
Temporal Dynamics of Fungal Communities in Alkali-Treated Round Bamboo Deterioration under Natural Weathering
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampling
2.2. Fungal Community Analysis
2.2.1. DNA Extraction and Sequencing
2.2.2. Sequence Data Processing
2.2.3. Statistical Analysis
2.3. Characteristics of Bamboo
2.3.1. Chemical Composition Determination
2.3.2. XPS Analysis
2.3.3. SEM Observation
2.3.4. Contact Angle Measurements
3. Results and Discussion
3.1. Variations in the Diversity of the Fungal Community
3.1.1. Alpha Diversity Analysis
3.1.2. Beta Diversity Analysis
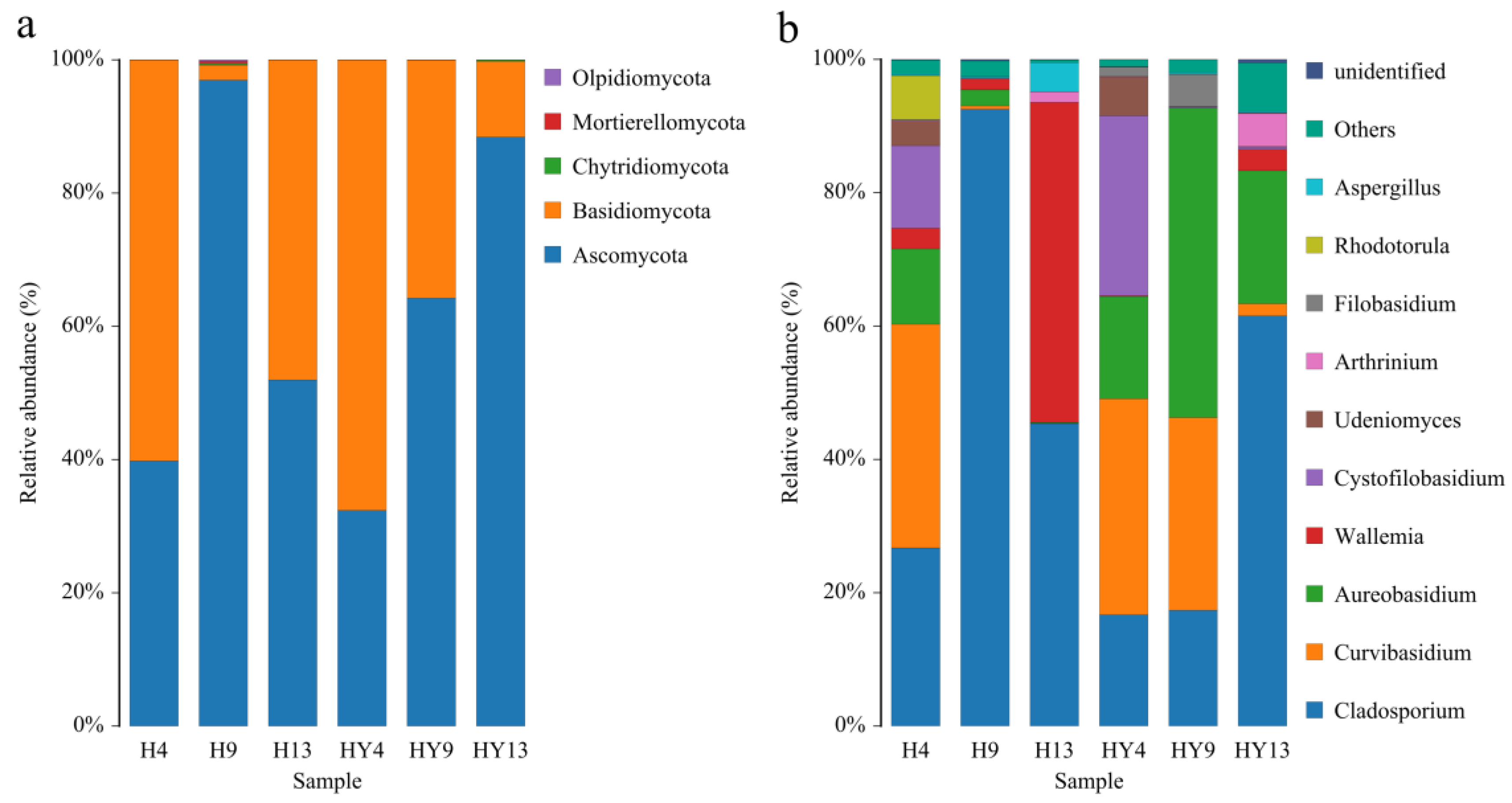
3.2. Microbial Community Composition
3.3. The Correlation between Environmental Factors and Fungal Communities
3.4. Function of Fungi
3.5. Chemical Composition Determination
3.6. SEM Analysis
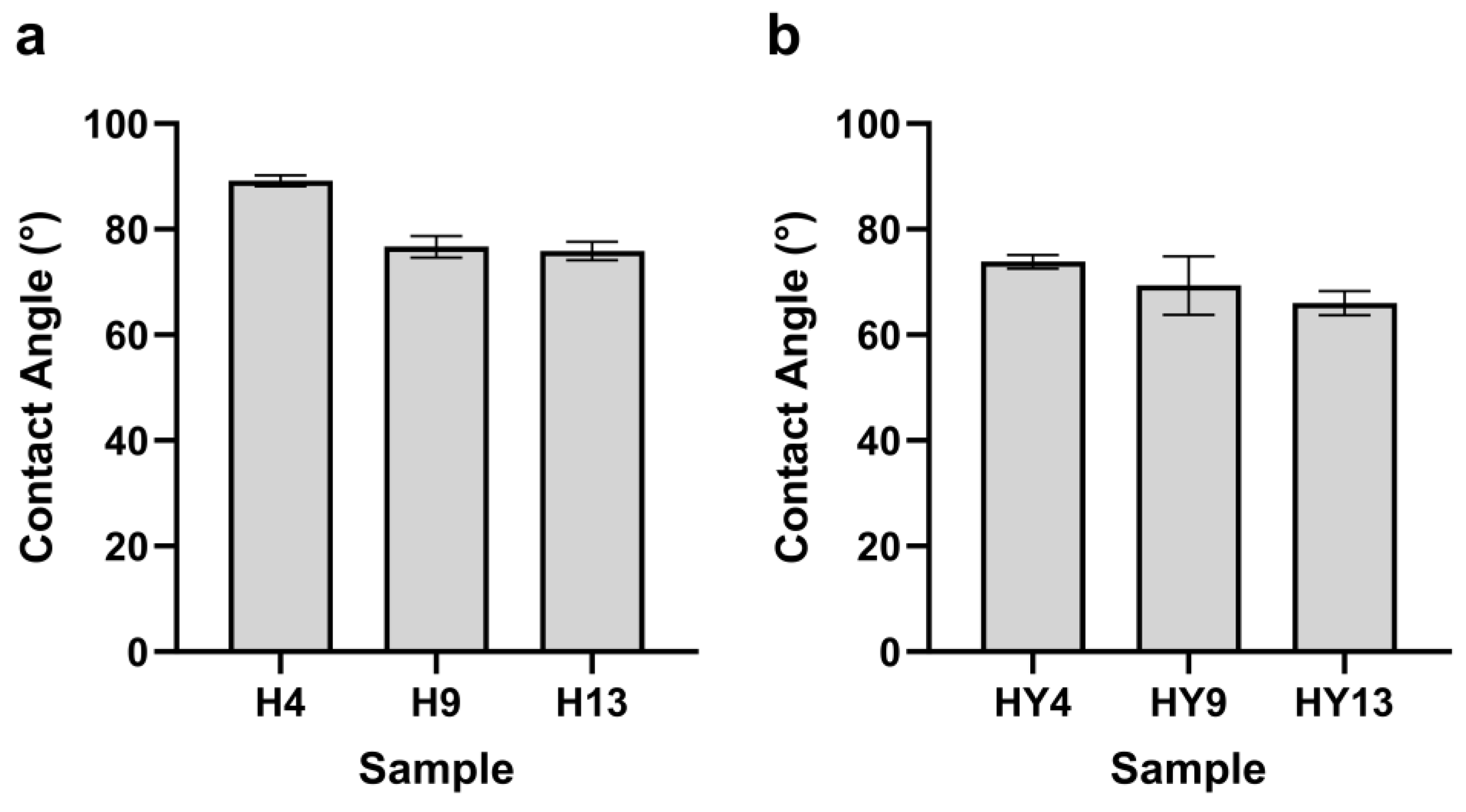
3.7. Contact Angle
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Archila, H.; Kaminski, S.; Trujillo, D.; Zea Escamilla, E.; Harries, K.A.J.M. Bamboo reinforced concrete: A critical review. Mater. Struct. 2018, 51, 102. [Google Scholar] [CrossRef]
- Fei, B.; Gao, Z.; Wang, J.; Liu, Z. Chapter 14—Biological, Anatomical, and Chemical Characteristics of Bamboo. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 283–306. [Google Scholar]
- Su, N.; Fang, C.; Yu, Z.; Zhou, H.; Wang, X.; Tang, T.; Zhang, S.; Fei, B. Effects of rosin treatment on hygroscopicity, dimensional stability, and pore structure of round bamboo culm. Constr. Build. Mater. 2021, 287, 123037. [Google Scholar] [CrossRef]
- Simi, A.; Shirehjini, R.M.; Shi, D.; Demartino, C.; Li, Z. A Review on Round Bamboo Structural Applications and Perspectives. In 2019 International Bamboo Construction Competition; Springer: Cham, Switzerland, 2022; pp. 29–40. [Google Scholar]
- Lv, H.F.; Chen, X.F.; Liu, X.M.; Fang, C.H.; Liu, H.R.; Zhang, B.; Fei, B.H. The vacuum-assisted microwave drying of round bamboos: Drying kinetics, color and mechanical property. Mater. Lett. 2018, 223, 159–162. [Google Scholar] [CrossRef]
- Rao, J.; Jiang, J.; Prosper, N.K.; Yang, X.S.; Liu, T.S.; Cai, W.; Wang, H.; Sun, F.L. Combination of polyethylene glycol impregnation and paraffin heat treatment to protect round bamboo from cracking. R. Soc. Open Sci. 2019, 6, 190105. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Li, Z.; Liu, K. Modern Engineered Bamboo Structures. In Proceedings of the Third International Conference on Modern Bamboo Structures (ICBS 2018), Beijing, China, 25–27 June 2018; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Okahisa, Y.; Yoshimura, T.; Imamura, Y.J. Seasonal and height-dependent fluctuation of starch and free glucose contents in moso bamboo (Phyllostachys pubescens) and its relation to attack by termites and decay fungi. J. Wood Sci. 2006, 52, 445–451. [Google Scholar] [CrossRef]
- Möller, R.; Mild, G. Protection of Moso bamboo (Phyllostachys pubescens) materials against fungal decay and discolouration by treatment with wood preservatives. Eur. J. Wood Wood Prod. 2019, 77, 139–145. [Google Scholar] [CrossRef]
- Tang, T.; Schmidt, O.; Liese, W. Environment-friendly short-term protection of bamboo against molding. J. Timber Dev. Assoc. India 2009, 55, 8–17. [Google Scholar]
- Ma, X.; Jiang, M.; Lu, W.; Qin, D. Study on the biological characteristics of stain fungi and mould fungi on bamboo wood. For. Res. 2009, 22, 819–823. [Google Scholar]
- Liu, Y.D. Bamboo timber mildew and anti-mold technology. Adv. Eng. Forum. 2012, 4, 139–144. [Google Scholar] [CrossRef]
- Borchers, A.T.; Chang, C.; Eric Gershwin, M. Mold and human health: A reality check. Clin. Rev. Allergy Immunol. 2017, 52, 305–322. [Google Scholar] [CrossRef]
- Schmidt, O.; Wei, D.; Tang, T.; Liese, W. Bamboo and fungi. J. Bamboo Ratt. 2013, 12, 1–14. [Google Scholar]
- Schmidt, O.; Wei, D.; Bahmani, M.; Tang, T.K.H.; Liese, W. Pilzbefall und Schutz von Bambushalmen und Palmenholz–eine Übersicht. Z. Für Mykol. 2015, 81, 1. [Google Scholar]
- Saharia, D.; Sarma, T. Screening of macro-fungi responsible for post harvest decay of bamboo culms in storage. Bioscan 2012, 7, 95–99. [Google Scholar]
- Hamid, N.H.; Sulaiman, O.; Mohammad, A.; Ahmad Ludin, N. The decay resistance and hyphae penetration of bamboo Gigantochloa scortechinii decayed by white and brown rot fungi. Int. J. For. Res. 2012, 2012, 572903. [Google Scholar]
- Schmidt, G.; Stute, T.; Lenz, M.T.; Melcher, E.; Ressel, J.B. Fungal deterioration of a novel scrimber composite made from industrially heat treated African highland bamboo. Ind. Crop Prod. 2020, 147, 112225. [Google Scholar] [CrossRef]
- Sulaiman, O.; Murphy, R. Ultrastructure of soft rot decay in bamboo cell walls. Mater. Und Org. 1995, 29, 243–251. [Google Scholar]
- Ladau, J.; Eloe-Fadrosh, E.A. Spatial, Temporal, and Phylogenetic Scales of Microbial Ecology. Trends Microbiol. 2019, 27, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.A.; Miyanishi, K.J. Testing the assumptions of chronosequences in succession. Ecol. Lett. 2008, 11, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, J.L.; Powell, R.D.; Nott, M.P.; Pimm, S.L. Assembling ecological communities in time and space. Oikos 1997, 80, 549–553. [Google Scholar] [CrossRef]
- Hannula, S.E.; Kielak, A.M.; Steinauer, K.; Huberty, M.; Jongen, R.; De Long, J.R.; Heinen, R.; Bezemer, T.M. Time after Time: Temporal Variation in the Effects of Grass and Forb Species on Soil Bacterial and Fungal Communities. Mbio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Gevers, D.; Cohan, F.M.; Lawrence, J.G.; Spratt, B.G.; Coenye, T.; Feil, E.J.; Stackebrandt, E.; de Peer, Y.V.; Vandamme, P.; Thompson, F.L. Re-evaluating prokaryotic species. Nat. Rev. Microbiol. 2005, 3, 733–739. [Google Scholar] [CrossRef]
- Vos, M. A species concept for bacteria based on adaptive divergence. Trends Microbiol. 2011, 19, 1–7. [Google Scholar] [CrossRef]
- Zhou, D.; Hyde, K.D. Fungal succession on bamboo in Hong Kong. Fungal Divers. 2002, 10, 213–227. [Google Scholar]
- Nakasaki, K.; Hirai, H.; Mimoto, H.; Quyen, T.N.M.; Koyama, M.; Takeda, K. Succession of microbial community during vigorous organic matter degradation in the primary fermentation stage of food waste composting. Sci. Total Environ. 2019, 671, 1237–1244. [Google Scholar] [CrossRef]
- Gonzalez, A.; King, A.; Robeson, M.S., II; Song, S.; Shade, A.; Metcalf, J.L.; Knight, R. Characterizing microbial communities through space and time. Curr. Opin. Biotechnol. 2012, 23, 431–436. [Google Scholar] [CrossRef]
- An, X.J.; Han, S.B.; Ren, X.; Sichone, J.; Fan, Z.W.; Wu, X.X.; Zhang, Y.; Wang, H.; Cai, W.; Sun, F.L. Succession of Fungal Community during Outdoor Deterioration of Round Bamboo. J. Fungi 2023, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, Y.H.; Ye, C.Y.; Lin, X.Y.; Su, N.; Fei, B.H. Improving the dimensional stability of round bamboo by environment-friendly modified rosin. Constr. Build. Mater. 2023, 365, 130078. [Google Scholar] [CrossRef]
- Salvati, E.; Brandt, L.R.; Uzun, F.; Zhang, H.; Papadaki, C.; Korsunsky, A.M. Multiscale analysis of bamboo deformation mechanisms following NaOH treatment using X-ray and correlative microscopy. Acta Biomater. 2018, 72, 329–341. [Google Scholar] [CrossRef]
- Chen, H.; Wu, J.Y.; Shi, J.J.; Wang, G.; Zhang, W.F. Dimensional Stability and Mold Resistance of Bamboo Slivers Treated by Alkali. Bioresources 2022, 17, 2827–2848. [Google Scholar] [CrossRef]
- Yihui, D.; Chan, J.C. The East Asian summer monsoon: An overview. Meteorol. Atmos. Phys. 2005, 89, 117–142. [Google Scholar] [CrossRef]
- Wang, X.; Dong, X.; Deng, Y.; Cui, C.; Wan, R.; Cui, W. Contrasting pre-Mei-Yu and Mei-Yu extreme precipitation in the Yangtze River valley: Influencing systems and precipitation mechanisms. J. Hydrometeorol. 2019, 20, 1961–1980. [Google Scholar] [CrossRef]
- GB/T 27651-2011; Use Category and Specification for Preservative-Treated Wood. Standardization Administration of China: Beijing, China, 2011.
- Omasa, T. Gene amplification and its application in cell and tissue engineering. J. Biosci. Bioeng. 2002, 94, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.J.; Czechowski, P.; Soubrier, J.; Stevens, M.I.; Cooper, A. Modular tagging of amplicons using a single PCR for high-throughput sequencing. Mol. Ecol. Resour. 2014, 14, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.; Ng, W.; Thao, K.; Agulto, R.; Weis, A.; Kim, K.S.; Korlach, J.; Hickey, L.; Kelly, L.; Lappin, S.; et al. Automation of PacBio SMRTbell NGS library preparation for bacterial genome sequencing. Stand. Genom. Sci. 2017, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Shokralla, S.; Porter, T.M.; Gibson, J.F.; Dobosz, R.; Janzen, D.H.; Hallwachs, W.; Golding, G.B.; Hajibabaei, M. Massively parallel multiplex DNA sequencing for specimen identification using an Illumina MiSeq platform. Sci. Rep. 2015, 5, 9687. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Rai, S.N.; Qian, C.; Pan, J.M.; Rai, J.P.; Song, M.; Bagaitkar, J.; Merchant, M.; Cave, M.; Egilmez, N.K.; McClain, C.J. Microbiome data analysis with applications to pre-clinical studies using QIIME2: Statistical considerations. Genes Dis. 2021, 8, 215–223. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D.J. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Ali, S.R.; Fradi, A.J.; Al-Aaraji, A.M.J. Effect of some physical factors on growth of five fungal species. Eur. Acad. Res. 2017, 2, 1069–1078. [Google Scholar]
- Liu, B.; Hu, Y.; Wang, Y.; Xue, H.; Li, Z.; Li, M. Effects of saline-alkali stress on bacterial and fungal community diversity in Leymus chinensis rhizosphere soil. Environ. Sci. Pollut. Res. 2022, 29, 70000–70013. [Google Scholar] [CrossRef]
- Kim, J.-J.; Lee, S.-S.; Ra, J.-B.; Lee, H.; Huh, N.; Kim, G.-H. Fungi associated with bamboo and their decay capabilities. Holzforschung 2011, 65, 271–275. [Google Scholar] [CrossRef]
- Osono, T. Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Vivelo, S.; Bhatnagar, J.M. An evolutionary signal to fungal succession during plant litter decay. FEMS Microbiol. Ecol. 2019, 95, fiz145. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.X.; Rudolf, V.H.W. Bridging theory and experiments of priority effects. Trends Ecol. Evol. 2023, 38, 1203–1216. [Google Scholar] [CrossRef] [PubMed]
- Sprockett, D.; Fukami, T.; Relman, D.A. Role of priority effects in the early-life assembly of the gut microbiota. Nat. Rev. Gastro Hepat. 2018, 15, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Debray, R.; Herbert, R.A.; Jaffe, A.L.; Crits-Christoph, A.; Power, M.E.; Koskella, B. Priority effects in microbiome assembly. Nat. Rev. Microbiol. 2022, 20, 109–121. [Google Scholar] [CrossRef]
- Liers, C.; Arnstadt, T.; Ullrich, R.; Hofrichter, M. Patterns of lignin degradation and oxidative enzyme secretion by different wood-and litter-colonizing basidiomycetes and ascomycetes grown on beech-wood. FEMS Microbiol. Ecol. 2011, 78, 91–102. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; De Sabata, D.; Fornasier, F. The enzyme patterns of Ascomycota and Basidiomycota fungi reveal their different functions in soil. Appl. Soil Ecol. 2024, 196, 105323. [Google Scholar] [CrossRef]
- COSTA, P.P.; Rosado, A.W.; Pereira, O.L. Six new species of Cladosporium associated with decayed leaves of native bamboo (Bambusoideae) in a fragment of Brazilian Atlantic Forest. Phytotaxa 2022, 560, 1–29. [Google Scholar] [CrossRef]
- Shen, X.Y.; Cheng, Y.L.; Cai, C.J.; Fan, L.; Gao, J.; Hou, C.L. Diversity and Antimicrobial Activity of Culturable Endophytic Fungi Isolated from Moso Bamboo Seeds. PLoS ONE 2014, 9, e95838. [Google Scholar] [CrossRef]
- Hyde, K.; Ho, W.; McKenzie, E.; Dalisay, T.J.F.D. Saprobic fungi on bamboo culms. Fungal Divers. 2001, 7, 35–48. [Google Scholar]
- Sampaio, J.P.; Golubev, W.I.; Fell, J.W.; Gadanho, M.; Golubev, N.W. Curvibasidium cygneicollum gen. nov., sp. nov. and Curvibasidium pallidicorallinum sp. nov., novel taxa in the Microbotryomycetidae (Urediniomycetes), and their relationship with Rhodotorula fujisanensis and Rhodotorula nothofagi. Int. J. Syst. Evol. Microbiol. 2004, 54, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Knapp, D.G.; Lázár, A.; Molnár, A.; Vajna, B.; Karácsony, Z.; Váczy, K.Z.; Kovács, G.M. Above-ground parts of white grapevine Vitis vinifera cv. Furmint share core members of the fungal microbiome. Environ. Microbiol. Rep. 2021, 13, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Mašínová, T.; Bahnmann, B.D.; Větrovský, T.; Tomšovský, M.; Merunková, K.; Baldrian, P. Drivers of yeast community composition in the litter and soil of a temperate forest. FEMS Microbiol. Ecol. 2017, 93, fiw223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of fungal communities in the marine sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, srep14524. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Schlatter, D.C.; Glawe, D.A.; Edwards, C.G.; Weller, D.M.; Paulitz, T.C.; Abatzoglou, J.T.; Okubara, P.A. Native yeast and non-yeast fungal communities of Cabernet Sauvignon berries from two Washington State vineyards, and persistence in spontaneous fermentation. Int. J. Food Microbiol. 2021, 350, 109225. [Google Scholar] [CrossRef] [PubMed]
- Oberwinkler, F.; Bandoni, R.; Blanz, P.; Kisimova-Horovitz, L. Cystofilobasidium: A new genus in the Filobasidiaceae. Syst. Appl. Microbiol. 1983, 4, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Gadanho, M.; van Broock, M.; Sampaio, J.P. Cystofilobasidium lacus-mascardii sp. nov., a basidiomycetous yeast species isolated from aquatic environments of the Patagonian Andes, and Cystofilobasidium macerans sp. nov., the sexual stage of Cryptococcus macerans. Int. J. Syst. Evol. Microbiol. 2009, 59, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Pontes, A.; Röhl, O.; Carvalho, C.; Maldonado, C.; Yurkov, A.M.; Paulo Sampaio, J. Cystofilobasidium intermedium sp. nov. and Cystofilobasidium alribaticum fa sp. nov., isolated from Mediterranean forest soils. Int. J. Syst. Evol. Microbiol. 2016, 66, 1058–1062. [Google Scholar] [CrossRef]
- Butinar, L.; Spencer-Martins, I.; Gunde-Cimerman, N. Yeasts in high Arctic glaciers: The discovery of a new habitat for eukaryotic microorganisms. Antonie van Leeuwenhoek 2007, 91, 277–289. [Google Scholar] [CrossRef]
- Jin, L.; Wu, D.; Li, C.; Zhang, A.; Xiong, Y.; Wei, R.; Zhang, G.; Yang, S.; Deng, W.; Li, T. Bamboo nutrients and microbiome affect gut microbiome of giant panda. Symbiosis 2020, 80, 293–304. [Google Scholar] [CrossRef]
- Jin, L.; Huang, Y.; Yang, S.; Wu, D.; Li, C.; Deng, W.; Zhao, K.; He, Y.; Li, B.; Zhang, G. Diet, habitat environment and lifestyle conversion affect the gut microbiomes of giant pandas. Sci. Total Environ. 2021, 770, 145316. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Yang, S.; Li, C.; Wu, D.; He, Y.; Li, B.; Zhang, G.; Xiong, Y.; Wei, R.; Li, G.J. Dietary changes and lifestyle shifts affect the gut microbiomes of giant pandas. Authorea Prepr. 2020. [Google Scholar] [CrossRef]
- Tokumasu, S. Fungal successions on pine needles fallen at different seasons: The succession of surface colonizers. Mycoscience 1998, 39, 417–423. [Google Scholar] [CrossRef]
- Bai, L.; Cheng, C.; Sun, M.-L.; Li, J.; Zou, Y.; Zhao, Q.; Zhao, X.-Q.J. Production of single cell oil by two novel nonconventional yeast strains of Curvibasidium sp. isolated from medicinal lichen. FEMS Yeast Res. 2023, 23, foad026. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-Y.; Wei, X.-Y.; Liu, X.-Z.; Bai, F.-Y.J. Cystofilobasidium josepaulonis sp. nov., a novel basidiomycetous yeast species. Int. J. Syst. Evol. Microbiol. 2023, 73, 005865. [Google Scholar] [CrossRef] [PubMed]
- Tervonen, K.; Oldén, A.; Taskinen, S.; Halme, P. The effects of grazing history, soil properties and stand structure on the communities of saprotrophic fungi in wood-pastures. Fungal Ecol. 2022, 60, 101163. [Google Scholar] [CrossRef]
- De Queiroz, M.E.F.; Monteiro, J.S.; Viana-Junior, A.B.; Praxedes, C.d.L.B.; Lavelle, P.; Vasconcelos, S.S. Litter thickness and soil pH influence the diversity of saprotrophic fungi in primary forest fragments in the Amazon. Pedobiologia 2021, 89, 150771. [Google Scholar] [CrossRef]
- Yamanaka, T. The effect of pH on the growth of saprotrophic and ectomycorrhizal ammonia fungi in vitro. Mycologia 2003, 95, 584–589. [Google Scholar] [CrossRef]
- Zhang, W.B.; Tian, G.L.; Polle, A.; Janz, D.; Euring, D.; Yue, X.H.; Zhao, H.S.; Fei, B.H.; Jiang, Z.H. Comparative characterization of ethanol organosolv lignin polymer from bamboo green, timber and yellow. Wood Sci. Technol. 2018, 52, 1331–1341. [Google Scholar] [CrossRef]
- Yu, H.X.; Pan, X.; Xu, M.P.; Yang, W.M.; Wang, J.; Zhuang, X.W. Surface chemical changes analysis of UV-light irradiated Moso bamboo (Phyllostachys pubescens Mazel). R. Soc. Open Sci. 2018, 5, 180110. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Kong, C.-C.; Wang, Z.-Z.; Wang, Z.; Liu, G.-L.; Hu, Z.; Chi, Z.-M. The signaling pathways involved in metabolic regulation and stress responses of the yeast-like fungi Aureobasidium spp. Biotechnol. Adv. 2022, 55, 107898. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.-N.; Chi, Z.; Liu, G.-L.; Chen, T.-J.; Jiang, H.; Hu, Z.; Chi, Z.-M. α-Amylase, glucoamylase and isopullulanase determine molecular weight of pullulan produced by Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2018, 117, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Zhang, L.; Chi, Z.; Wang, X. A carboxymethyl cellulase from a marine yeast (Aureobasidium pullulans 98): Its purification, characterization, gene cloning and carboxymethyl cellulose digestion. J. Ocean Univ. China 2015, 14, 913–921. [Google Scholar] [CrossRef]
- Rich, J.O.; Leathers, T.D.; Anderson, A.M.; Bischoff, K.M.; Manitchotpisit, P. Laccases from Aureobasidium pullulans. Enzym. Microb. Technol. 2013, 53, 33–37. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, W.; Wang, J.; Shen, G.; Yuan, Y.; Yan, L.; Tang, H.; Wang, W.J.A.; Microbiology, E. Isolation and characterization of a novel laccase for lignin degradation, LacZ1. Appl. Environ. Microbiol. 2021, 87, e01355-21. [Google Scholar] [CrossRef]
- Hoegger, P.J.; Kilaru, S.; James, T.Y.; Thacker, J.R.; Kües, U. Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J. 2006, 273, 2308–2326. [Google Scholar] [CrossRef]
- Wang, X.; He, J.; Pang, S.; Yao, S.; Zhu, C.; Zhao, J.; Liu, Y.; Liang, C.; Qin, C. High-efficiency and high-quality extraction of hemicellulose of bamboo by freeze-thaw assisted two-step alkali treatment. Int. J. Mol. Sci. 2022, 23, 8612. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Lv, Z.; Rao, J.; Sun, D.; Hu, Y.; Yue, P.; Tian, R.; Lü, B.; Bian, J.; Peng, F. The effect of bamboo (Phyllostachys pubescens) cell types on the structure of hemicelluloses. Ind. Crop. Prod. 2022, 187, 115464. [Google Scholar] [CrossRef]
- Brito, F.M.S.; Paes, J.B.; Oliveira, J.T.d.S.; Arantes, M.D.C.; Fantuzzi Neto, H. Caracterização anatômica e física do bambu gigante (Dendrocalamus giganteus Munro). Floresta Ambiente 2015, 22, 559–566. [Google Scholar] [CrossRef]
- Abdella, A.; Segato, F.; Wilkins, M.R. Optimization of process parameters and fermentation strategy for xylanase production in a stirred tank reactor using a mutant Aspergillus nidulans strain. Biotechnol. Rep. 2020, 26, e00457. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, S.; Shang, W.; Yan, Z.; Wu, X.; Li, Y.; Chen, G.; Liu, X.; Wang, L. Synergistic mechanism of GH11 xylanases with different action modes from Aspergillus niger An76. Biotechnol. Biofuels 2021, 14, 118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Bao, C.; Dong, B.; Cao, Y. Characterization of a novel GH10 xylanase with a carbohydrate binding module from Aspergillus sulphureus and its synergistic hydrolysis activity with cellulase. Int. J. Biol. Macromol. 2021, 182, 701–711. [Google Scholar] [CrossRef]
- Brandt, S.C.; Ellinger, B.; Van Nguyen, T.; Harder, S.; Schlüter, H.; Hahnke, R.L.; Rühl, M.; Schäfer, W.; Gand, M. Aspergillus sydowii: Genome analysis and characterization of two heterologous expressed, non-redundant xylanases. Front. Microbiol. 2020, 11, 2154. [Google Scholar] [CrossRef]
- Intasit, R.; Cheirsilp, B.; Suyotha, W.; Boonsawang, P. Purification and characterization of a highly-stable fungal xylanase from Aspergillus tubingensis cultivated on palm wastes through combined solid-state and submerged fermentation. Prep. Biochem. Biotechnol. 2022, 52, 311–317. [Google Scholar] [CrossRef]
- Chreptowicz, K.; Mierzejewska, J.; Tkáčová, J.; Młynek, M.; Čertik, M. Carotenoid-producing yeasts: Identification and characteristics of environmental isolates with a valuable extracellular enzymatic activity. Microorganisms 2019, 7, 653. [Google Scholar] [CrossRef] [PubMed]
- Ra, J.-B.; Oh, J.-J.; Kim, G.-H. Optimization of alkali pretreatment conditions for wax removal from bamboo culm. BioResources 2020, 15, 8202. [Google Scholar] [CrossRef]
- Chang, S.-T.; Wu, J.-H.; Yeh, T.-F. Effects of chromated-phosphate treatment process on the green color protection of ma bamboo (Dendrocalamus latiflorus). J. Wood Sci. 2002, 48, 227–231. [Google Scholar] [CrossRef]
- Chang, S.-T.; Yeh, T.-F. Protection and fastness of green color of moso bamboo (Phyllostachys pubescens Mazel) treated with chromium-based reagents. J. Wood Sci. 2001, 47, 228–232. [Google Scholar] [CrossRef]
- Petridis, L.; Smith, J.C. Molecular-level driving forces in lignocellulosic biomass deconstruction for bioenergy. Nat. Rev. Chem. 2018, 2, 382–389. [Google Scholar] [CrossRef]
- Ma, F.Y.; Huang, X.; Ke, M.; Shi, Q.P.; Chen, Q.; Shi, C.C.; Zhang, J.; Zhang, X.Y.; Yu, H.B. Role of Selective Fungal Delignification in Overcoming the Saccharification Recalcitrance of Bamboo Culms. ACS Sustain. Chem. Eng. 2017, 5, 8884–8894. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; An, X.; He, X.; Ren, X.; Sichone, J.; Wu, X.; Zhang, Y.; Wang, H.; Sun, F. Temporal Dynamics of Fungal Communities in Alkali-Treated Round Bamboo Deterioration under Natural Weathering. Microorganisms 2024, 12, 858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050858
Han S, An X, He X, Ren X, Sichone J, Wu X, Zhang Y, Wang H, Sun F. Temporal Dynamics of Fungal Communities in Alkali-Treated Round Bamboo Deterioration under Natural Weathering. Microorganisms. 2024; 12(5):858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050858
Chicago/Turabian StyleHan, Shuaibo, Xiaojiao An, Xiaolong He, Xin Ren, John Sichone, Xinxing Wu, Yan Zhang, Hui Wang, and Fangli Sun. 2024. "Temporal Dynamics of Fungal Communities in Alkali-Treated Round Bamboo Deterioration under Natural Weathering" Microorganisms 12, no. 5: 858. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050858