
The Impact of Artificial Restoration of Alpine Grasslands in the Qilian Mountains on Vegetation, Soil Bacteria, and Soil Fungal Community Diversity

Abstract
:1. Introduction
2. Materials and Methods
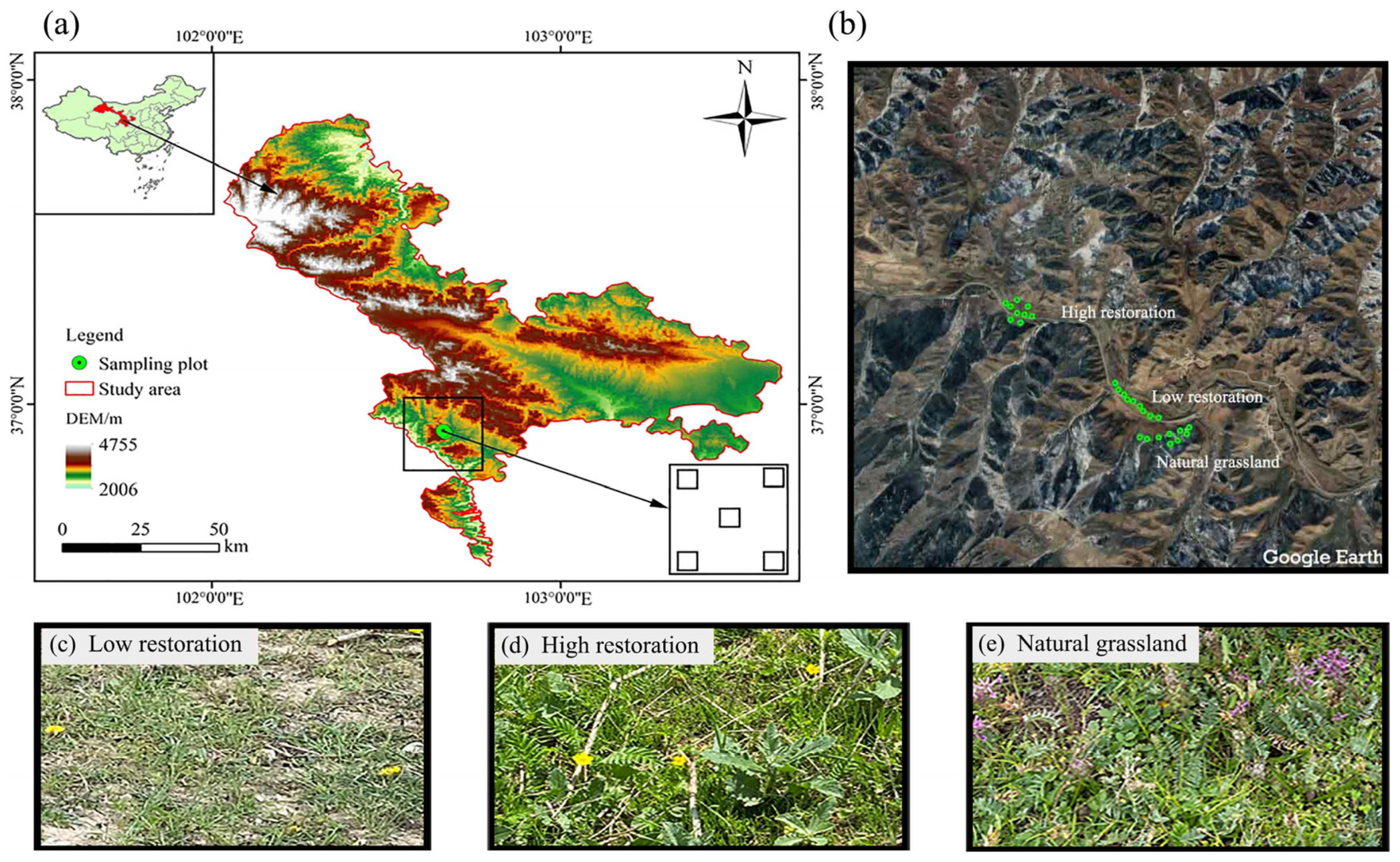
2.1. Study Region
2.2. Plotting and Sample Collection
2.3. Sample Analysis
2.4. Bioinformatics Analysis
2.5. Data Analysis
3. Results
3.1. Traits of Vegetation Communities within Alpine Grasslands of Mining Regions across Varying Restoration Levels
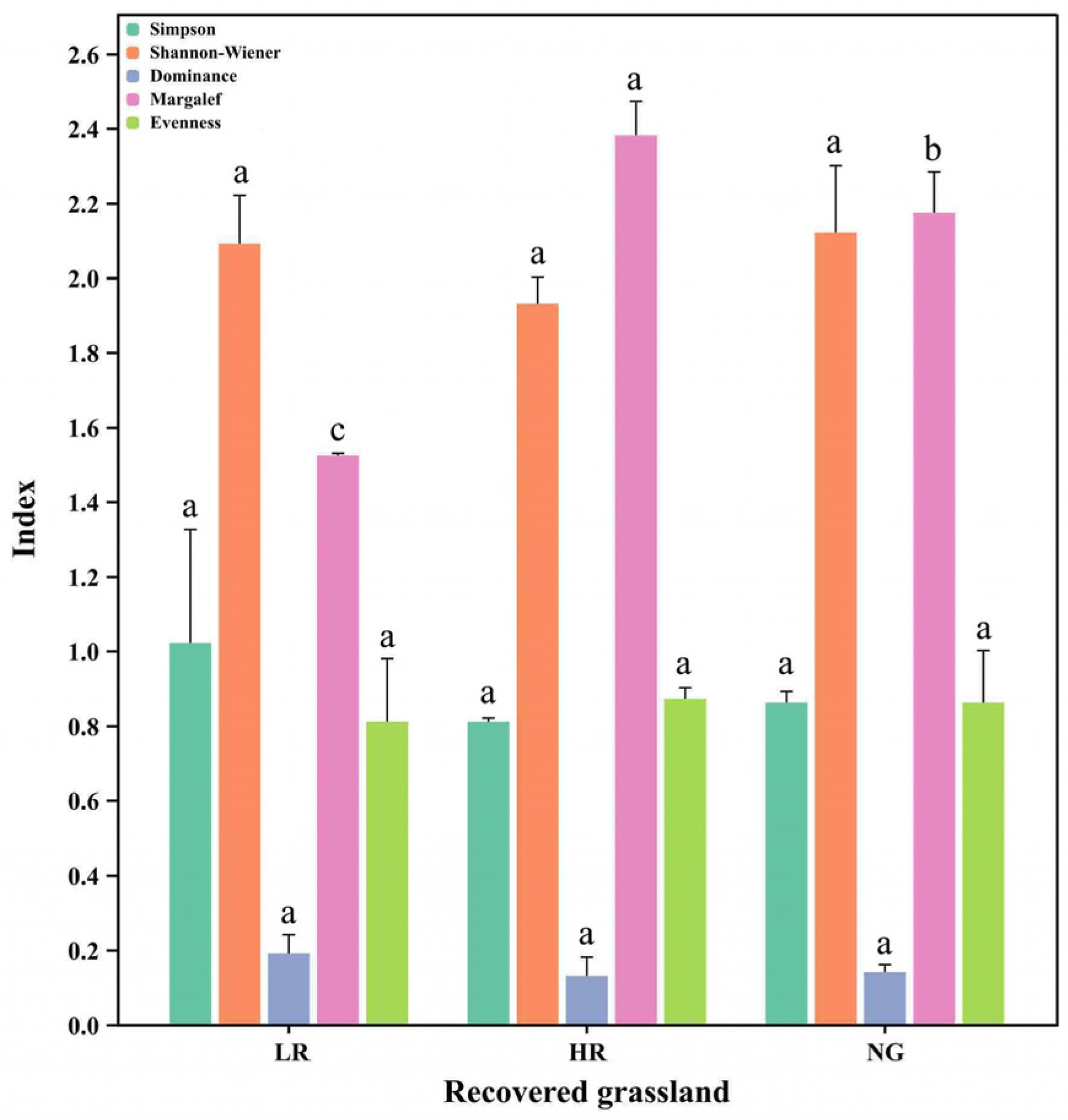
3.2. Effects of Vegetation Restoration in Alpine Mining Areas on Grassland Species Diversity
3.3. Effects of Vegetation Restoration on Soil Properties in Alpine Mining Areas
3.4. Impact of Vegetation Restoration on Soil Bacterial Communities in Alpine Mining Areas
3.4.1. Structure of Soil Bacterial Populations in Grassland Ecosystems within Mining Zones at Varying Stages of Ecological Restoration
3.4.2. Relative Abundance of Soil Bacterial Community Composition in Grasslands of Mining Areas with Different Degrees of Restoration
3.4.3. Examination of the Hierarchical Structure of Soil Bacterial Populations in Grassland Regions within Mining Zones across Varying Levels of Ecological Restoration
3.4.4. Variability in the Soil Bacterial Populations within Grasslands of Mining Areas with Different Degrees of Restoration
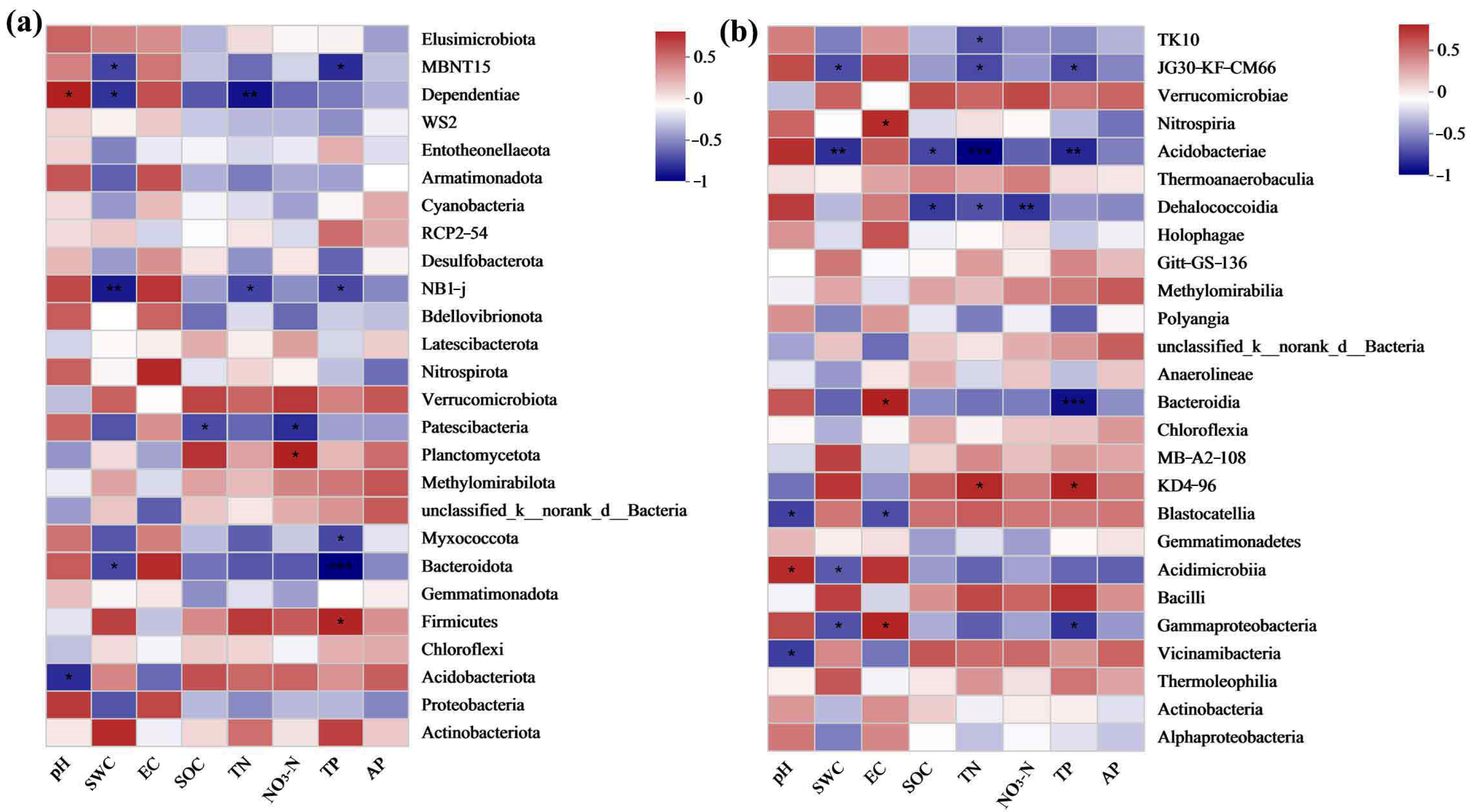
3.4.5. Association between Soil Bacterial Communities and Soil Properties in Grasslands of Mining Areas with Different Degrees of Restoration
3.4.6. Analysis of Soil Bacterial Covariance Networks in Grasslands of Mining Sites with Different Levels of Restoration
3.5. Impact of Vegetation Recovery on Soil Fungal Populations in High-Altitude Mining Regions
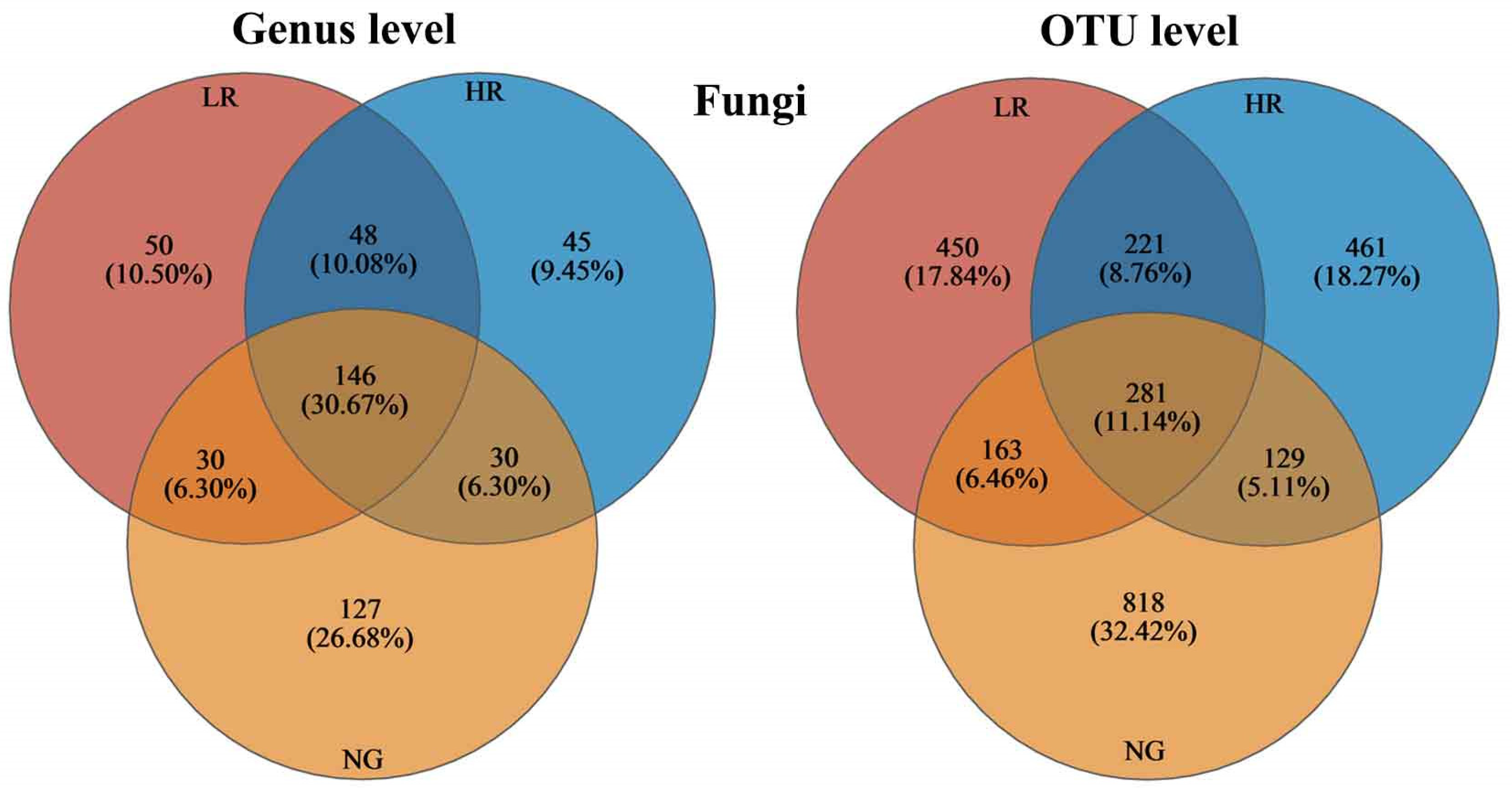
3.5.1. Variation in Soil Fungal Community Composition across Grasslands in Mining Areas with Varying Levels of Restoration
3.5.2. Relative Abundance of Soil Fungal Community Composition in Grasslands of Mining Areas with Different Degrees of Restoration
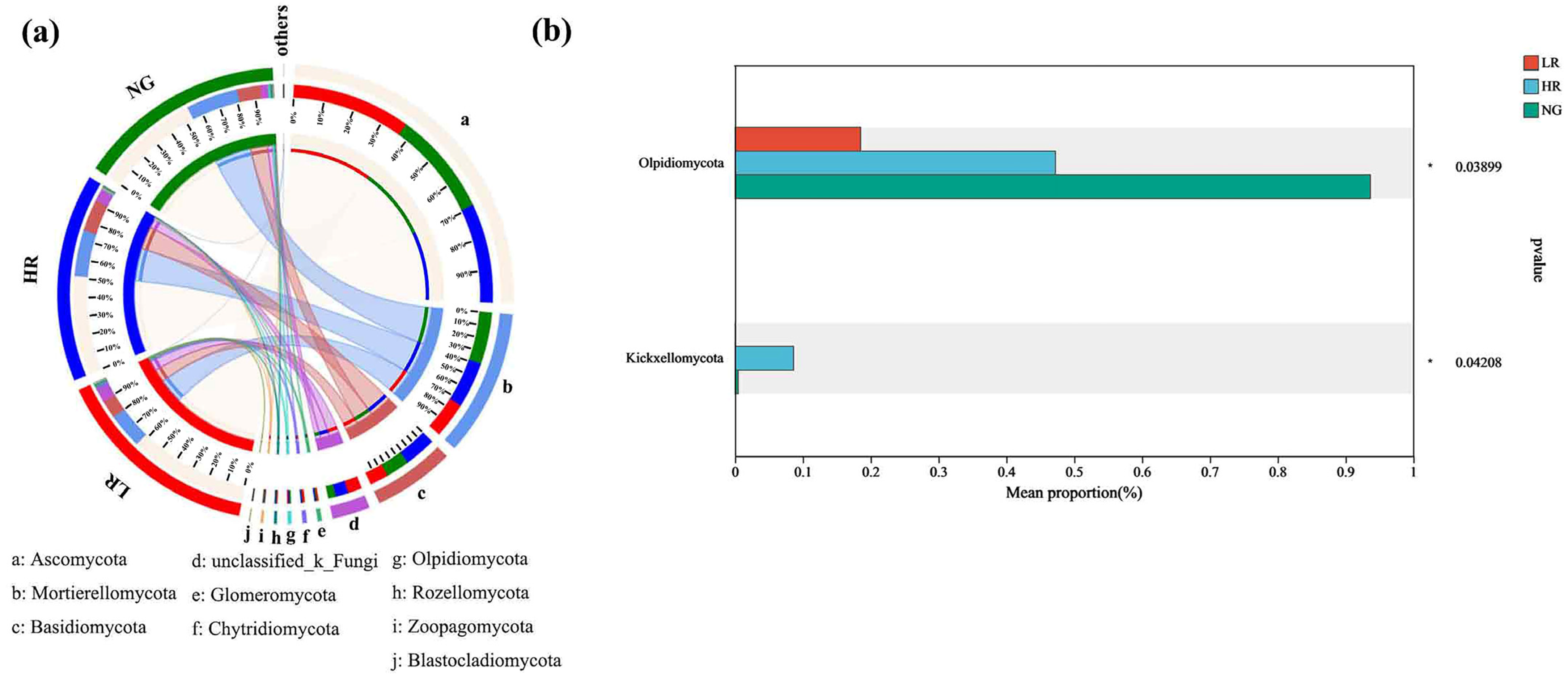
3.5.3. Analysis of the Gate-Level Composition of Soil Fungal Communities within Grasslands in Mining Regions Exhibiting Varying Levels of Restoration
3.5.4. Diversity of Soil Fungal Communities in Grasslands of Mining Regions Exhibiting Varying Levels of Restoration
3.5.5. Correlation between Soil Fungal Communities and Soil Properties in Grasslands of Mining Regions Exhibiting Varying Levels of Restoration
3.5.6. Analysis of Soil Fungal Covariance Networks in Grasslands of Mining Sites with Different Levels of Restoration
4. Discussion
4.1. Impacts of Varying Degrees of Vegetation Restoration on Plant Diversity
4.2. Impacts of Different Restoration Levels of Vegetation on Soil Properties
4.3. Analysis of Variations in the Diversity of Soil Microbial Communities
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Choudoir, M.J.; Deangelis, K.M. A framework for integrating microbial dispersal modes into soil ecosystem ecology. iScience. 2022, 25, 103887. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.W.; Liu, Z.Y.; Wu, Z.H.; Huang, J.; Zhang, L.; Zhang, G.G.; Jia, M.Q.; Han, G.D. Soil nematode communities and functions: Ecological package restoration responses in damaged desert steppe open-pit mining ecosystems. Land. Degrad. Dev. 2022, 11, 1015–1028. [Google Scholar] [CrossRef]
- Rania, R.; Joana, D.; Omar, D. A tunnel under an in-pit mine waste dump to improve environmental and landscape recovery of the site. Minerals 2021, 11, 2–11. [Google Scholar]
- Lei, T.; Feng, J.; Lv, J.; Wang, J.; Song, H.; Gao, X. Net primary productivity loss under different drought levels in different grassland ecosystems. J. Environ. Manag. 2020, 274, 111144. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ali, A. Climate regulates the functional traits aboveground biomass relationships at a community-level in forests: A global meta-analysis. Sci. Total Environ. 2021, 761, 143238. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Wang, Y.H.; Webb, A.A. Influence of climatic and geographic factors on the spatial distribution of Qinghai spruce forests in the dryland Qilian Mountains of northwest China. Sci. Total Environ. 2018, 612, 1007–1017. [Google Scholar] [CrossRef]
- Kou, Z.Y.; Li, C.Y.; Chang, S. Effects of nitrogen and phosphorus additions on soil microbial community structure and ecological processes in the farmland of Chinese Loess Plateau. J. Arid. Land. 2023, 15, 960–974. [Google Scholar] [CrossRef]
- Prach, K.; Šebelíková, L.; Řehounková, K. Possibilities and limitations of passive restoration of heavily disturbed sites. Landsc. Res. 2020, 45, 247–253. [Google Scholar] [CrossRef]
- Lorite, J.; Ballesteros, M.; García-Robles, H. Economic evaluation of ecological restoration options in gypsum habitats after mining. J. Nat. Conserv. 2021, 59, 125935. [Google Scholar] [CrossRef]
- Han, X.; Li, Y.; Du, X.; Li, Y.; Wang, Z.; Jiang, S.; Li, Q. Effect of grassland degradation on soil quality and soil biotic community in a semi-arid temperate steppe. Ecol. Process. 2020, 9, 63. [Google Scholar] [CrossRef]
- Li, P.; Zhang, X.; Hao, M.; Cui, Y.; Zhu, S.; Zhang, Y. Effects of vegetation restoration on soil bacterial communities, enzyme activities, and nutrients of reconstructed soil in a mining area on the Loess Plateau, China. Sustainability 2019, 11, 2295. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, F.; Wang, X.; Lu, N. Effects of meteorology and soil on the herb species diversity in plantations in a reclamation area of coal mine after 6 years. Environ. Sci. Pollut. Res. 2020, 27, 24231–24241. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Bai, X.; Zhou, Y. Variations of soil microbial communities accompanied by different vegetation restoration in an open-cut iron mining area. Sci. Total Environ. 2020, 704, 135243. [Google Scholar] [CrossRef] [PubMed]
- Ilunga, E.I.W.; Mahy, G.; Piqueray, J. Plant functional traits as a promising tool for the ecological restoration of degraded tropical metal-rich habitats and revegetation of metal-rich bare soils: A case study in copper vegetation of Katanga, DRC. Ecol. Eng. 2015, 82, 214–221. [Google Scholar] [CrossRef]
- Islam, M.S.; Haque, K.A.; Jahan, N.; Atikullah, M.; Uddin, M.N.; Naser, A.M.; Faruk-E-Azam, A.K.M. Soil salinity mitigation by naturally grown halophytes in seawater affected coastal Bangladesh. Int. J. Environ. Sci. Technol. 2022, 35, 11013–11022. [Google Scholar] [CrossRef]
- Aili, A.; Xu, H.; Zhao, X.; Zhang, P.; Yang, R. Dynamics of Vegetation Productivity in Relation to Surface Meteorological Factors in the Altay Mountains in Northwest China. Forests 2022, 13, 1907. [Google Scholar] [CrossRef]
- Kong, J.Q.; He, Z.B.; Chen, L.F.; Yang, R.; Du, J. Efficiency of biochar, nitrogen addition, and microbial agent amendments in remediation of soil properties and microbial community in Qilian Mountains mine soils. Ecol. Evol. 2021, 11, 9318–9331. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Feng, Q.; Liu, W.; Xia, H.H.; Zhang, J.T.; Yang, L.S.; Zhang, C.Q.; Wang, Z.Y.; Feng, Y.L. Community structure and plant diversity under different degrees of restored grassland in mining areas of the Qilian Mountains, Northwestern China. Front. Environ. Sci. 2023, 11, 1191599. [Google Scholar] [CrossRef]
- Gao, Y.; An, Y.; Qi, B.; Liu, J.; Yu, H.; Wang, D. Grazing exclusion mediates the trade-off between plant diversity and productivity in Leymus chinensis meadows along a chronosequence on the Songnen Plain. China Ecol. Indic. 2021, 126, 107655. [Google Scholar] [CrossRef]
- Pansu, M.; Gautheyrou, J. pH Measurement; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 3540312102. [Google Scholar]
- Pei, L.; Xiao, J.; Sun, L. The effects of reclaimed water irrigation on the soil characteristics and microbial populations of plant rhizosphere. Environ. Sci. Pollut. Res. 2021, 29, 17570–17579. [Google Scholar]
- Yang, X.M.; Feng, Q.; Zhu, M.; Yang, L.S.; Zhang, C.Q.; Zhang, J.T.; Wang, Z.Y.; Feng, Y.L. Changes in nutrient-regulated soil microbial communities in soils concomitant with grassland restoration in the alpine mining region of the Qilian Mountains. Agronomy 2023, 13, 3052. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Wu, L.Y.; Deng, Y.; Zhi, X.Y.; Jiang, Y.H.; Tu, Q.C.; Xie, J.P.; Van Nostrand, J.D.; He, Z.L.; Yang, Y.F. Reproducibility and quantitation of amplicon sequencing-based detection. ISME J. 2011, 5, 1303–1313. [Google Scholar] [CrossRef]
- Leung, M.H.Y.; Chan, K.C.K.; Lee, P.K.H. Skin fungal community and its correlation with bacterial community of urban Chinese individuals. Microbiome 2016, 4, 46. [Google Scholar] [CrossRef]
- Liu, Y.; Lei, S.G.; Chen, X.Y.; Chen, M.; Zhang, X.Y.; Long, L.L. Study of plant configuration pattern in guided vegetation restoration: A case study of semiarid underground mining areas in Western China. Ecol. Eng. 2021, 170, 106334. [Google Scholar] [CrossRef]
- Tan, J.J.; Liang, J.; Yang, Y.J.; Zhang, S.L.; Hou, H.P.; Zhu, X.X. Revealing the Structure and Composition of the Restored Vegetation Cover in Semi-Arid Mine Dumps Based on LiDAR and Hyperspectral Images. Remote Sens. 2022, 14, 978. [Google Scholar]
- Xiao, W.; Lv, X.J.; Zhao, Y.L.; Sun, H.X.; Li, J.Q. Ecological resilience assessment of an arid coal mining area using index of entropy and linear weighted analysis: A case study of Shendong coalfield. China. Ecol. Indic. 2020, 109, 105843. [Google Scholar] [CrossRef]
- Yang, Y.J.; Erskine, P.D.; Zhang, S.L.; Wang, Y.J.; Bian, Z.F.; Lei, S.G. Effects of underground mining on vegetation and environmental patterns in a semi-arid watershed with implications for resilience management. Environ. Earth Sci. 2018, 77, 1–12. [Google Scholar] [CrossRef]
- Wei, Z.F.; Huang, Q.Y.; Zhang, R. Dynamics of vegetation coverage and response to climate change in China-South Asia-Southeast Asia during 1982–2013. Appl. Ecol. Environ. Res. 2019, 17, 2865–2879. [Google Scholar] [CrossRef]
- Zhang, Q.D.; Wei, W.; Chen, L.D.; Yang, L.; Luo, Y.Q.; Cai, A.D. Plant traits in influencing soil moisture in semiarid grasslands of the Loess Plateau, China. Sci. Total Environ. 2020, 718, 137355. [Google Scholar] [CrossRef]
- Chen, L.; He, Z.; Zhao, W.; Liu, J.; Zhou, H.; Li, J.; Meng, Y.; Wang, L. Soil structure and nutrient supply drive changes in soil microbial communities during conversion of virgin desert soil to irrigated cropland. Eur. J. Soil Sci. 2020, 71, 768–781. [Google Scholar] [CrossRef]
- Dai, Z.; Wang, Y.; Muhammad, N.; Yu, X.; Xiao, K.; Meng, J.; Liu, X.; Xu, J.; Brookes, P.C. The effects and mechanisms of soil acidity changes, following incorporation of biochars in three soils differing in initial pH. Soil. Sci. Soc. Am. J. 2014, 78, 1606–1614. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Lavallee, J.M. Soil organic matter formation, persistence, and functioning: A synthesis of current understanding to inform its conservation and regeneration. Adv. Agron. 2022, 172, 1–66. [Google Scholar]
- Schweigert, M.; Herrmann, S.; Miltner, A.; Fester, T.; Kastner, M. Fate of ectomycorrhizal fungal biomass in a soil bioreactor system and its contribution to soil organic matter formation. Soil. Biol. Biochem. 2015, 88, 120–127. [Google Scholar] [CrossRef]
- Billet, L.; Pesce, S.; Martin-Laurent, F.; Devers-Lamrani, F. Experimental evidence for manure-borne bacteria invasion in soil during a coalescent event: Influence of the antibiotic sulfamethazine. Microb. Ecol. 2023, 85, 1463–1472. [Google Scholar] [CrossRef]
- Fang, B.Z.; Salam, N.; Han, M.X.; Jiao, J.Y.; Cheng, J.; Wei, D.Q.; Xiao, M.; Li, W.J. Insights on the effects of heat pretreatment, pH, and calcium salts on isolation of rare Actinobacteria from karstic caves. Front. Microbiol. 2017, 8, 1535. [Google Scholar] [CrossRef]
- Li, G.; Zhang, Z.; Shi, L.; Zhou, Y.; Yang, M.; Cao, J.; Wu, S.; Lei, G. Effects of different grazing intensities on soil C, N, and P in an alpine meadow on the Qinghai—Tibetan plateau, China. Int. J. Environ. Res. Public Health 2018, 15, 2584. [Google Scholar] [CrossRef]
- Racheal, N.U.; Elizabeth, M.B.; Kirsten, S.H. Spatio-temporal microbial community dynamics within soil aggregates. Soil. Biol. Biochem. 2019, 132, 58–68. [Google Scholar]
- Hu, T.B.; Chang, J.; Liu, X.X.; Feng, S.S. Integrated methods for determining restoration priorities of coal mining subsidence areas based on green infrastructure:—A case study in the Xuzhou urban area, of China. Ecol. Indic. 2018, 94, 164–174. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Jiang, L.; Zhang, L.; Cui, S.; Meng, F.; Wang, Q.; Li, X.; Zhou, Y. Changes of soil microbial community under different degraded gradients of alpine meadow. Agric. Ecosyst. Environ. 2016, 222, 213–222. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, W.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass, diversity, and compositions to altitudinal gradients depend on plant and soil characteristics. Sci. Total Environ. 2018, 610–611, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Hogues, H.; Whyte, L.G. The functional potential of high Arctic permafrost revealed by metagenomic sequencing, qPCR and microarray analyses. ISME J. 2010, 4, 1206–1214. [Google Scholar] [CrossRef]
- Su, K.; Yu, Q.; Yue, D.; Zhang, Q.; Yang, L.; Liu, Z.; Niu, T.; Sun, X. Simulation of a forest-grass ecological network in a typical desert oasis based on multiple scenes. Ecol. Model. 2019, 413, 108834. [Google Scholar] [CrossRef]
- Wang, J.; Liu, G.B.; Zhang, C.; Wang, G.L.; Fang, L.C.; Cui, Y.X. Higher temporal turnover of soil fungi than bacteria during long-term secondary succession in a semiarid abandoned farmland. Soil. Tillage Res. 2019, 194, 104305. [Google Scholar] [CrossRef]
- Comer, J.; Perkins, L. Resistance of the soil microbial community to land-surface disturbances of high-intensity winter grazing and wildfire. J. Environ. Manag. 2021, 279, 111596. [Google Scholar] [CrossRef]
- Joergensen, R.G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biol. Fert. Soils. 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, N.; Wang, H.; Li, J.; Zhong, H.; Dong, H.; Zeng, Z.; Zong, C. Variations in the diversity of the soil microbial community and structure under various categories of degraded wetland in Sanjiang Plain, northeastern China. Land. Degrad. Dev. 2021, 32, 2143–2156. [Google Scholar] [CrossRef]
- Rabbi, S.M.F.; Minasny, B.; McBratney, A.B.; Young, I.M. Microbial processing of organic matter drives stability and pore geometry of soil aggregates. Geoderma 2020, 360, 114033. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, J.J.; Banerjee, S.; White, J.F.; Zhou, N.; Zhao, Z.Y. Not by salinity alone: How environmental factors shape fungal communities in saline soils. Soil. Sci. Soc. Am. J. 2019, 83, 1387–1398. [Google Scholar] [CrossRef]
- Zhang, H.; Fu, G. Responses of plant, soil bacterial and fungal communities to grazing vary with pasture seasons and grassland types, northern Tibet. Land. Degrad. Dev. 2021, 32, 1821–1832. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recovered Degree | Geography Coordinate | Altitude/m | Above-Ground Biomass (g m−2) | Vegetation Cover/% |
|---|---|---|---|---|
| Low restoration (LR) | 36°91′95″ N 102°66′83″ E | 2802 | 128.93 ± 1.19 b | 0.36 ± 0.02 b |
| High restoration (HR) | 36°91′94″ N 102°66′86″ E | 2811 | 420.83 ± 3.29 a | 0.72 ± 0.05 a |
| Natural grassland (NG) | 36°91′69″ N 102°66′42″ E | 2858 | 457.47 ± 1.75 a | 0.68 ± 0.03 a |
| LR | HR | NG | |||
|---|---|---|---|---|---|
| Species | IV | Species | IV | Species | IV |
| Cyperus rotundus | 24.53 | Cyperus rotundus | 29.43 | Cyperus rotundus | 21.18 |
| Carex spp. | 16.71 | Elymus nutans | 25.86 | Elymus nutans | 15.50 |
| Elymus nutans | 11.54 | Carex spp. | 10.67 | Carex spp. | 15.02 |
| Daucus carota | 10.46 | Artemisia smithii Mattf | 8.89 | Potentilla chinensis | 13.78 |
| Potentilla chinensis | 8.88 | Chenopodium glaucum | 7.46 | Oxytropis ochrocephala | 12.93 |
| Sonchus oleraceus | 4.69 | Sonchus oleraceus | 6.14 | Chenopodium glaucum | 8.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Feng, Q.; Zhu, M.; Zhang, J.; Yang, L.; Li, R. The Impact of Artificial Restoration of Alpine Grasslands in the Qilian Mountains on Vegetation, Soil Bacteria, and Soil Fungal Community Diversity. Microorganisms 2024, 12, 854. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050854
Yang X, Feng Q, Zhu M, Zhang J, Yang L, Li R. The Impact of Artificial Restoration of Alpine Grasslands in the Qilian Mountains on Vegetation, Soil Bacteria, and Soil Fungal Community Diversity. Microorganisms. 2024; 12(5):854. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050854
Chicago/Turabian StyleYang, Xiaomei, Qi Feng, Meng Zhu, Jutao Zhang, Linshan Yang, and Ruolin Li. 2024. "The Impact of Artificial Restoration of Alpine Grasslands in the Qilian Mountains on Vegetation, Soil Bacteria, and Soil Fungal Community Diversity" Microorganisms 12, no. 5: 854. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050854