
Relationship of the “Dequada” Phenomenon with Mercury Methylation in Pantanal, Brazil
 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.2. Collection and Preparation of Sediment and Water Samples
2.3. Physical and Chemical Parameters
2.4. Microcosm Assemblies for the Study of Mercury Methylation Potential
2.5. Determination of Total Hg
2.6. Determination of Methylmercury
2.7. Determination of Total Organic Carbon
3. Results and Discussion
3.1. Results of Two Physical and Chemical Parameters without Water Samples
3.2. Results of Total Hg in Water and Sediment Samples
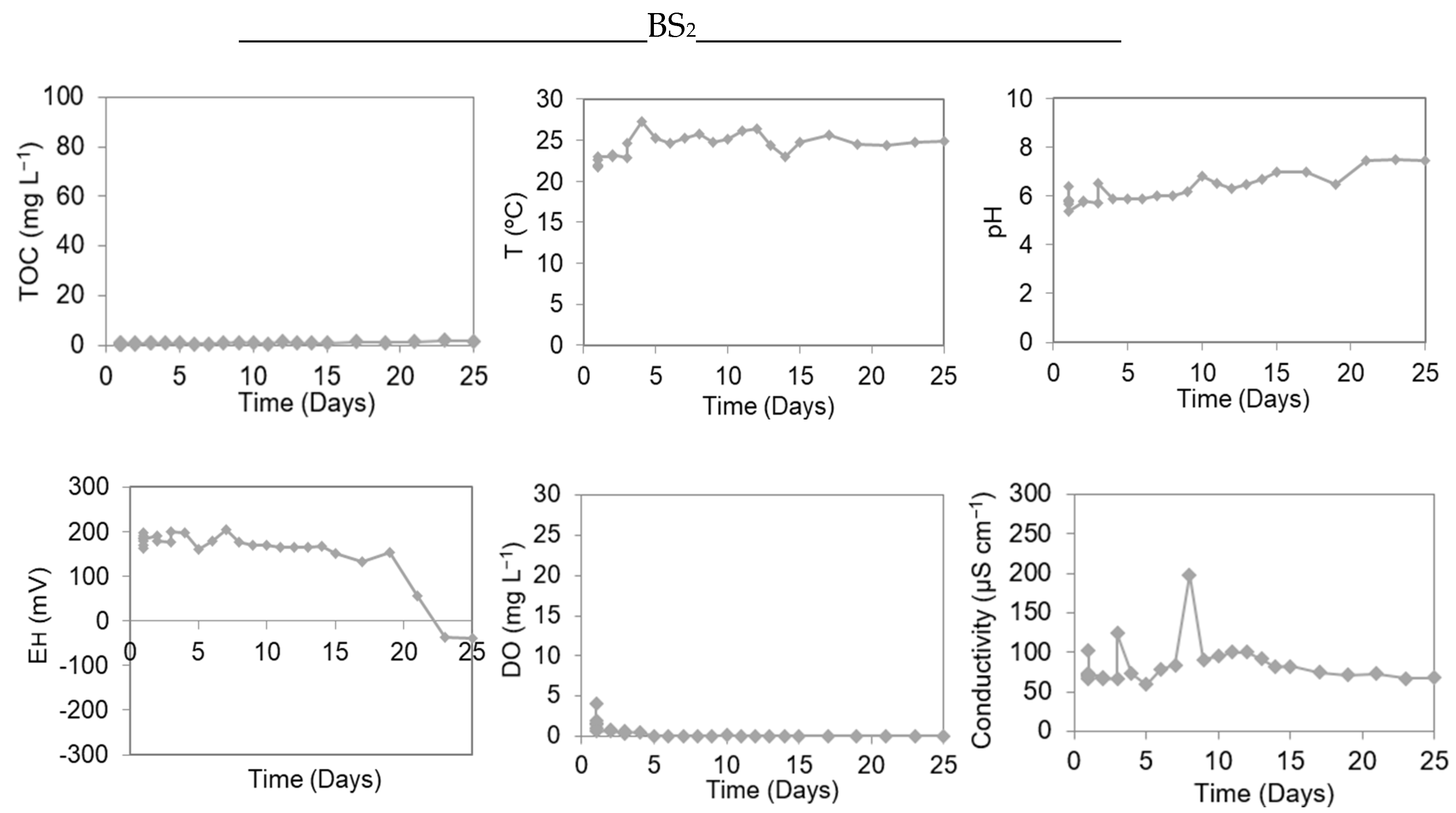
3.3. Methylation Study in Microcosm Experiments Assembled with Sediment and Water from the Castelo Bay
3.4. Methylation Study in Microcosm Experiments Assembled with Sediment and Water from the Saracura Bay
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hamilton, S.K.; Sippel, S.J.; Melack, J.M. Comparison of inundation patterns among major South American floodplains. J. Geophys. Res. Atmos. 2002, 107, LBA 5-1–LBA 5-14. [Google Scholar] [CrossRef]
- Lázaro, W.L.; Guimarães, J.R.D.; Ignácio, A.R.; Da Silva, C.J.; Díez, S. Cyanobacteria enhance methylmercury production: A hypothesis tested in the periphyton of two lakes in the Pantanal floodplain, Brazil. Sci. Total. Environ. 2013, 457, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Schleder, A.; Froehner, S.; Sanez, J.; Parron, L.; Hansel, F.; Guerreiro, R.L.; Bahniuk, A. Insights into the organic matter composition of soda lakes in the Pantanal, Brazil, through fatty acids analysis in sediments. Environ. Sci. Pollut. Res. 2023, 30, 103932–103946. [Google Scholar] [CrossRef] [PubMed]
- Calheiros, D.; Seidl, A.; Ferreira, C. Participatory research methods in environmental science: Local and scientific knowledge of a limnological phenomenon in the Pantanal wetland of Brazil. J. Appl. Ecol. 2000, 37, 684–696. [Google Scholar] [CrossRef]
- Stevaux, J.C.; Macedo, H.d.A.; Assine, M.L.; Silva, A. Changing fluvial styles and backwater flooding along the Upper Paraguay River plains in the Brazilian Pantanal wetland. Geomorphology 2019, 350, 106906. [Google Scholar] [CrossRef]
- Calheiros, D.F.; Ferreira, C.J.A. Alterações Limnológicas no rio Paraguai (“Dequada”) e o Fenômeno Natural de Mortandade de Peixes no Pantanal Mato-Grossense–Corumbá/MS. EMBRAPA-CPAP. 1996, pp. 48–51. Available online: https://www.embrapa.br/en/busca-de-publicacoes/-/publicacao/792189/alteracoes-limnologicas-no-rio-paraguai-dequada-e-o-fenomeno-natural-de-mortandade-de-peixes-no-pantanal-mato-grossense---ms (accessed on 8 February 2024).
- Coringa, J.D.E.S.; Pezza, L.; Coringa, E.d.A.O.; Weber, O.L.d.S. Distribuição geoquímica e biodisponibilidade de metais traço em sedimentos no Rio Bento Gomes, Poconé—MT, Brasil. Acta Amaz. 2016, 46, 161–174. [Google Scholar] [CrossRef]
- Bisinoti, M.C.; Jardim, W.F. O comportamento do metilmercúrio (metilHg) no ambiente. Quimica Nova 2004, 27, 593–600. [Google Scholar] [CrossRef]
- Schwieger, A.-C.; Gebauer, K.; Ohle, A.; Beckmann, M. Determination of mercury binding forms in humic substances of lignite. Fuel 2020, 274, 117800. [Google Scholar] [CrossRef]
- Jiang, P.; Liu, G.; Cui, W.; Cai, Y. Geochemical modeling of mercury speciation in surface water and implications on mercury cycling in the everglades wetland. Sci. Total. Environ. 2018, 640, 454–465. [Google Scholar] [CrossRef]
- Miranda, M.R.; Coelho-Souza, S.A.; Guimarães, J.R.D.; Correia, R.R.S.; Oliveira, D. Mercúrioemsistemas aquáticos: Fatores ambientais que afetam a metilação. Oecologia Bras. 2007, 11, 240–251. [Google Scholar] [CrossRef]
- Hamelin, S.; Amyot, M.; Barkay, T.; Wang, Y.; Planas, D. Methanogens: Principal methylators of mercury in lake periphyton. Environ. Sci. Technol. 2011, 45, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zakem, E.J.; Dutkiewicz, S.; Zhang, Y. Biomagnification of methylmercury in a marine plankton ecosystem. Environ. Sci. Technol. 2020, 54, 5446–5455. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, F.; Silva, E.C.E.; Junk, W. Mercury from gold minings in the Pantanal of Pocone (Mato Grosso, Brazil). Int. J. Environ. Health Res. 1997, 7, 181–192. [Google Scholar] [CrossRef]
- Calheiros, D.F. Influência do Pulso de Inundação na Composição Isotópica (d13c e d15n) das Fontes Primárias de Energia na Planície de Inundação do rio Paraguai (Pantanal/MS); University Paulista: São Paulo, Brazil, 2004; pp. 30–35. [Google Scholar] [CrossRef]
- Gomes, V.M.; dos Santos, A.; Zara, L.F.; Ramos, D.D.; Forti, J.C.; Ramos, D.D.; Santos, F.A. Study on Mercury Methylation in the Amazonian Rivers in Flooded Areas for Hydroelectric Use. Water Air Soil Pollut. 2019, 230, 211. [Google Scholar] [CrossRef]
- Rozas, O.; Vidal, C.; Baeza, C.; Jardim, W.F.; Rossner, A.; Mansilla, H.D. Organic micropollutants (OMPs) in natural waters: Oxidation by UV/H2O2 treatment and toxicity assessment. Water Res. 2016, 98, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Bisinoti, M.C.; Jardim, W.F.; Júnior, J.L.B.; Malm, O.; Guimarães, J.R. Um novo método para quantificar mercúrio orgânico (Hg orgânico) empregando a espectrometria de fluorescência atômica do vapor frio. Quimica Nova 2006, 29, 1169–1174. [Google Scholar] [CrossRef]
- Santos, F.A.; Cavecci, B.; Vieira, J.C.S.; Franzini, V.P.; Santos, A.; Leite, A.d.L.; Buzalaf, M.A.R.; Zara, L.F.; Padilha, P.d.M. A Metalloproteomics study on the association of Mercury with breast milk in samples from lactating women in the Amazon region of Brazil. Arch. Environ. Contam. Toxicol. 2015, 69, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Hintelmann, H.; Falter, R.; Ilgen, G.; Evans, R.D. Determination of artifactual formation of monomethylmercury (CH 3 Hg + ) in environmental samples using stable Hg2+ isotopes with ICP-MS detection: Calculation of contents applying species specific isotope addition. Fresenius J. Anal. Chem. 1997, 358, 363–370. [Google Scholar] [CrossRef]
- Effler, S.W.; Bloom, N.S. Seasonal variability in the mercury speciation of Onondaga Lake (New York). Water Air Soil Pollut. 1990, 53, 251–260. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, L.; Wang, Y.; Gao, S.; Daut, G. A comparison of different methods for determining the organic and inorganic carbon content of lake sediment from two lakes on the Tibetan Plateau. Quat. Int. 2012, 250, 49–54. [Google Scholar] [CrossRef]
- Vanhala, P.; Karhu, K.; Tuomi, M.; Sonninen, E.; Jungner, H.; Fritze, H.; Liski, J. Old soil carbon is more temperature sensitive than the young in an agricultural field. Soil Biol. Biochem. 2007, 39, 2967–2970. [Google Scholar] [CrossRef]
- Yousef, E.A.; El-Mallah, A.M.; Abdel-Baki, A.-A.S.; Al-Quraishy, S.; Reyad, A.; Abdel-Tawab, H. Efeito das Variáveis Ambientais no Zooplâncton em Vários Habitats do Rio Nilo. Água 2024, 16, 915. [Google Scholar] [CrossRef]
- Stein, E.D.; Cohen, Y.; Winer, A.M. Environmental distribution and transformation of mercury compounds. Crit. Rev. Environ. Sci. Technol. 1996, 26, 1–43. [Google Scholar] [CrossRef]
- Li, Y.; Cai, Y. Progress in the study of mercury methylation and demethylation in aquatic environments. Chin. Sci. Bull. 2012, 58, 177–185. [Google Scholar] [CrossRef]
- Xun, L.; Campbell, N.E.R.; Rudd, J.W.M. Measurements of Specific Rates of Net Methyl Mercury Production in the Water Column and Surface Sediments of Acidified and Circumneutral Lakes. Can. J. Fish. Aquat. Sci. 1987, 44. [Google Scholar] [CrossRef]
- Zhang, D.; Xiang, Y.; Liu, G.; Liang, L.; Chen, L.; Shi, J.; Yin, Y.; Cai, Y.; Jiang, G. Mechanism and controlling factors on rapid methylmercury degradation by ligand-enhanced Fenton-like reaction at circumneutral pH. Chemosphere 2023, 324, 138291. [Google Scholar] [CrossRef]
- Zhao, L.; Guo, Y.; Meng, B.; Yao, H.; Feng, X. Effects of damming on the distribution and methylation of mercury in Wujiang River, Southwest China. Chemosphere 2017, 185, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Hudelson, K.E.; Drevnick, P.E.; Wang, F.; Armstrong, D.; Fisk, A.T. Mercury methylation and demethylation potentials in Arctic lake sediments. Chemosphere 2020, 248, 126001. [Google Scholar] [CrossRef]
- Wright, D.R.; Hamilton, R.D. Release of Methyl Mercury from Sediments: Effects of Mercury Concentration, Low Temperature, and Nutrient Addition. Can. J. Fish. Aquat. Sci. 1982, 39, 1459–1466. [Google Scholar] [CrossRef]
- Lázaro, W.L.; Oliveira-Júnior, E.S.; da Silva, C.J.; Castrillon, S.K.I.; Muniz, C.C. Climate change reflected in one of the largest wetlands in the world: An overview of the Northern Pantanal water regime. Acta Limnol. Bras. 2020, 32, e104. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Point | Period | Water Column (m) | Reference Coordinate |
|---|---|---|---|
| P1 Castelo bay | 1 (flood period) 2 (drought period) | 8.0 5.5 | 18°35.580′ S 57°14.233′ W |
| P2 Castelo bay | 1 (flood period) 2 (drought period) | 8.5 6.0 | 18°35.597′ S 57°32.929′ W |
| P3 Castelo bay | 1 (flood period) 2 (drought period) | 9.5 7.5 | 18°35.334′ S 57°32.728′ W |
| P6 Saracura bay | 1 (flood period) 2 (drought period) | 3.0 1.5 | 18°42.252′ S 57°35.803′ W |
| P7 Saracura bay | 1 (flood period) 2 (drought period) | 3.5 2.0 | 18°43.337′ S 57°36.683′ W |
| P8 Saracura bay | 1 (flood period) 2 (drought period) | 2.8 1.5 | 18°43.410′ S 57°35.154′ W |
| Point | Sampling | T (°C) | pH | DO (mg L−1) | EH (mV) | Condut. (μS cm−1) |
|---|---|---|---|---|---|---|
| P1 | 1 2 | 26.18 23.00 | 5.04 5.78 | 1.49 7.50 | 79.50 65.15 | 66 177 |
| P2 | 1 2 | 26.18 23.15 | 5.25 5.64 | 1.49 6.27 | 79.50 65.15 | 66 151 |
| P3 | 1 2 | 26.18 25.05 | 5.30 5.85 | 1.49 6.45 | 79.50 65.15 | 66 89 |
| P6 | 1 2 | 26.38 25.34 | 5.90 5.95 | 0.72 5.87 | 38.90 22.00 | 60 92 |
| P7 | 1 2 | 26.38 25.10 | 6.02 6.01 | 0.72 4.79 | 38.90 22.00 | 60 94 |
| P8 | 1 2 | 26.38 25.00 | 6.02 6.11 | 0.72 5.15 | 38.90 22.00 | 60 90 |
| Point | Sampling | [Hgtotal] (ng L−1) Water | [Hgtotal] (ng g−1) Sediment |
|---|---|---|---|
| P1 | 1 2 | 6.78 ± 0.04 <LQ | 56.24 ± 0.19 19.51 ±0.15 |
| P2 | 1 2 | <LQ <LQ | 65.08 ± 0.16 12.45 ± 0.08 |
| P3 | 1 2 | <LQ <LQ | 64.57 ± 0.21 7.81 ± 0.04 |
| P6 | 1 2 | <LQ <LQ | 43.33 ± 0.15 30.98 ± 0.11 |
| P7 | 1 2 | <LQ <LQ | 45.77 ± 0.18 17.58 ± 0.16 |
| P8 | 1 2 | <LQ <LQ | 46.46 ± 0.15 28.33 ± 0.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, G.A.; Ramos, D.D.; Forti, J.C.; Ramos, D.D.; Oliveira, L.C.S.; Junior, D.S.S.; Santo, G.E.; Santos, F.A. Relationship of the “Dequada” Phenomenon with Mercury Methylation in Pantanal, Brazil. Water 2024, 16, 1204. https://0-doi-org.brum.beds.ac.uk/10.3390/w16091204
Nunes GA, Ramos DD, Forti JC, Ramos DD, Oliveira LCS, Junior DSS, Santo GE, Santos FA. Relationship of the “Dequada” Phenomenon with Mercury Methylation in Pantanal, Brazil. Water. 2024; 16(9):1204. https://0-doi-org.brum.beds.ac.uk/10.3390/w16091204
Chicago/Turabian StyleNunes, Glaucia A., Dayana D. Ramos, Juliane C. Forti, Diovany D. Ramos, Lincoln C. S. Oliveira, David S. S. Junior, Gabriel E. Santo, and Felipe A. Santos. 2024. "Relationship of the “Dequada” Phenomenon with Mercury Methylation in Pantanal, Brazil" Water 16, no. 9: 1204. https://0-doi-org.brum.beds.ac.uk/10.3390/w16091204