
Protective Effects of Long-Term Escitalopram Administration on Memory and Hippocampal BDNF and BCL-2 Gene Expressions in Rats Exposed to Predictable and Unpredictable Chronic Mild Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
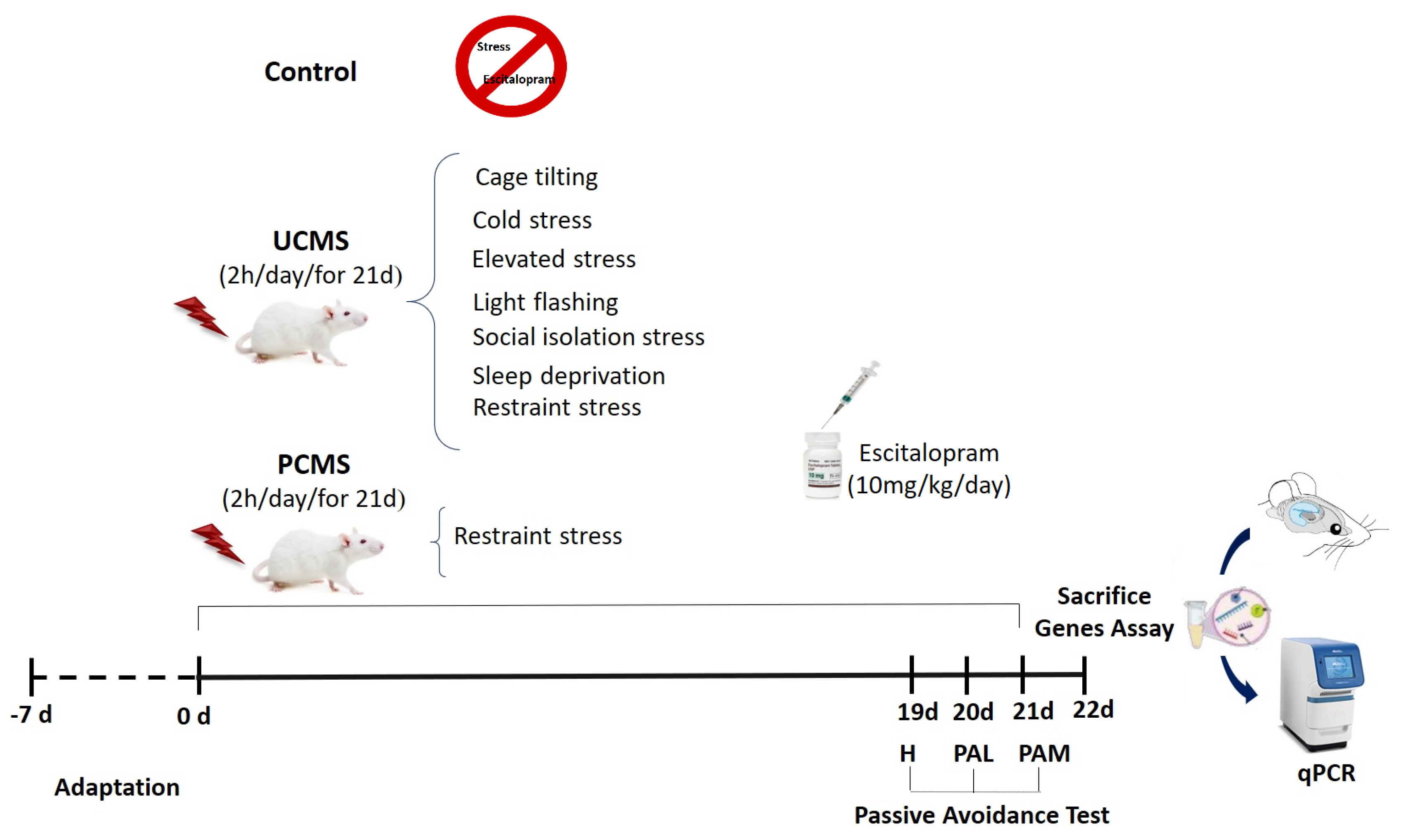
2.2. Stress Paradigm
2.3. Drug Treatment
2.4. Passive Avoidance Test
2.5. Gene Expression Assessment
2.6. Statistical Analysis
3. Results
3.1. Passive Avoidance Test
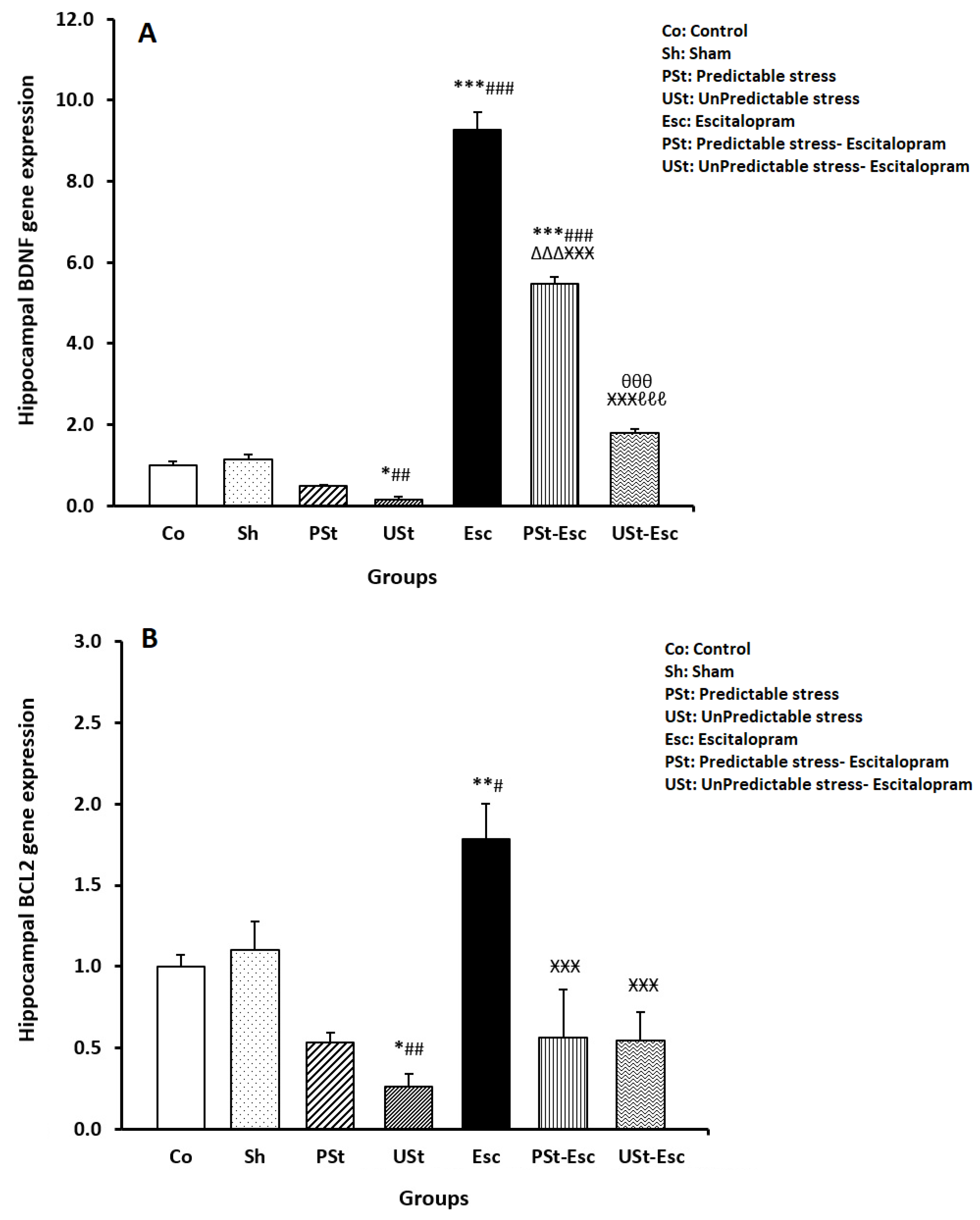
3.2. Hippocampal BDNF and BCL-2 Gene Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ortiz, J.B.; Conrad, C.D. The impact from the aftermath of chronic stress on hippocampal structure and function: Is there a recovery? Front. Neuroendocrinol. 2018, 49, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Algamal, M.; Pearson, A.J.; Hahn-Townsend, C.; Burca, I.; Mullan, M.; Crawford, F.; Ojo, J.O. Repeated unpredictable stress and social isolation induce chronic HPA axis dysfunction and persistent abnormal fear memory. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 104, 110035. [Google Scholar] [CrossRef] [PubMed]
- Zoeram, S.B.; Salmani, M.E.; Lashkarbolouki, T.; Goudarzi, I. Hippocampal orexin receptor blocking prevented the stress induced social learning and memory deficits. Neurobiol. Learn Mem. 2019, 157, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Leem, Y.H.; Park, J.S.; Chang, H.; Park, J.; Kim, H.S. Exercise Prevents Memory Consolidation Defects Via Enhancing Prolactin Responsiveness of CA1 Neurons in Mice Under Chronic Stress. Mol. Neurobiol. 2019, 56, 6609–6625. [Google Scholar] [CrossRef] [PubMed]
- Ionita, R.; Postu, P.A.; Mihasan, M.; Gorgan, D.L.; Hancianu, M.; Cioanca, O.; Hritcu, L. Ameliorative effects of Matricaria chamomilla L. hydroalcoholic extract on scopolamine-induced memory impairment in rats: A behavioral and molecular study. Phytomedicine 2018, 47, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-I.; Paulus, W.; Batsikadze, G.; Jamil, A.; Kuo, M.-F.; Nitsche, M.A. Chronic enhancement of serotonin facilitates excitatory transcranial direct current stimulation-induced neuroplasticity. Neuropsychopharmacology 2016, 41, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Gruca, P.; Lason, M.; Tota-Glowczyk, K.; Litwa, E.; Niemczyk, M.; Papp, M. Validation of chronic mild stress in the Wistar-Kyoto rat as an animal model of treatment-resistant depression. Behav. Pharmacol. 2019, 30, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Radahmadi, M.; Reisi, P. Effects of repeated treatment with cholecystokinin sulfated octapeptide on passive avoidance memory under chronic restraint stress in male rats. Adv. Biomed. Res. 2015, 4, 150. [Google Scholar]
- Kamińska, K.; Górska, A.; Noworyta-Sokołowska, K.; Wojtas, A.; Rogóż, Z.; Gołembiowska, K. The effect of chronic co-treatment with risperidone and novel antidepressant drugs on the dopamine and serotonin levels in the rats frontal cortex. Pharmacol. Rep. 2018, 70, 1023–1031. [Google Scholar] [CrossRef]
- Meneses, A.; Liy-Salmeron, G. Serotonin and emotion, learning and memory. Rev. Neurosci. 2012, 23, 543–553. [Google Scholar] [CrossRef]
- Lu, Q.H.; Mouri, A.; Yang, Y.; Kunisawa, K.; Teshigawara, T.; Hirakawa, M.; Mori, Y.; Yamamoto, Y.; Zou, L.B.; Nabeshima, T.; et al. Chronic unpredictable mild stress-induced behavioral changes are coupled with dopaminergic hyperfunction and serotonergic hypofunction in mouse models of depression. Behav. Brain Res. 2019, 372, 112053. [Google Scholar] [CrossRef]
- Dale, E.; Pehrson, A.L.; Jeyarajah, T.; Li, Y.; Leiser, S.C.; Smagin, G.; Olsen, C.K.; Sanchez, C. Effects of serotonin in the hippocampus: How SSRIs and multimodal antidepressants might regulate pyramidal cell function. CNS Spectr. 2016, 21, 143–161. [Google Scholar] [CrossRef]
- Skandali, N.; Rowe, J.B.; Voon, V.; Deakin, J.B.; Cardinal, R.N.; Cormack, F.; Passamonti, L.; Bevan-Jones, W.R.; Regenthal, R.; Chamberlain, S.R. Dissociable effects of acute SSRI (escitalopram) on executive, learning and emotional functions in healthy humans. Neuropsychopharmacology 2018, 43, 2645–2651. [Google Scholar] [CrossRef]
- Reed, M.B.; Vanicek, T.; Seiger, R.; Klöbl, M.; Spurny, B.; Handschuh, P.; Ritter, V.; Unterholzner, J.; Godbersen, G.M.; Gryglewski, G. Neuroplastic effects of a selective serotonin reuptake inhibitor in relearning and retrieval. Neuroimage 2021, 236, 118039. [Google Scholar] [CrossRef]
- Ren, Q.-G.; Wang, Y.-J.; Gong, W.-G.; Xu, L.; Zhang, Z.-J. Escitalopram ameliorates Tau hyperphosphorylation and spatial memory deficits induced by protein kinase A activation in sprague dawley rats. J. Alzheimers Dis. 2015, 47, 61–71. [Google Scholar] [CrossRef]
- Montagrin, A.; Saiote, C.; Schiller, D. The social hippocampus. Hippocampus 2018, 28, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Benatti, C.; Alboni, S.; Blom, J.M.C.; Mendlewicz, J.; Tascedda, F.; Brunello, N. Molecular changes associated with escitalopram response in a stress-based model of depression. Psychoneuroendocrinology 2018, 87, 74–82. [Google Scholar] [CrossRef]
- Bartsch, J.C.; Von Cramon, M.; Gruber, D.; Heinemann, U.; Behr, J. Stress-induced enhanced long-term potentiation and reduced threshold for N-methyl-D-aspartate receptor-and β-adrenergic receptor-mediated synaptic plasticity in rodent ventral subiculum. Front. Mol. Neurosci. 2021, 14, 658465. [Google Scholar] [CrossRef]
- Karbowski, J. Metabolic constraints on synaptic learning and memory. J. Neurophysiol. 2019, 122, 1473–1490. [Google Scholar] [CrossRef]
- Seo, M.K.; Lee, J.G.; Park, S.W. Effects of escitalopram and ibuprofen on a depression-like phenotype induced by chronic stress in rats. Neurosci. Lett. 2019, 696, 168–173. [Google Scholar] [CrossRef]
- Dionisie, V.; Ciobanu, A.M.; Toma, V.A.; Manea, M.C.; Baldea, I.; Olteanu, D.; Sevastre-Berghian, A.; Clichici, S.; Manea, M.; Riga, S. Escitalopram targets oxidative stress, caspase-3, BDNF and MeCP2 in the hippocampus and frontal cortex of a rat model of depression induced by chronic unpredictable mild stress. Int. J. Mol. Sci. 2021, 22, 7483. [Google Scholar] [CrossRef]
- Asl, S.S.; Farhadi, M.H.; Moosavizadeh, K.; Saraei, A.S.K.; Soleimani, M.; Jamei, S.B.; Joghataei, M.T.; Samzadeh-Kermani, A.; Hashemi-Nasl, H.; Mehdizadeh, M. Evaluation of Bcl-2 family gene expression in hippocampus of 3, 4-methylenedioxymethamphetamine treated rats. Cell J. 2012, 13, 275. [Google Scholar]
- Koohpar, Z.K.; Hashemi, M.; Mahdian, R.; Parivar, K. The effect of pentoxifylline on bcl-2 gene expression changes in hippocampus after long-term use of ecstasy in wistar rats. Iran J. Pharm. Res. 2013, 12, 521. [Google Scholar]
- Sheikh, A.M.; Malik, M.; Wen, G.; Chauhan, A.; Chauhan, V.; Gong, C.X.; Liu, F.; Brown, W.T.; Li, X. BDNF-Akt-Bcl2 antiapoptotic signaling pathway is compromised in the brain of autistic subjects. J. Neurosci. Res. 2010, 88, 2641–2647. [Google Scholar] [CrossRef] [PubMed]
- Eidelkhani, N.; Radahmadi, M.; Kazemi, M.; Rafiee, L.; Alaei, H.; Reisi, P. Effects of doxepin on brain-derived neurotrophic factor, tumor necrosis factor alpha, mitogen-activated protein kinase 14, and AKT1 genes expression in rat hippocampus. Adv. Biomed. Res. 2015, 4, 203. [Google Scholar]
- Dastgerdi, H.H.; Radahmadi, M.; Reisi, P. Comparative study of the protective effects of crocin and exercise on long-term potentiation of CA1 in rats under chronic unpredictable stress. Life Sci. 2020, 256, 118018. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Rodrigues, A.J.; Silva, J.M.; Tronche, F.; Almeida, O.F.X.; Sousa, N.; Sotiropoulos, I. Chronic Stress and Glucocorticoids: From Neuronal Plasticity to Neurodegeneration. Neural Plast 2016, 2016, 6391686. [Google Scholar] [CrossRef] [PubMed]
- Zelada, M.I.; Garrido, V.; Liberona, A.; Jones, N.; Zúñiga, K.; Silva, H.; Nieto, R.R. Brain-Derived Neurotrophic Factor (BDNF) as a Predictor of Treatment Response in Major Depressive Disorder (MDD): A Systematic Review. Int. J. Mol. Sci. 2023, 24, 14810. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xie, K.; Yang, X.; Gu, J.; Ge, L.; Wang, X.; Wang, Z. Resveratrol reverses the effects of chronic unpredictable mild stress on behavior, serum corticosterone levels and BDNF expression in rats. Behav. Brain Res. 2014, 264, 9–16. [Google Scholar] [CrossRef]
- Wang, X.; Xie, Y.; Zhang, T.; Bo, S.; Bai, X.; Liu, H.; Li, T.; Liu, S.; Zhou, Y.; Cong, X. Resveratrol reverses chronic restraint stress-induced depression-like behaviour: Involvement of BDNF level, ERK phosphorylation and expression of Bcl-2 and Bax in rats. Brain Res. Bull. 2016, 125, 134–143. [Google Scholar] [CrossRef]
- Sikora, M.; Konopelski, P.; Pham, K.; Wyczalkowska-Tomasik, A.; Ufnal, M. Repeated restraint stress produces acute and chronic changes in hemodynamic parameters in rats. Stress 2016, 19, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Bondi, C.O.; Rodriguez, G.; Gould, G.G.; Frazer, A.; Morilak, D.A. Chronic unpredictable stress induces a cognitive deficit and anxiety-like behavior in rats that is prevented by chronic antidepressant drug treatment. Neuropsychopharmacology 2008, 33, 320–331. [Google Scholar] [CrossRef] [PubMed]
- El Marzouki, H.; Aboussaleh, Y.; Najimi, M.; Chigr, F.; Ahami, A. Effect of Cold Stress on Neurobehavioral and Physiological Parameters in Rats. Front. Physiol. 2021, 12, 660124. [Google Scholar] [CrossRef] [PubMed]
- Yarnell, A.M.; Grunberg, N. A neurobehavioral phenotype of blast traumatic brain injury and psychological stress in male and female rats. In Proceedings of the Society for Neuroscience Meeting, New Orleans, LA, USA, 13-17 October 2012. [Google Scholar]
- Hosseini Dastgerdi, A.; Radahmadi, M.; Pourshanazari, A.A. Comparing the effects of crocin at different doses on excitability and long-term potentiation in the CA1 area, as well as the electroencephalogram responses of rats under chronic stress. Metab. Brain Dis. 2021, 36, 1879–1887. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.; Ma, P.; Yang, H.; Xiong, H.; Wang, M.; Peng, C.; Tu, P.; Li, X. Antidepressant-like effects of Cistanche tubulosa extract on chronic unpredictable stress rats through restoration of gut microbiota homeostasis. Front. Pharmacol. 2018, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Fabries, P.; Gomez-Merino, D.; Sauvet, F.; Malgoyre, A.; Koulmann, N.; Chennaoui, M. Sleep loss effects on physiological and cognitive responses to systemic environmental hypoxia. Front. Physiol. 2022, 13, 1046166. [Google Scholar] [CrossRef] [PubMed]
- Rowson, S.A.; Harrell, C.S.; Bekhbat, M.; Gangavelli, A.; Wu, M.J.; Kelly, S.D.; Reddy, R.; Neigh, G.N. Neuroinflammation and behavior in HIV-1 transgenic rats exposed to chronic adolescent stress. Front. Psychiatry 2016, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Farahbakhsh, Z.; Radahmadi, M. The protective effects of escitalopram on chronic restraint stress-induced memory deficits in adult rats. Physiol. Pharmacol. 2022, 26, 39–48. [Google Scholar] [CrossRef]
- Kaminska, K.; Rogoz, Z. The antidepressant- and anxiolytic-like effects following co-treatment with escitalopram and risperidone in rats. J. Physiol. Pharmacol. 2016, 67, 471–480. [Google Scholar]
- Jastrzębska, J.; Frankowska, M.; Suder, A.; Wydra, K.; Nowak, E.; Filip, M.; Przegaliński, E. Effects of escitalopram and imipramine on cocaine reinforcement and drug-seeking behaviors in a rat model of depression. Brain Res. 2017, 1673, 30–41. [Google Scholar] [CrossRef]
- Yang, S.N.; Wang, Y.H.; Tung, C.S.; Ko, C.Y.; Liu, Y.P. Effects of Escitalopram on a Rat Model of Persistent Stress-Altered Hedonic Activities: Towards a New Understanding of Stress and Depression. Chin. J. Physiol. 2015, 58, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Kalantarzadeh, E.; Radahmadi, M.; Reisi, P. Effects of different dark chocolate diets on memory functions and brain corticosterone levels in rats under chronic stress. Physiol. Pharmacol. 2020, 24, 185–196. [Google Scholar] [CrossRef]
- Salehifard, K.; Radahmadi, M.; Reisi, P. The effect of photoperiodic stress on anxiety-like behaviors, learning, memory, locomotor activity and memory consolidation in rats. Physiol. Pharmacol. 2023, 27, 244–253. [Google Scholar] [CrossRef]
- Shabani, M.; Divsalar, K.; Janahmadi, M. Destructive effects of prenatal WIN 55212-2 exposure on central nervous system of neonatal rats. Addict. Health 2012, 4, 9–19. [Google Scholar]
- Reisi, P.; Eidelkhani, N.; Rafiee, L.; Kazemi, M.; Radahmadi, M.; Alaei, H. Effects of doxepin on gene expressions of Bcl-2 family, TNF-α, MAP kinase 14, and Akt1 in the hippocampus of rats exposed to stress. Res. Pharm. Sci. 2017, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Roustazade, R.; Radahmadi, M.; Yazdani, Y. Therapeutic effects of saffron extract on different memory types, anxiety, and hippocampal BDNF and TNF-α gene expressions in sub-chronically stressed rats. Nutr. Neurosci. 2022, 25, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Radahmadi, M.; Hosseini Dastgerdi, A.; Fallah, N.; Alaei, H. The effects of acute, sub-chronic and chronic psychical stress on the brain electrical activity in male rats. Physiol. Pharmacol. 2017, 21, 185–192. [Google Scholar]
- Sharpe, M.J.; Marchant, N.J.; Whitaker, L.R.; Richie, C.T.; Zhang, Y.J.; Campbell, E.J.; Koivula, P.P.; Necarsulmer, J.C.; Mejias-Aponte, C.; Morales, M. Lateral hypothalamic GABAergic neurons encode reward predictions that are relayed to the ventral tegmental area to regulate learning. Curr. Biol. 2017, 27, 2089–2100.e2085. [Google Scholar] [CrossRef] [PubMed]
- Gulyaeva, N. Glucocorticoid regulation of the glutamatergic synapse: Mechanisms of stress-dependent neuroplasticity. J. Evol. Biochem. Physiol. 2021, 57, 564–576. [Google Scholar] [CrossRef]
- Bhagya, V.; Srikumar, B.; Raju, T.; Shankaranarayana Rao, B. Chronic escitalopram treatment restores spatial learning, monoamine levels, and hippocampal long-term potentiation in an animal model of depression. Psychopharmacology 2011, 214, 477–494. [Google Scholar] [CrossRef]
- Ceglia, I.; Acconcia, S.; Fracasso, C.; Colovic, M.; Caccia, S.; Invernizzi, R. Effects of chronic treatment with escitalopram or citalopram on extracellular 5-HT in the prefrontal cortex of rats: Role of 5-HT1A receptors. Br. J. Pharmacol. 2004, 142, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Drozd, R.; Rychlik, M.; Fijalkowska, A.; Rygula, R. Effects of cognitive judgement bias and acute antidepressant treatment on sensitivity to feedback and cognitive flexibility in the rat version of the probabilistic reversal-learning test. Behav. Brain Res. 2019, 359, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Sardari, M.; Rezayof, A.; Zarrindast, M.-R. 5-HT1A receptor blockade targeting the basolateral amygdala improved stress-induced impairment of memory consolidation and retrieval in rats. Neuroscience 2015, 300, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Pinzón-Parra, C.; Vidal-Jiménez, B.; Camacho-Abrego, I.; Flores-Gómez, A.A.; Rodríguez-Moreno, A.; Flores, G. Juvenile stress causes reduced locomotor behavior and dendritic spine density in the prefrontal cortex and basolateral amygdala in Sprague–Dawley rats. Synapse 2019, 73, e22066. [Google Scholar] [CrossRef] [PubMed]
- Dastgerdi, H.H.; Radahmadi, M.; Reisi, P.; Dastgerdi, A.H. Effect of Crocin, Exercise, and Crocin-accompanied Exercise on Learning and Memory in Rats under Chronic Unpredictable Stress. Adv. Biomed. Res. 2018, 7, 137. [Google Scholar]
- Ma, L.; Lu, Z.-n.; Hu, P.; Yao, C.-j. Neuroprotective effect of escitalopram oxalate in rats with chronic hypoperfusion. J. Huazhong Univ. Sci. Technol. 2015, 35, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.R.; Murphy, T.; De Jong, S.; Lee, S.H.; Tansey, K.E.; Hodgson, K.; Uher, R.; Price, J.; Thuret, S.; Breen, G. The genome-wide expression effects of escitalopram and its relationship to neurogenesis, hippocampal volume, and antidepressant response. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2017, 174, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.B.; du Jardin, K.G.; Song, D.; Budac, D.; Smagin, G.; Sanchez, C.; Pehrson, A.L. Vortioxetine, but not escitalopram or duloxetine, reverses memory impairment induced by central 5-HT depletion in rats: Evidence for direct 5-HT receptor modulation. Eur. Neuropsychopharmacol. 2014, 24, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Zamani, M.; Radahmadi, M.; Reisi, P. Therapeutic effects of exercise, escitalopram and exercise-accompanied escitalopram on brain functions in rats with depression. Physiol. Pharmacol. 2022, 26, 188–199. [Google Scholar] [CrossRef]
- Gammoh, O.; Mayyas, F.; Darwish Elhajji, F. Chlorpheniramine and escitalopram: Similar antidepressant and nitric oxide lowering roles in a mouse model of anxiety. Biomed. Rep. 2017, 6, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Joodaki, M.; Radahmadi, M.; Alaei, H. Comparing the Therapeutic Effects of Crocin, Escitalopram and Co-Administration of Escitalopram and Crocin on Learning and Memory in Rats with Stress-Induced Depression. Malays J. Med. Sci. 2021, 28, 50. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Tung, C.-S.; Liu, Y.-P. Escitalopram reversed the traumatic stress-induced depressed and anxiety-like symptoms but not the deficits of fear memory. Psychopharmacology 2016, 233, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yi, L.; Xia, X.; Chen, X.; Hou, X.; Zhang, L.; Yang, F.; Liao, J.; Han, Z.; Fu, Y. Transcriptome comparative analysis of amygdala-hippocampus in depression: A rat model induced by chronic unpredictable mild stress (CUMS). J. Affect Disord. 2023, 334, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Minami, S.; Satoyoshi, H.; Ide, S.; Inoue, T.; Yoshioka, M.; Minami, M. Suppression of reward-induced dopamine release in the nucleus accumbens in animal models of depression: Differential responses to drug treatment. Neurosci. Lett. 2017, 650, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Salari, M.; Eftekhar-Vaghefi, S.H.; Asadi-Shekaari, M.; Esmaeilpour, K.; Solhjou, S.; Amiri, M.; Ahmadi-Zeidabadi, M. Impact of ketamine administration on chronic unpredictable stress-induced rat model of depression during extremely low-frequency electromagnetic field exposure: Behavioral, histological and molecular study. Brain Behav. 2023, 13, e2986. [Google Scholar] [CrossRef] [PubMed]
- Scotton, E.; Colombo, R.; Reis, J.C.; Possebon, G.M.; Hizo, G.H.; Valiati, F.E.; Gea, L.P.; Bristot, G.; Salvador, M.; Silva, T.M. BDNF prevents central oxidative damage in a chronic unpredictable mild stress model: The possible role of PRDX-1 in anhedonic behavior. Behav. Brain Res. 2020, 378, 112245. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, N.; Jain, V.; Deep, S.; Prasad, D.; Singh, S.B.; Khan, N. Neuroprotective role of intermittent hypobaric hypoxia in unpredictable chronic mild stress induced depression in rats. PLoS ONE 2016, 11, e0149309. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, N.; Dey, S.K.; Chakravarty, S.; Kumar, A. miR-30 Family miRNAs Mediate the Effect of Chronic Social Defeat Stress on Hippocampal Neurogenesis in Mouse Depression Model. Front. Mol. Neurosci. 2019, 12, 465796. [Google Scholar] [CrossRef]
- Carneiro, C.A.; Santos, A.M.F.; Guedes, É.C.; Cavalcante, I.L.; Santos, S.G.; Oliveira, A.M.F.; Barbosa, F.F.; Silva, M.S.; Almeida, R.N.; Salvadori, M.S. Unpredictable subchronic stress induces depressive-like behavior: Behavioral and neurochemical evidences. Psychol. Neurosci. 2022, 15, 236. [Google Scholar] [CrossRef]
- Chai, H.H.; Fu, X.C.; Ma, L.; Sun, H.T.; Chen, G.Z.; Song, M.Y.; Chen, W.X.; Chen, Y.S.; Tan, M.X.; Guo, Y.W.; et al. The chemokine CXCL1 and its receptor CXCR2 contribute to chronic stress-induced depression in mice. FASEB J. 2019, 33, 8853–8864. [Google Scholar] [CrossRef] [PubMed]
- Kosten, T.A.; Galloway, M.P.; Duman, R.S.; Russell, D.S.; D’sa, C. Repeated unpredictable stress and antidepressants differentially regulate expression of the bcl-2 family of apoptotic genes in rat cortical, hippocampal, and limbic brain structures. Neuropsychopharmacology 2008, 33, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, H.; Radahmadi, M.; Alaei, H.; Reisi, P.; Karimi, S. The effect of basolateral amygdala nucleus lesion on memory under acute, mid and chronic stress in male rats. Turk J. Med. Sci. 2016, 46, 1915–1925. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.N.; El-Aidi, A.A.; Ali, M.M.; Attia, Y.M.; Rashed, L.A. Modification of hippocampal markers of synaptic plasticity by memantine in animal models of acute and repeated restraint stress: Implications for memory and behavior. Neuromolecular Med. 2015, 17, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, A.; Gavini, K.; Yang, E.; Lyman-Henley, L.; Parameshwaran, K. Chronic traumatic stress impairs memory in mice: Potential roles of acetylcholine, neuroinflammation and corticotropin releasing factor expression in the hippocampus. Behav. Brain Res. 2017, 335, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sun, G.Q.; Yang, F.; Guan, Z.W.; Zhang, Z.; Zhao, J.; Liu, Y.Y.; Chu, L.; Pei, L. Baicalin regulates depression behavior in mice exposed to chronic mild stress via the Rac/LIMK/cofilin pathway. Biomed. Pharmacother. 2019, 116, 109054. [Google Scholar] [CrossRef] [PubMed]
- Dvojkovic, A.; Perkovic, M.N.; Sagud, M.; Erjavec, G.N.; Peles, A.M.; Strac, D.S.; Cusa, B.V.; Tudor, L.; Kusevic, Z.; Konjevod, M. Effect of vortioxetine vs. escitalopram on plasma BDNF and platelet serotonin in depressed patients. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110016. [Google Scholar] [CrossRef] [PubMed]
- Mayegowda, S.B.; Rao, B.V.; Rao, B.S.R.S. Escitalopram treatment ameliorates chronic immobilization stress-induced depressive behavior and cognitive deficits by modulating BDNF expression in the hippocampus. J. Appl. Pharm. Sci. 2024, 14, 170–182. [Google Scholar] [CrossRef]
- Shen, J.; Qu, C.; Xu, L.; Sun, H.; Zhang, J. Resveratrol exerts a protective effect in chronic unpredictable mild stress–induced depressive-like behavior: Involvement of the AKT/GSK3β signaling pathway in hippocampus. Psychopharmacology 2019, 236, 591–602. [Google Scholar] [CrossRef]
- Saad, M.A.; El-Sahar, A.E.; Sayed, R.H.; Elbaz, E.M.; Helmy, H.S.; Senousy, M.A. Venlafaxine mitigates depressive-like behavior in ovariectomized rats by activating the EPO/EPOR/JAK2 signaling pathway and increasing the serum estradiol level. Neurotherapeutics 2019, 16, 404–415. [Google Scholar] [CrossRef]
- Henn, L.; Zanta, N.C.; Girardi, C.E.N.; Suchecki, D. Chronic Escitalopram Treatment Does Not Alter the Effects of Neonatal Stress on Hippocampal BDNF Levels, 5-HT 1A Expression and Emotional Behaviour of Male and Female Adolescent Rats. Mol. Neurobiol. 2021, 58, 926–943. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′ to 3′) | |
|---|---|---|
| BCL-2 | Forward | TAACGGAGGCTGGGATGC |
| Reverse | TGAGCAGCGTCTTCAGAGA | |
| BDNF | Forward | AGAATGAGGGCGTTTGCGTA |
| Reverse | CCTGGTGGAACATTGTGGCT | |
| ACTB | Forward | AGGCCCCTCTGAACCCTAAG |
| Reverse | CCAGAGGCATACAGGGACAA | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saedi Marghmaleki, V.; Radahmadi, M.; Alaei, H.; Khanahmad, H. Protective Effects of Long-Term Escitalopram Administration on Memory and Hippocampal BDNF and BCL-2 Gene Expressions in Rats Exposed to Predictable and Unpredictable Chronic Mild Stress. Brain Sci. 2024, 14, 420. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci14050420
Saedi Marghmaleki V, Radahmadi M, Alaei H, Khanahmad H. Protective Effects of Long-Term Escitalopram Administration on Memory and Hippocampal BDNF and BCL-2 Gene Expressions in Rats Exposed to Predictable and Unpredictable Chronic Mild Stress. Brain Sciences. 2024; 14(5):420. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci14050420
Chicago/Turabian StyleSaedi Marghmaleki, Vajihe, Maryam Radahmadi, Hojjatallah Alaei, and Hossein Khanahmad. 2024. "Protective Effects of Long-Term Escitalopram Administration on Memory and Hippocampal BDNF and BCL-2 Gene Expressions in Rats Exposed to Predictable and Unpredictable Chronic Mild Stress" Brain Sciences 14, no. 5: 420. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci14050420