl Oxidase in Ectopic Cardiovascular Calcification: Role of Oxidative Stress
 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. VIC Isolation and Osteogenic Induction
2.3. Immunocytochemistry
2.4. Analysis of Calcium Deposition in VICs
2.5. Lentiviral Overexpression in VICs
2.6. LOX Silencing in VICs
2.7. Real-Time PCR
2.8. Animal Handling
2.9. Histology and Immunohistochemistry
2.10. ROS Detection
2.11. Statistical Analysis
3. Results
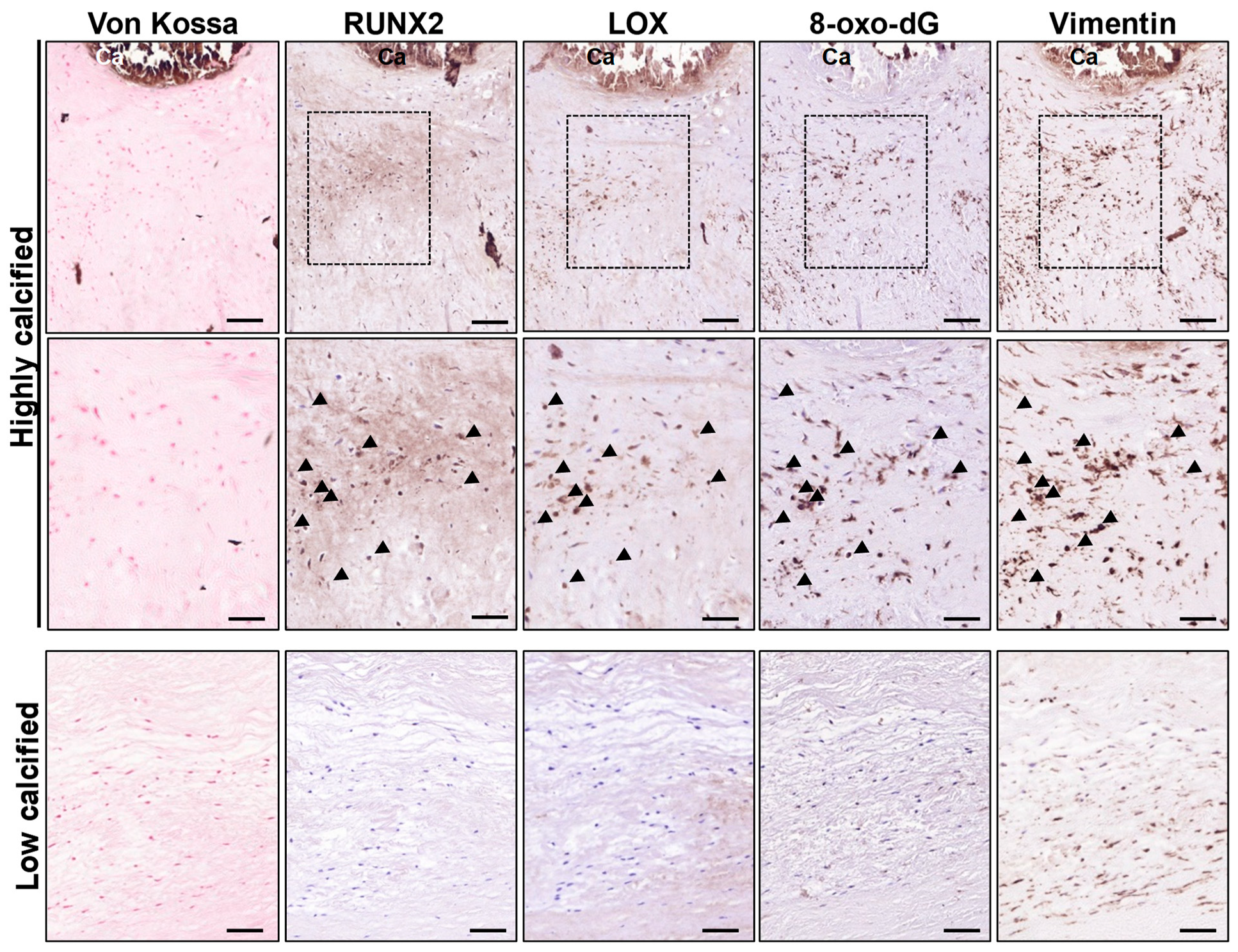
3.1. LOX Colocalized with 8-oxo-dG in Human-Calcified Atherosclerotic Lesions
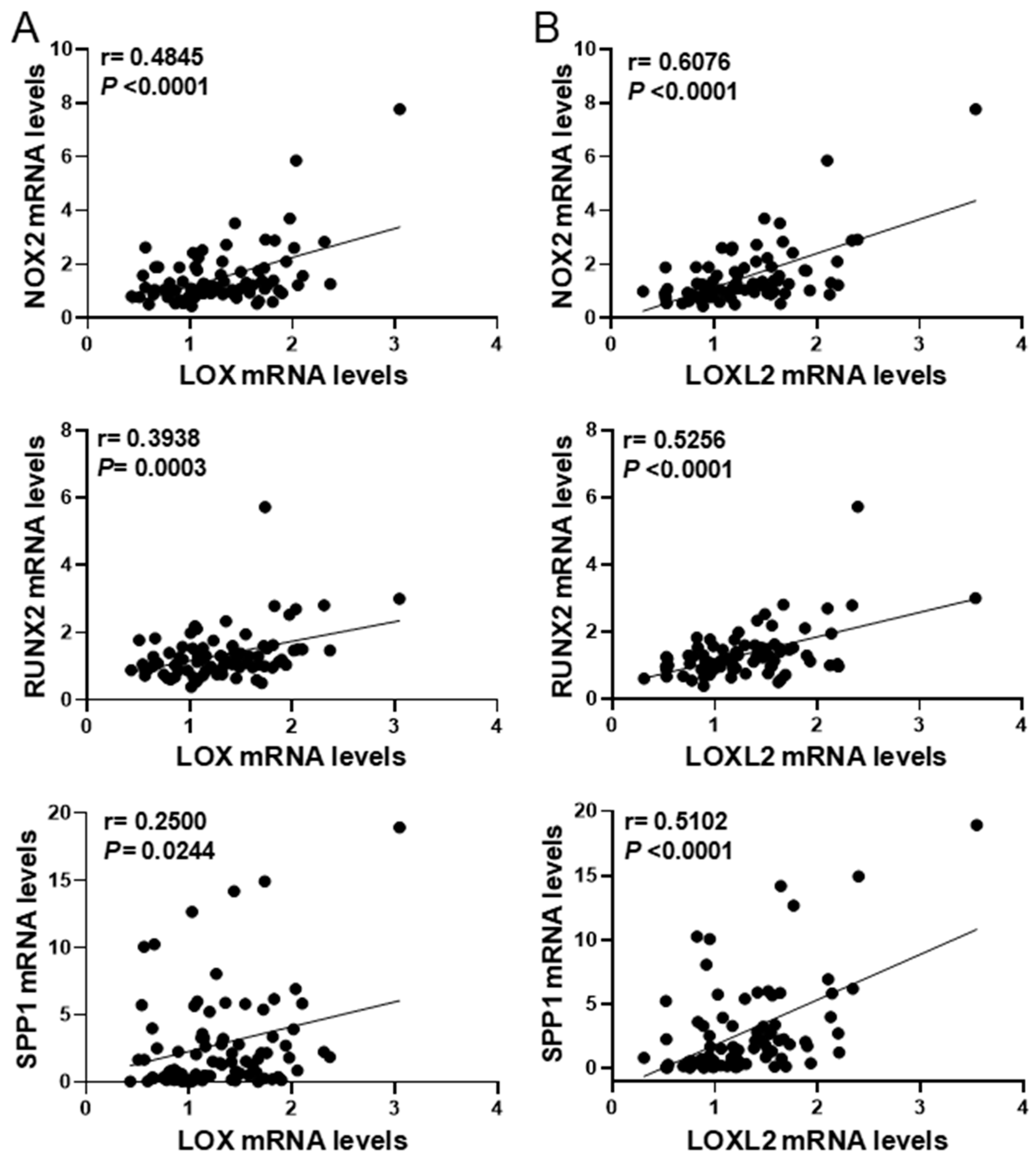
3.2. LOX Transgenesis Enhances Oxidative Stress in Atherosclerotic Lesions
3.3. LOX and Oxidative Stress in Calcified Human Aortic Valves
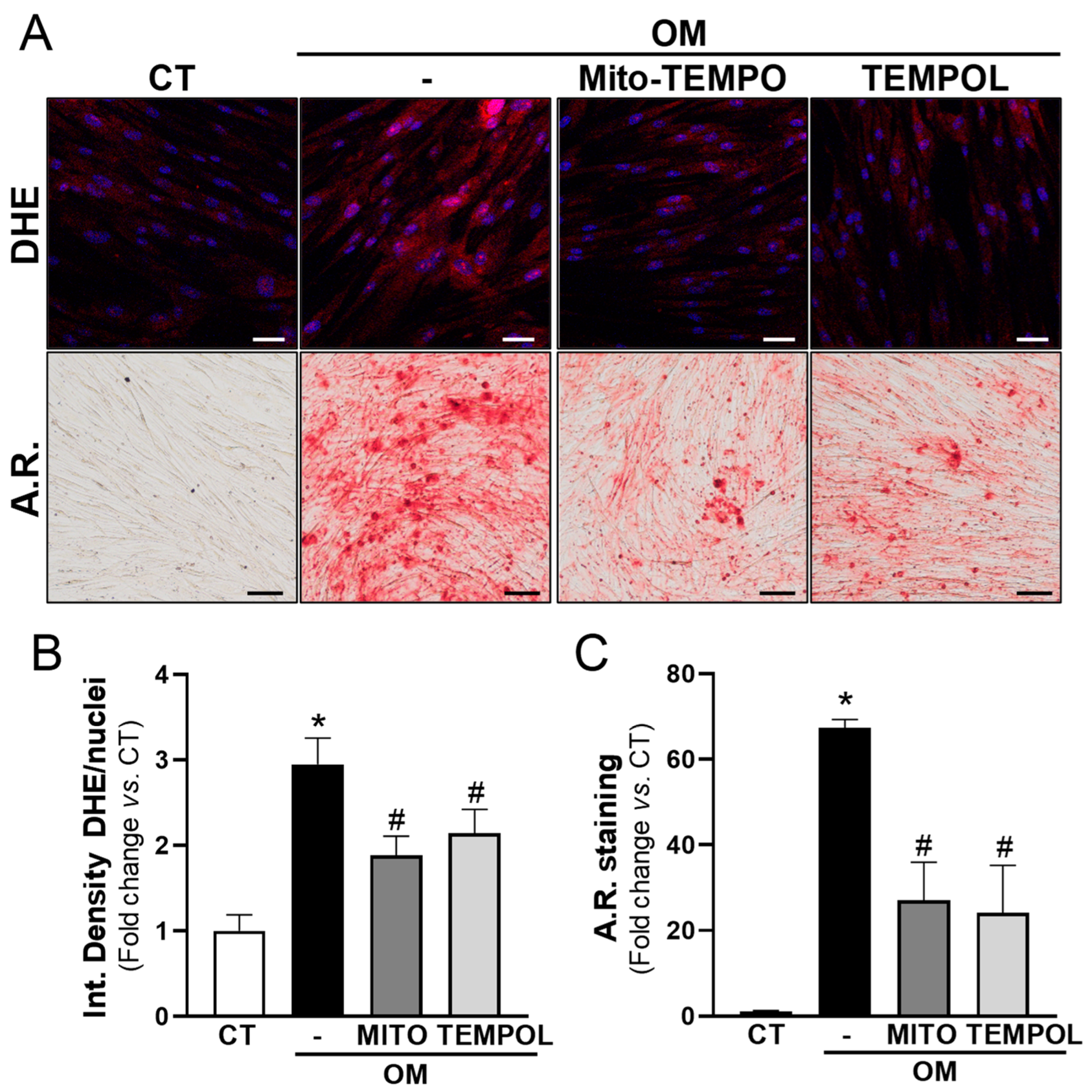
3.4. Antioxidant Treatment Prevents the Calcification of VIC Cultures Induced by High-Phosphate Media
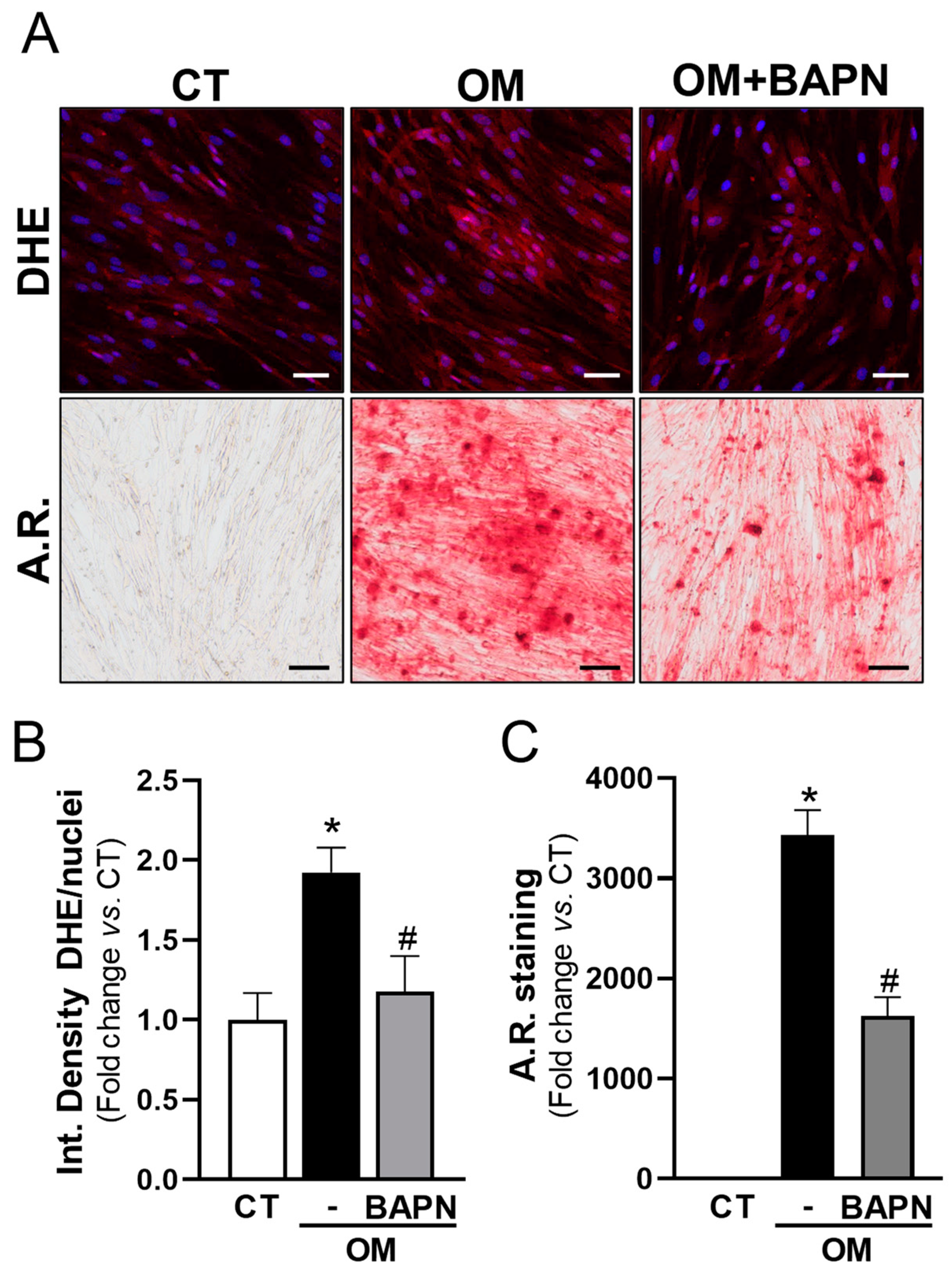
3.5. LOX Increases Oxidative Stress and Mineralization of VICs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoffmann, U.; Massaro, J.M.; D’Agostino, R.B.; Kathiresan, S.; Fox, C.S.; O’Donnell, C.J. Cardiovascular Event Prediction and Risk Reclassification by Coronary, Aortic, and Valvular Calcification in the Framingham Heart Study. J. Am. Heart Assoc. 2016, 5, e003144. [Google Scholar] [CrossRef]
- Martin, S.S.; Blaha, M.J.; Blankstein, R.; Agatston, A.; Rivera, J.J.; Virani, S.S.; Ouyang, P.; Jones, S.R.; Blumenthal, R.S.; Budoff, M.J.; et al. Dyslipidemia, coronary artery calcium, and incident atherosclerotic cardiovascular disease: Implications for statin therapy from the multi-ethnic study of atherosclerosis. Circulation 2014, 129, 77–86. [Google Scholar] [CrossRef]
- New, S.E.; Aikawa, E. Cardiovascular calcification: An inflammatory disease. Circ. J. 2011, 75, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Vliegenthart, R.; Oudkerk, M.; Hofman, A.; Oei, H.H.; van Dijck, W.; van Rooij, F.J.; Witteman, J.C. Coronary calcification improves cardiovascular risk prediction in the elderly. Circulation 2005, 112, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Lindman, B.R.; Clavel, M.A.; Mathieu, P.; Iung, B.; Lancellotti, P.; Otto, C.M.; Pibarot, P. Calcific aortic stenosis. Nat. Rev. Dis. Primers 2016, 2, 16006. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, J.D.; Goettsch, C.; Rogers, M.A.; Aikawa, E. Revisiting cardiovascular calcification: A multifaceted disease requiring a multidisciplinary approach. Semin Cell. Dev. Biol. 2015, 46, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Phua, K.; Chew, N.W.; Kong, W.K.; Tan, R.S.; Ye, L.; Poh, K.K. The mechanistic pathways of oxidative stress in aortic stenosis and clinical implications. Theranostics 2022, 12, 5189–5203. [Google Scholar] [CrossRef] [PubMed]
- Tóth, A.; Balogh, E.; Jeney, V. Regulation of Vascular Calcification by Reactive Oxygen Species. Antioxidants 2020, 9, 963. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, H.Z.E.; Zhao, G.; Shah, A.M.; Zhang, M. Role of oxidative stress in calcific aortic valve disease and its therapeutic implications. Cardiovasc Res. 2022, 118, 1433–1451. [Google Scholar] [CrossRef] [PubMed]
- Jover, E.; Silvente, A.; Marín, F.; Martínez-González, J.; Orriols, M.; Martinez, C.M.; Puche, C.M.; Valdés, M.; Rodriguez, C.; Hernández-Romero, D. Inhibition of enzymes involved in collagen cross-linking reduces vascular smooth muscle cell calcification. FASEB J. 2018, 32, 4459–4469. [Google Scholar] [CrossRef]
- Xue, Y.; Kossar, A.P.; Abramov, A.; Frasca, A.; Sun, M.; Zyablitskaya, M.; Paik, D.; Kalfa, D.; Della Barbera, M.; Thiene, G.; et al. Age-related enhanced degeneration of bioprosthetic valves due to leaflet calcification, tissue crosslinking, and structural changes. Cardiovasc. Res. 2023, 119, 302–315. [Google Scholar] [CrossRef]
- Ballester-Servera, C.; Alonso, J.; Cañes, L.; Vázquez-Sufuentes, P.; Puertas-Umbert, L.; Fernández-Celis, A.; Taurón, M.; Rodríguez-Sinovas, A.; López-Andrés, N.; Rodríguez, C.; et al. Lysyl oxidase-dependent extracellular matrix crosslinking modulates calcification in atherosclerosis and aortic valve disease. Biomed. Pharmacother. 2023, 167, 115469. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.; Martínez-González, J. The Role of Lysyl Oxidase Enzymes in Cardiac Function and Remodeling. Cells 2019, 8, 1483. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, J.; Varona, S.; Cañes, L.; Galán, M.; Briones, A.M.; Cachofeiro, V.; Rodríguez, C. Emerging Roles of Lysyl Oxidases in the Cardiovascular System: New Concepts and Therapeutic Challenges. Biomolecules 2019, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Galán, M.; Varona, S.; Guadall, A.; Orriols, M.; Navas, M.; Aguiló, S.; de Diego, A.; Navarro, M.A.; García-Dorado, D.; Rodríguez-Sinovas, A.; et al. Lysyl oxidase overexpression accelerates cardiac remodeling and aggravates angiotensin II-induced hypertrophy. FASEB J. 2017, 31, 3787–3799. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Revelles, S.; García-Redondo, A.B.; Avendaño, M.S.; Varona, S.; Palao, T.; Orriols, M.; Roque, F.R.; Fortuño, A.; Touyz, R.M.; Martínez-González, J.; et al. Lysyl Oxidase Induces Vascular Oxidative Stress and Contributes to Arterial Stiffness and Abnormal Elastin Structure in Hypertension: Role of p38MAPK. Antioxid. Redox Signal. 2017, 27, 379–397. [Google Scholar] [CrossRef]
- Ballester-Servera, C.; Cañes, L.; Alonso, J.; Puertas-Umbert, L.; Vázquez-Sufuentes, P.; Taurón, M.; Roselló-Díez, E.; Marín, F.; Rodríguez, C.; Martínez-González, J. Upregulation of NOR-1 in calcified human vascular tissues: Impact on osteogenic differentiation and calcification. Transl. Res. 2024, 264, 1–14. [Google Scholar] [CrossRef]
- Garaikoetxea, M.; Martín-Núñez, E.; Navarro, A.; Matilla, L.; Fernández-Celis, A.; Arrieta, V.; García-Peña, A.; Gainza, A.; Álvarez, V.; Sádaba, R.; et al. Targeting fatty acid-binding protein 4 improves pathologic features of aortic stenosis. Int. J. Mol. Sci. 2022, 23, 8439. [Google Scholar] [CrossRef] [PubMed]
- Roche-Molina, M.; Sanz-Rosa, D.; Cruz, F.M.; García-Prieto, J.; López, S.; Abia, R.; Muriana, F.J.; Fuster, V.; Ibáñez, B.; Bernal, J.A. Induction of sustained hypercholesterolemia by single adeno-associated virus-mediated gene transfer of mutant hPCSK9. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 50–59. [Google Scholar] [CrossRef]
- Rossner, P., Jr.; Sram, R.J. Immunochemical detection of oxidatively damaged DNA. Free Radic. Res. 2012, 46, 492–522. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Pan, Y.; Wang, X.; Ding, Y.; Zhou, C.; Shah, A.M.; Zhao, G.; Zhang, M. Celastrol Alleviates Aortic Valve Calcification via Inhibition of NADPH Oxidase 2 in Valvular Interstitial Cells. JACC Basic Transl. Sci. 2019, 5, 35–49. [Google Scholar] [CrossRef]
- Liberman, M.; Bassi, E.; Martinatti, M.K.; Lario, F.C.; Wosniak, J., Jr.; Pomerantzeff, P.M.; Laurindo, F.R. Oxidant generation predominates around calcifying foci and enhances progression of aortic valve calcification. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 463–470. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, F.; Xu, Y.; Sun, S.; Wang, H.; Du, Q.; Gu, C.; Black, S.M.; Han, Y.; Tang, H. Salusin-β Promotes Vascular Calcification via Nicotinamide Adenine Dinucleotide Phosphate/Reactive Oxygen Species-Mediated Klotho Downregulation. Antioxid. Redox Signal. 2019, 31, 1352–1370. [Google Scholar] [CrossRef]
- Valls-Lacalle, L.; Puertas-Umbert, L.; Varona, S.; Martínez-González, J.; Rodríguez, C.; Rodríguez-Sinovas, A. Human Lysyl Oxidase Over-Expression Enhances Baseline Cardiac Oxidative Stress but Does Not Aggravate ROS Generation or Infarct Size Following Myocardial Ischemia-Reperfusion. Antioxidants 2021, 11, 75. [Google Scholar] [CrossRef] [PubMed]
- Majora, M.; Wittkampf, T.; Schuermann, B.; Schneider, M.; Franke, S.; Grether-Beck, S.; Wilichowski, E.; Bernerd, F.; Schroeder, P.; Krutmann, J. Functional consequences of mitochondrial DNA deletions in human skin fibroblasts: Increased contractile strength in collagen lattices is due to oxidative stress-induced lysyl oxidase activity. Am. J. Pathol. 2009, 175, 1019–1029. [Google Scholar] [CrossRef]
- Ballester-Servera, C.; Alonso, J.; Taurón, M.; Rotllán, N.; Rodríguez, C.; Martínez-González, J. Lysyl oxidase expression in smooth muscle cells determines the level of intima calcification in hypercholesterolemia-induced atherosclerosis. Clin. Investig. Arterioscler 2024, S0214-9168(24)00007-X. [Google Scholar]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of oxidant stress that are clinically relevant in aging and age-related disease. Mech. Ageing Dev. 2013, 134, 139–157. [Google Scholar] [CrossRef] [PubMed]
- Kroese, L.J.; Scheffer, P.G. 8-hydroxy-2'-deoxyguanosine and cardiovascular disease: A systematic review. Curr. Atheroscler. Rep. 2014, 16, 452. [Google Scholar] [CrossRef]
- Martinet, W.; Knaapen, M.W.; De Meyer, G.R.; Herman, A.G.; Kockx, M.M. Elevated levels of oxidative DNA damage and DNA repair enzymes in human atherosclerotic plaques. Circulation 2002, 106, 927–932. [Google Scholar] [CrossRef]
- Hu, C.T.; Shao, Y.D.; Liu, Y.Z.; Xiao, X.; Cheng, Z.B.; Qu, S.L.; Huang, L.; Zhang, C. Oxidative stress in vascular calcification. Clin. Chim. Acta 2021, 519, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Xu, M.J.; Cai, Y.; Zhao, G.; Guan, Y.; Kong, W.; Tang, C.; Wang, X. Mitochondrial reactive oxygen species promote p65 nuclear translocation mediating high-phosphate-induced vascular calcification in vitro and in vivo. Kidney Int. 2011, 79, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Khan, K.; Hamid, Q.; Mardini, A.; Siddique, A.; Aguilar-Gonzalez, L.P.; Makhoul, G.; Alaws, H.; Genest, J.; Thanassoulis, G.; et al. Pathological significance of lipoprotein(a) in aortic valve stenosis. Atherosclerosis 2018, 272, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Yu, B.; Tardif, J.C.; Rhéaume, E.; Al-Kindi, H.; Filimon, S.; Pop, C.; Genest, J.; Cecere, R.; Schwertani, A. Significance of the Wnt signaling pathway in coronary artery atherosclerosis. Front. Cardiovasc. Med. 2024, 22, 1360380. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, G.; Chou, I.N.; Kagan, H.M. Hydrogen peroxide-mediated, lysyl oxidase-dependent chemotaxis of vascular smooth muscle cells. J. Cell. Biochem. 2000, 78, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.L.; Fogelgren, B.; Hess, A.R.; Seftor, E.A.; Wiley, E.L.; Fong, S.F.; Csiszar, K.; Hendrix, M.J.; Kirschmann, D.A. Lysyl oxidase regulates breast cancer cell migration and adhesion through a hydrogen peroxide-mediated mechanism. Cancer Res. 2005, 65, 11429–11436. [Google Scholar] [CrossRef]
- Byon, C.H.; Javed, A.; Dai, Q.; Kappes, J.C.; Clemens, T.L.; Darley-Usmar, V.M.; McDonald, J.M.; Chen, Y. Oxidative stress induces vascular calcification through modulation of the osteogenic transcription factor Runx2 by AKT signaling. J. Biol. Chem. 2008, 283, 15319–15327. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Chu, Y.; Brooks, R.M.; Richenbacher, W.E.; Peña-Silva, R.; Heistad, D.D. Dysregulation of antioxidant mechanisms contributes to increased oxidative stress in calcific aortic valvular stenosis in humans. J. Am. Coll. Cardiol. 2008, 52, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Branchetti, E.; Sainger, R.; Poggio, P.; Grau, J.B.; Patterson-Fortin, J.; Bavaria, J.E.; Chorny, M.; Lai, E.; Gorman, R.C.; Levy, R.J.; et al. Antioxidant enzymes reduce DNA damage and early activation of valvular interstitial cells in aortic valve sclerosis. Arter. Thromb. Vasc. Biol. 2013, 33, e66–e74. [Google Scholar] [CrossRef]
- Yamada, S.; Taniguchi, M.; Tokumoto, M.; Toyonaga, J.; Fujisaki, K.; Suehiro, T.; Noguchi, H.; Iida, M.; Tsuruya, K.; Kitazono, T. The antioxidant tempol ameliorates arterial medial calcification in uremic rats: Important role of oxidative stress in the pathogenesis of vascular calcification in chronic kidney disease. J. Bone Miner. Res. 2012, 27, 474–485. [Google Scholar] [CrossRef]
- Henze, L.A.; Luong, T.T.D.; Boehme, B.; Masyout, J.; Schneider, M.P.; Brachs, S.; Lang, F.; Pieske, B.; Pasch, A.; Eckardt, K.U.; et al. Impact of C-reactive protein on osteo-/chondrogenic transdifferentiation and calcification of vascular smooth muscle cells. Aging 2019, 11, 5445–5462. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Nguyen, T.T.; Da Ly, D.; Xia, J.B.; Qi, X.F.; Lee, I.K.; Cha, S.K.; Park, K.S. Oxidative stress by Ca2+ overload is critical for phosphate-induced vascular calcification. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H1302–H1312. [Google Scholar] [CrossRef] [PubMed]
- Sulistyowati, E.; Hsu, J.H.; Lee, S.J.; Huang, S.E.; Sihotang, W.Y.; Wu, B.N.; Dai, Z.K.; Lin, M.C.; Yeh, J.L. Potential Actions of Baicalein for Preventing Vascular Calcification of Smooth Muscle Cells In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 5673. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, X.; Liang, E.; Yang, R.; Liu, Y.; Liu, X.; Yan, F.; Xing, Y. Panax quinquefolius saponin inhibits vascular smooth muscle cell calcification via activation of nuclear factor-erythroid 2-related factor 2. BMC Complement. Med. Ther. 2023, 23, 129. [Google Scholar] [CrossRef] [PubMed]
- Zoubdane, N.; Abdo, R.A.; Nguyen, M.; Bentourkia, M.; Turcotte, E.E.; Berrougui, H.; Fulop, T.; Khalil, A. High Tyrosol and Hydroxytyrosol Intake Reduces Arterial Inflammation and Atherosclerotic Lesion Microcalcification in Healthy Older Populations. Antioxidants 2024, 13, 130. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, W.; Branchetti, E.; Grau, J.B.; Li, G.; Ayoub, S.; Lai, E.K.; Rioux, N.; Tovmasyan, A.; Fortier, J.H.; Sacks, M.S.; et al. Porphyrin-Based SOD Mimic MnTnBu OE -2-PyP5+ Inhibits Mechanisms of Aortic Valve Remodeling in Human and Murine Models of Aortic Valve Sclerosis. J. Am. Heart Assoc. 2018, 7, e007861. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, R.; Shiwakoti, S.; Ko, J.Y.; Dhakal, B.; Park, S.H.; Choi, I.J.; Kim, H.J.; Oak, M.H. Oxidative Stress in Calcific Aortic Valve Stenosis: Protective Role of Natural Antioxidants. Antioxidants 2022, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.T.; Yeh, H.Y.; Tsai, Y.T.; Chuang, P.H.; Yuan, T.H.; Huang, J.W.; Chen, H.W. Natural and non-natural antioxidative compounds: Potential candidates for treatment of vascular calcification. Cell Death Discov. 2019, 5, 145. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Elbatreek, M.H.; Pachado, M.P.; Cuadrado, A.; Jandeleit-Dahm, K.; Schmidt, H.H.H.W. Reactive Oxygen Comes of Age: Mechanism-Based Therapy of Diabetic End-Organ Damage. Trends Endocrinol. Metab. 2019, 30, 312–327. [Google Scholar] [CrossRef]
- Schmidt, H.H.; Stocker, R.; Vollbracht, C.; Paulsen, G.; Riley, D.; Daiber, A.; Cuadrado, A. Antioxidants in Translational Medicine. Antioxid. Redox Signal. 2015, 23, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yang, A.; Jia, J.; Popov, Y.V.; Schuppan, D.; You, H. Lysyl Oxidase (LOX) Family Members: Rationale and Their Potential as Therapeutic Targets for Liver Fibrosis. Hepatology 2020, 72, 729–741. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballester-Servera, C.; Alonso, J.; Cañes, L.; Vázquez-Sufuentes, P.; García-Redondo, A.B.; Rodríguez, C.; Martínez-González, J. Lysyl Oxidase in Ectopic Cardiovascular Calcification: Role of Oxidative Stress. Antioxidants 2024, 13, 523. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050523
Ballester-Servera C, Alonso J, Cañes L, Vázquez-Sufuentes P, García-Redondo AB, Rodríguez C, Martínez-González J. Lysyl Oxidase in Ectopic Cardiovascular Calcification: Role of Oxidative Stress. Antioxidants. 2024; 13(5):523. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050523
Chicago/Turabian StyleBallester-Servera, Carme, Judith Alonso, Laia Cañes, Paula Vázquez-Sufuentes, Ana B. García-Redondo, Cristina Rodríguez, and José Martínez-González. 2024. "Lysyl Oxidase in Ectopic Cardiovascular Calcification: Role of Oxidative Stress" Antioxidants 13, no. 5: 523. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox13050523