
Chemical Characterization and Effect of a Lactobacilli-Postbiotic on Streptococcus mutans Biofilm In Vitro

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining the Postbiotic (Conditioned Culture Medium)
2.2. Gas Chromatography Coupled with Mass Spectrometry (GC–MS)
2.2.1. Analysis Method
2.2.2. Silylation Method
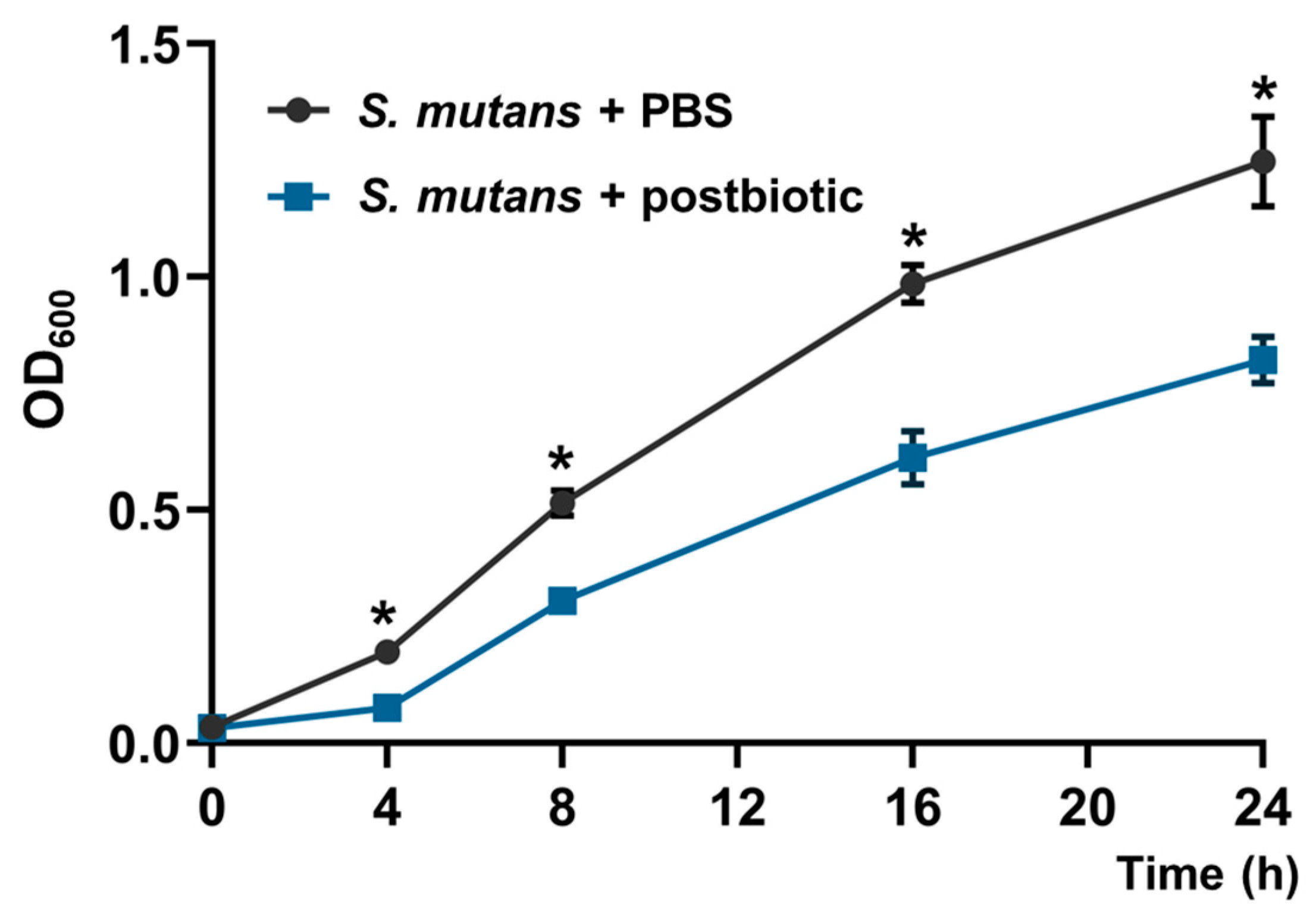
2.3. Effect on Planktonic Cells of S. mutans
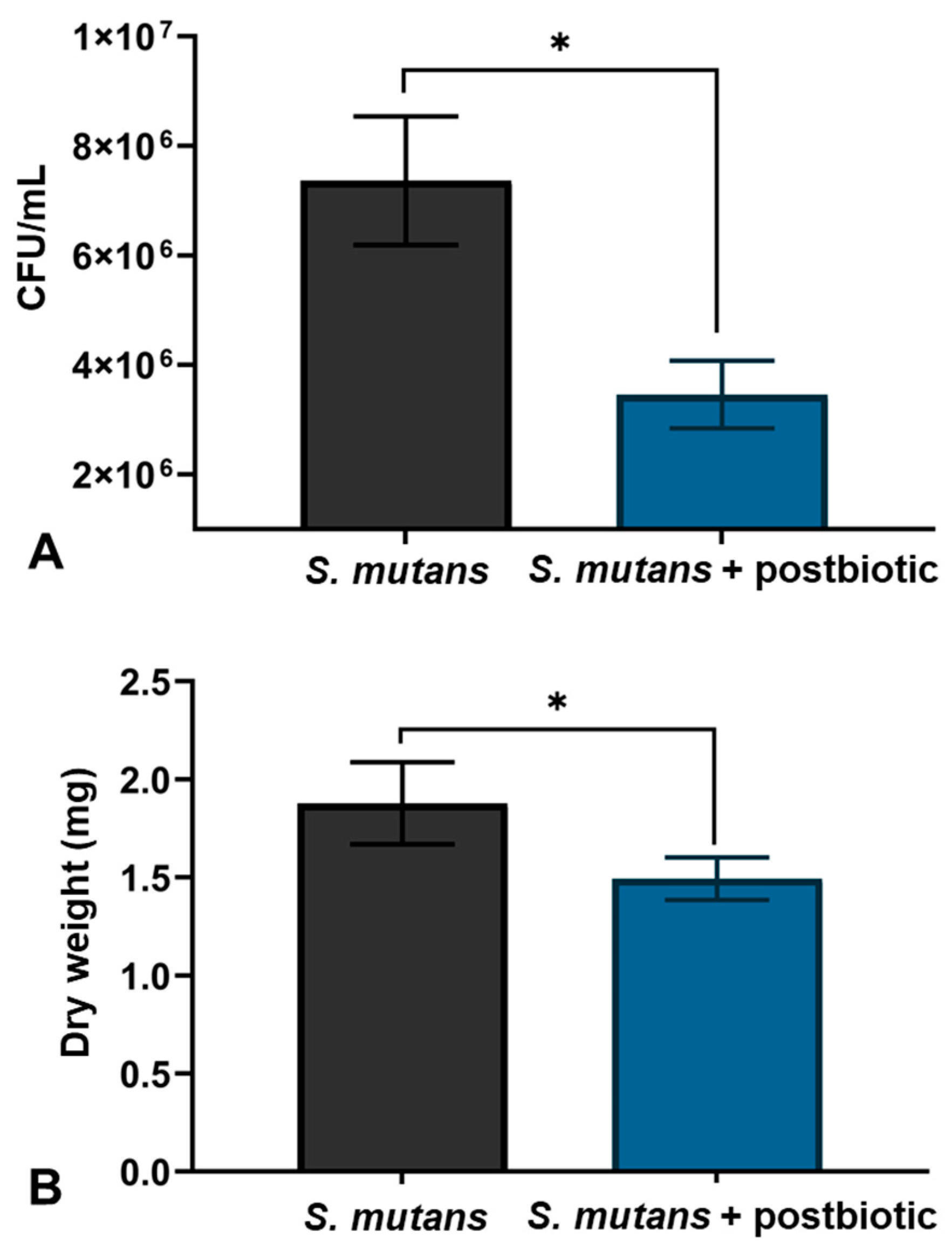
2.4. Effect on Biofilm of S. mutans
2.4.1. Determination of Live Bacteria Count
2.4.2. Determination of Dry Weight of Biofilms
2.4.3. Effect of Postbiotics on Gene Expression of S. mutans
2.5. Statistical Analysis
3. Results
3.1. Chemical Characterization
3.2. Effect of the Postbiotics on S. mutans
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Cicenia, A.; Scirocco, A.; Carabotti, M.; Pallotta, L.; Marignani, M.; Severi, C. Postbiotic activities of lactobacilli-derived factors. J. Clin. Gastroenterol. 2014, 48 (Suppl. S1), S18–S22. [Google Scholar] [CrossRef] [PubMed]
- Fayol-Messaoudi, D.; Berger, C.N.; Coconnier-Polter, M.H.; Liévin-Le Moal, V.; Servin, A.L. pH-, Lactic acid-, and non-lactic acid-dependent activities of probiotic Lactobacilli against Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2005, 71, 6008–6013. [Google Scholar] [CrossRef] [PubMed]
- Memorandum, D.T. Lactobacillus rhamnosus, 2008; Volume 32.
- Shen, S.; Ren, F.; Qin, H.; Bukhari, I.; Yang, J.; Gao, D.; Ouwehand, A.C.; Lehtinen, M.J.; Zheng, P.; Mi, Y. Lactobacillus acidophilus NCFM and Lactiplantibacillus plantarum Lp-115 inhibit Helicobacter pylori colonization and gastric inflammation in a murine model. Front. Cell Infect. Microbiol. 2023, 13, 1196084. [Google Scholar] [CrossRef] [PubMed]
- Alanzi, A.; Honkala, S.; Honkala, E.; Varghese, A.; Tolvanen, M.; Söderling, E. Effect of Lactobacillus rhamnosus and Bifidobacterium lactis on gingival health, dental plaque, and periodontopathogens in adolescents: A randomised placebocontrolled clinical trial. Benef. Microbes 2018, 9, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Cagetti, M.G.; Mastroberardino, S.; Milia, E.; Cocco, F.; Lingström, P.; Campus, G. The use of probiotic strains in caries prevention: A systematic review. Nutrients 2013, 5, 2530–2550. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.K.; Hasslöf, P.; Stecksén-Blicks, C.; Twetman, S. Co-aggregation and growth inhibition of probiotic lactobacilli and clinical isolates of mutans streptococci: An in vitro study. Acta Odontol. Scand. 2011, 69, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.C.; Wei, S.M.; Luo, X.T.; Yang, Q.Q.; Wong, K.H.; Cheung, P.C.K.; Zhang, B.B. How probiotics, prebiotics, synbiotics, and postbiotics prevent dental caries: An oral microbiota perspective. NPJ Biofilms Microbiomes 2024, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Vale, G.C.; Mayer, M.P.A. Effect of probiotic Lactobacillus rhamnosus by-products on gingival epithelial cells challenged with Porphyromonas gingivalis. Arch. Oral. Biol. 2021, 128, 105174. [Google Scholar] [CrossRef]
- Laleman, I.; Detailleur, V.; Slot, D.E.; Slomka, V.; Quirynen, M.; Teughels, W. Probiotics reduce mutans streptococci counts in humans: A systematic review and meta-analysis. Clin. Oral. Investig. 2014, 18, 1539–1552. [Google Scholar] [CrossRef]
- Liévin-Le Moal, V.; Servin, A.L. Anti-infective activities of lactobacillus strains in the human intestinal microbiota: From probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef] [PubMed]
- Miyazima, T.Y.; Ishikawa, K.H.; Mayer, M.; Saad, S.; Nakamae, A. Cheese supplemented with probiotics reduced the Candida levels in denture wearers-RCT. Oral. Dis. 2017, 23, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Leonhard, M.; Moser, D.; Schneider-Stickler, B. Inhibition activity of Lactobacilli supernatant against fungal-bacterial multispecies biofilms on silicone. Microb. Pathog. 2017, 113, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Hall, F.G.; Urbizo-Reyes, U.C.; Garcia, H.S.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A.; Liceaga, A.M. In Silico Prediction and In Vitro Assessment of Multifunctional Properties of Postbiotics Obtained From Two Probiotic Bacteria. Probiotics Antimicrob. Proteins 2020, 12, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Xing, D. The Current and Future Perspectives of Postbiotics. Probiotics Antimicro. Prot. 2023, 15, 1626–1643. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.H.; Bueno, M.R.; Kawamoto, D.; Simionato, M.R.L.; Mayer, M.P.A. Lactobacilli postbiotics reduce biofilm formation and alter transcription of virulence genes of Aggregatibacter actinomycetemcomitans. Mol. Oral. Microbiol. 2021, 36, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Mardani, K.; Tajik, H. Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. Lebensm.-Wiss. Technol. 2019, 111, 457–464. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, V.H.; Wang, Y.; Bandara, H.M.H.N.; Mayer, M.P.A.; Samaranayake, L.P. Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotech. 2016, 100, 6415–6426. [Google Scholar] [CrossRef]
- Koohestani, M.; Moradi, M.; Tajik, H.; Badali, A. Effects of cellfree supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet. Res. Forum 2018, 9, 301–306. [Google Scholar]
- Ishikawa, K.H.; Mita, D.; Kawamoto, D.; Nicoli, J.R.; Albuquerque-Souza, E.; Lorenzetti, M.R.; Mayer, M.P.A. Probiotics alter biofilm formation and the transcription of Porphyromonas gingivalis virulence-associated genes. J. Oral. Microbiol. 2020, 20, 1805553. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, Y. A comparative study of the effect of probiotics on cariogenic biofilm model for preventing dental caries. Arch. Microbiol. 2014, 196, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fan, Y.; Wang, X.; Jiang, X.; Zou, J.; Huang, R. Effects of the natural compound, oxyresveratrol, on the growth of Streptococcus mutans, and on biofilm formation, acid production, and virulence gene expression. Eur. J. Oral. Sci. 2020, 128, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Surendran, N.M.; Amalaradjou, M.A.; Venkitanarayanan, K. Antivirulence Properties of Probiotics in Combating Microbial Pathogenesis. Adv. Appl. Microbiol. 2017, 98, 1–29. [Google Scholar]
- Hossain, M.I.; Mizan, F.R.; Roy, P.K.; Nahar, S.; Toushik, S.H.; Ashrafudoulla, M.; Jahid, I.K.; Lee, J.; Ha, S. Listeria monocytogenes biofilm inhibition on food contact surfaces by application of postbiotics from Lactobacillus curvatus B.67 and Lactobacillus plantarum M.2. Food Res. Int. 2021, 148, 110595. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Martínez, M.I.; Suárez, A.M.; Martínez, J.M.; Hernández, P.E. Research note: Unsuitability of the MRS medium for the screening of hydrogen peroxide-producing lactic acid bacteria. Lett. Appl. Microbiol. 1997, 25, 73–74. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.I.; Franco, A.R.; Rodrigues, F.J.; Leonor, I.B.; Reis, R.L. Unveiling the effect of three-dimensional bioactive fibre mesh scaffolds functionalized with silanol groups on bacteria growth. J. Biomed. Mater. Res. A 2016, 104, 2189–2199. [Google Scholar] [CrossRef]
- Mahdizadeh, S.; Sawford, K.; van Andel, M.; Browning, G.F. Efficacy of citric acid and sodium hypochlorite as disinfectants against Mycoplasma bovis. Vet. Microbiol. 2020, 243, 108630. [Google Scholar] [CrossRef]
- Burel, C.; Kala, A.; Purevdorj-Gage, L. Impact of pH on citric acid antimicrobial activity against Gram-negative bacteria. Lett. Appl. Microbiol. 2021, 72, 332–340. [Google Scholar] [CrossRef]
- Diop, M.B.; Alvarez, V.B.; Guiro, A.T.; Thonart, P. Efficiency of Neutralized Antibacterial Culture Supernatant from Bacteriocinogenic Lactic Acid Bacteria Supplemented With Salt in Control of Microorganisms Present in Senegalese Artisanally Handled Fish by Immersion Preservative Technology During Guedj Seafood Processing at 10 °C and 30 °C. J. Food Microbiol. Saf. Hyg. 2016, 1, 1. [Google Scholar] [CrossRef]
- Lin, X.; Chen, X.; Chen, Y.; Jiang, W.; Chen, H. The effect of five probiotic lactobacilli strains on the growth and biofilm formation of Streptococcus mutans. Oral. Dis. 2015, 21, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Söderling, E.M.; Marttinen, A.M.; Haukioja, A.L. Probiotic lactobacilli interfere with Streptococcus mutans biofilm formation in vitro. Curr. Microbiol. 2011, 62, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans—Derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Tahmourespour, A.; Salehi, R.; Kermanshahi, R.K. Lactobacillus Acidophilus-Derived Biosurfactant Effect on GTFB and GTFC Expression Level in Streptococcus mutans Biofilm Cells. Braz. J. Microbiol. 2011, 42, 330–339. [Google Scholar] [CrossRef]
- Kim, M.Y.; Hyun, I.K.; An, S.; Kim, D.; Kim, K.H.; Kang, S.S. In vitro anti-inflammatory and antibiofilm activities of bacterial lysates from lactobacilli against oral pathogenic bacteria. Food Funct. 2022, 13, 12755–12765. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | Primer Sequence (5′-3′) | Reference |
|---|---|---|---|
| gtfB | Production of water-insoluble glucan | 5′-AGC AAT GCA GCC AAT CTA CAA AT-3′ 5′-ACG AAC TTT GCC GTT ATT GTC A-3′ | [23] |
| gtfC | Production of water-soluble and insoluble glucan | 5′-CTC AAC CAA CCG CCA CTG TT-3′ 5′-GGT TAA CGT CAA TAG AAT CTG TAG TAG C-3′ | [23] |
| gtfD | Production of water-soluble glucan | 5′-CAC AGG CAA AAG CTG AAT TAC A-3′ 5′-GAT GGC CGC TAA GTC AAC AG-3′ | [23] |
| vicR | Response Regulator | 5′-TGA CACGA TTA CAG CCT TTG ATG-3′ 5′-CGT CTA GTT CTG GTA ACA TTA AGT CCA ATA-3′ | [24] |
| 16S rRNA | Internal standard normalizer | 5′-AGT CTG AGT 5′-GAA TAA AAG GCT A-3′ 5′-GTT AGC TCC GGC ACT AAG CC-3′ | [23] |
| Nº | RT | Similarity | Constituent | Molecular Formula | Absolute Area | Relative Area (%) |
|---|---|---|---|---|---|---|
| 1 | 10.119 | 96 | Trimethylsilyl Ester of L-valine | C11H27NO2Si2 | 2,565,504 | 1.59% |
| 2 | 15.082 | 94 | Pentanoic acid | C5H10O2 | 712,900 | 0.44% |
| 3 | 16.336 | 94 | Silanol, Phosphoric acid tri-TMS | C9H27O4PSi3 | 28,993,178 | 17.98% |
| 4 | 16.492 | 94 | 3,7-Dioxa-2,8-disilanonane | C9H24O2Si2 | 3,325,745 | 2.06% |
| 5 | 17.056 | 95 | Trimethylsilyl ester of O-trimethylsilyl-L-Threonine | C10H25NO3Si2 | 2,381,219 | 1.48% |
| 6 | 17.821 | 95 | Butanedioic acid | C4H6O4 | 1,288,829 | 0.80% |
| 7 | 24.228 | 94 | L-Proline, Pyroglutamic acid di-TMS | C5H7NO3 | 4,212,701 | 2.61% |
| 8 | 24.978 | 95 | Phenylalanine 1TMS | C12H19NO2Si | 2,606,598 | 1.62% |
| 9 | 31.267 | 90 | Phosphoric acid, α.-Glycer | C3H9O6P | 428,872 | 0.27% |
| 10 | 32.768 | 87 | Citric acid | C6H8O7 | 31,491,768 | 19.53% |
| 11 | 33.372 | 90 | Glucofuranoside, methyl 2,3,5,6-tetrakis-O-(trimethylsilyl) | C19H46O6Si4 | 6,408,426 | 3.98% |
| 12 | 34.591 | 96 | α-D-Galactopyranose | C6H12O6 | 21,217,490 | 13.16% |
| 13 | 36.844 | 96 | β-D-Glucopyranose | C6H12O6 | 28,402,983 | 17.62% |
| 14 | 39.255 | 91 | Myo-inositol | C6H12O6 | 1,018,404 | 0.63% |
| 15 | 41.648 | 90 | Tryptophan 2TMS | C17H28N2O2Si2 | 724,774 | 0.45% |
| 16 | 50.705 | 94 | Lactose-Octarms | C36H86O11Si8 | 12,470,922 | 7.74% |
| 17 | 52.994 | 92 | Maltose-Octarms | C36H86O11Si8 | 12,963,518 | 8.04% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, G.B.; Quelemes, P.V.; da Silva Neta, E.R.; de Lima, S.G.; Vale, G.C. Chemical Characterization and Effect of a Lactobacilli-Postbiotic on Streptococcus mutans Biofilm In Vitro. Microorganisms 2024, 12, 843. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050843
Santana GB, Quelemes PV, da Silva Neta ER, de Lima SG, Vale GC. Chemical Characterization and Effect of a Lactobacilli-Postbiotic on Streptococcus mutans Biofilm In Vitro. Microorganisms. 2024; 12(5):843. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050843
Chicago/Turabian StyleSantana, Guilherme Bandeira, Patrick Veras Quelemes, Enedina Rodrigues da Silva Neta, Sidney Gonçalo de Lima, and Gláuber Campos Vale. 2024. "Chemical Characterization and Effect of a Lactobacilli-Postbiotic on Streptococcus mutans Biofilm In Vitro" Microorganisms 12, no. 5: 843. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms12050843