1. Introduction
Muscle satellite cells (MuSCs) are a major cell type involved in skeletal muscle regeneration and are essential for the growth, maintenance, and repair of skeletal muscle postnatally [
1,
2,
3]. These cells exist in a quiescent state between the sarcolemma and the basal lamina of muscle fibers, exhibiting heterogeneity [
2]. In mature resting muscle, MuSCs mainly remain in a quiescent state. However, upon muscle damage, these cells are activated; and undergo self-renewal, proliferation, differentiation, and fusion. Ultimately, they will form new muscle fibers and repair damaged muscle tissue [
4,
5,
6].
Research has shown that paired box protein 7 (
PAX7) serves as a marker gene and important regulatory factor for satellite cells [
7]. The differentiation ability of MuSCs is controlled by myogenic regulatory factors (MRFs), such as myogenic factor 5 (MYF5), myogenic differentiation factor (MYOD), myogenic regulatory factor 4 (MRF4), and myogenin (MYOG) [
8]. MYOG is responsible for driving terminal differentiation and activating the expression of the target genes necessary for this process [
9]. Upon activation, the expression of
MYF5 and
MYOD rapidly increased [
10,
11]. Subsequently, myogenic cells enter the final differentiation stage, expressing
MYOG,
MRF4, and other genes specific to mature muscle fibers, such as myosin heavy chain (
MyHC). Therefore, the expression of
PAX7,
MYOD, and
MYOG can be used to distinguish different stages of MuSCs, including
PAX7+
MYOD- (quiescent/self-renewal),
PAX7+
MYOD+ (proliferation), and
PAX7-MYOD+
MYOG+ (differentiation) [
12,
13].
Pigs have many advantages in modeling human diseases due to their similar anatomic and physiological features to human beings. Pig MuSCs (pMuSCs) are crucial materials for studying skeletal muscle development and regeneration and also can be used to produce cultured meat [
14]. The characterization of MuSCs in pigs will be a valuable addition to our understanding of the porcine model system. Like MuSCs from other species, pig MuSCs also lose their proliferation and differentiation abilities during long-term in vitro culture [
15]. Furthermore, the isolation of primary cells is associated with drawbacks such as time consumption, labor-intensive procedures, and significant inter-individual variations. Researches on pig muscle often limited by using primary MuSCs. Therefore, it is necessary to establish porcine muscle satellite cells capable of long-term stable passaging in vitro.
One common method to inhibit cell apoptosis is using of simian virus 40 large T-antigen (SV40 T-antigen) expression constructs [
16]. In in vitro cell cultures, SV40 T-antigen can facilitate the transformation of oncogenes. The T-antigen interacts with tumor-suppressor proteins Rb (retinoblastoma) and p53, forming complexes with the Rb-binding domain and p53-binding domain. This interaction inhibits the tumor-suppressing effects of Rb and p53, thereby relieving their inhibition on the cell cycle. This leads to accelerated cell division and unlimited growth [
17]. In most cases, the T-antigen induces the inactivation of Rb protein, leading to the activation of E2F-dependent transcription and subsequent entry into the S-phase. Additionally, the antigen inhibits the activity of p53, thereby preventing subsequent cell cycle arrest and apoptosis [
18].
In recent years, scientists have developed various immortalized cell lines. The currently known immortalized pig cell lines mainly comprise the following: PK15 (Porcine Kidney 15), a commonly used pig kidney cell line that is widely applied in virus research, vaccine production, and biopharmaceutical fields [
19]. LLC-PK1 (Lilly Laboratory Porcine Kidney 1), derived from pig kidney tissue, is frequently utilized to investigate drug metabolism, toxicity, and cellular biology [
20]. IPEC-J2 (Intestinal Porcine Epithelial Cell-J2), derived from pig small intestinal epithelial cells, is commonly employed to study pig intestinal physiology, diseases, and nutrition [
21]. In addition to these cell lines, there are also other cell lines utilized in pig including pig embryonic cell lines, fibroblast cell lines, and lymphoblast cell lines [
22,
23]. These cell lines play a significant role in studying various aspects of pig health, reproduction, immunity, and nutrition. However, no relevant reports on pig satellite cell lines have been found to date.
In this study, primary MuSCs was isolated from a 1-day-old Western commercial pig’s longissimus dorsi muscle using the differential adhesion method. Subsequently, a pig MuSCs line was established by transducing the SV40 T-antigen. The established SV40 T-antigen pig muscle satellite cells (SV40 T-pMuSCs) could provide a valuable source of cells for studying pig skeletal muscle development and differentiation.
2. Materials and Methods
2.1. Pig Primary MuSCs Isolation and Differentiation
Pig muscle primary satellite cells (pMuSCs) were isolated from the longissimus dorsi muscle of one 1-day-old Western commercial pig using the differential adhesion method. Briefly, the longissimus dorsi muscle on both sides was minced and incubated with 0.2% type I collagenase (Sigma, Darmstadt, Germany, Cat# SCR103) (twice the volume) at 37 °C and 100 rpm in a water bath for 2–2.5 h until the solution became a yellow viscous substance. After centrifugation to remove the collagenase I, the cell pellet was resuspended in 0.25% trypsin–EDTA (Gibco, Glen Island, New York, NY, USA, Cat# 25200072) (three times the volume) and digested at 37 °C and 100 rpm for 30 min. An equal volume of growth medium (GM) was added to stop the digestion, and the cells were successively filtered through 100 μm and 70 μm cell strainers. The GM medium consisted of DMEM (Gibco, Carlsbad, CA, USA, Cat# 6123118) supplemented with 10% Fetal Bovine Serum (FBS) (Dcell, Darmstadt, Germany, Cat# 23055525) and 1% penicillin–streptomycin (Gibco, Carlsbad, CA, USA, Cat# 15140122). The supernatant was removed by centrifugation at 2000 rpm for 12 min. The cells were resuspended in PBS, centrifuged at 1500 rpm for 10 min to remove the PBS, and this step was repeated twice. Finally, the cells were resuspended in complete GM medium and incubated for 2 h on three of 25 cm2 cell culture flasks. The complete GM medium consisted of F12 (Gibco, Grand Island, NE, USA, Cat# 11330032) supplemented with 20% FBS (Dcell, Darmstadt, Germany, Cat# 23055525), 1% penicillin–streptomycin (Gibco, Carlsbad, CA, USA, Cat# 15140122), 1% GlutaMAX (Gibco Grande Island, New York, NY, USA Cat# 35050079), 1% sodium pyruvate (Gibco, Grande Island, NY, USA, Cat# 11360070), 1% non-essential amino acid (Gibco, Grande Island, NY, USA, Cat# 11140050), and 2 ng/mL bFGF (Peprotech, Rocky Hill, NJ, USA, Cat# 450-33). The supernatant was collected as satellite cells and cultured in 37 °C, 25 cm2 cell culture flasks, with the medium being replaced every 48 h. The pMuSCs were differentiated for 5 days, the differentiation medium consisted of DMEM supplemented with 0.4% Ultroser G (SARTORIUS, Berlin, Germany, Cat# 15950-017) and 1% penicillin–streptomycin (Gibco, Carlsbad, CA, USA, Cat# 15140122). For the statistical analysis, the differentiation index is defined as the percentage of MyHC+ nuclei compared to the total number of nuclei, while the myotube fusion index is defined as the ratio of the total number of nuclei within myotubes (consisting of three or more nuclei) to the total number of MyHC+ nuclei.
2.2. Immunofluorescence Staining
The cells with designated generation were plated in a 24-well plate, and the experiment was initiated once the confluence reached 50%. Firstly, the cell culture medium was removed, and the cells were washed with PBS and fixed for 30 min using 4% paraformaldehyde. Then, the cells were permeabilized with 0.3% Triton X-100 for 10 min and washed three times with PBS. Quick Blocking solution (Beyotime, Shanghai, China, Cat# P0260) was applied and incubated for 1 h. The primary antibody was added and incubated overnight at 4 °C, followed by PBS wash for three times. The secondary antibody was added and incubated at room temperature in dark for 1 h. After removing the secondary antibody, the cells were washed with PBS three times, and the cell nuclei were stained using anti-fade mounting medium with DAPI (Bioshrap, Shanghai, China, Cat# BL739A). The experiments were repeated three times. Images were captured using a fluorescence Nexcope NIB610-FL microscope, and representative images were shown. Primary antibodies used in this study include anti-MyHC (MF-20, 1:100) (Developmental Studies Hybridodma Bank, Ithaca, New York, NY, USA, Cat# AB_2147781), anti-PAX7 (Developmental Studies Hybridodma Bank, Ithaca, New York, NY, USA, Cat# AB_528428), and anti-SV40 T-antigen (Abcam, Cambridge, UK, Cat# ab234426). Secondary antibodies used in this study include Cy3-labeled goat anti-mouse (1:1000) (Beyotime, Shanghai, China, Cat# A5021), Alexa Fluor 488-labeledgoat anti-mouse (Beyotime, Shanghai, China, Cat# A0428), and Alexa Fluor 488-labeled goat anti-rabbit (Beyotime, Shanghai, China, Cat# A0423).
2.3. Real-Time Quantitative PCR (qPCR)
The total RNAs were extracted using TRIzol (Ambion, Carlsbad, CA, USA, Cat# 380512). cDNA was synthesized using an Evo M-MLV Mix Tracking Kit with gDNA Clean (Accurate, Shanghai, China, Cat# AG11734), following the manufacturer’s instructions. qPCR was performed using Hieff UNICON Universal Blue qPCR SYBR Green Master Mix (Yeasen, Shanghai, China, Cat# 11141ES) on a CFX96 Optics Module. The PCR program is as follows: pre-denaturation—95 °C, 2 min; denaturation—95 °C, 10 s; and annealing—60 °C, 30 s. Cycle number is 40. The primers used in these study were as follows: PAX7-F, 5′-GTGCCCTCAGTGAGTTCGATT-3′, PAX7-R, 5′-TCCAGACGGTTCCCTTTGTC-3′; MYOD-F, 5′- GCTCCGCGACGTAGATTTGA-3′, MYOD-R, 5′-GGAGTCGAAACACGGGTCAT-3′, SV40 T-antigen-F: GTGCCTAAAACACTGCAGGC, SV40 T-antigen-R: CAGCCACAGGTCTGTACCAA. GAPDH-F, 5′-TGAGATCAGGGAGCCATCA-3′, GAPDH-R, 5′ATGGTCAGGGGTCCGAT-GTA-3′. The relative expression levels of the target gene were calculated using the 2−ΔΔCt method. The experimental results are presented as the mean ± standard error (SEM). Statistical analysis and tests for significant differences among groups were performed using IBM SPSS Statistics 26 software.
2.4. Transduction of SV40 T-Antigen to pMuSCs
Lentiviral particles containing the SV40 T-antigen were obtained using human 293T cells. The 293T cells reached 70% density in a six-well plate were transfected with a lipofectamine 2000 DNA reagent. LV-EF1a-SV40-Large-T-Antigen-P2A-Hygro-Barcode (Addgene, Watertown, MA, USA, Cat# 170255) (1 μg), psPAX2 (Addgene, Watertown, MA, USA, Cat# 12260) (750 ng), and pMD2.G (Addgene, Watertown, MA, USA, Cat# 12259) (250 ng) were co-transfected into the cells. The supernatants from the cells were collected at both 24 h and 48 h, and the supernatants can be used as lentiviral particle solutions after filtration through a 0.45 μm filter, and then stored at −80 °C for later use. pMuSCs were grown in a six-well plate to reach 20% density, then added 2 mL of complete GM medium with 500 μL SV40 T lentiviral particles and 8 μg/mL polybrene. After 24 h of infection, the medium was changed, and the cells were allowed to rest for one day. Subsequently, the cells were cultured in complete GM medium containing 200 μg/mL hygromycin B for 7 days to select the positive cells expressing SV40 T-antigen (SV40T-pMuSCs).
2.5. EdU Assay
EdU assay was conducted using pMuSCs and SV40 T-pMuSCs at different passages. The 5-ethynyl-2′-deoxyuridine (EdU) assay was performed according to the manufacturer’s protocol (Beyotime, Shanghai, China, Cat# C0078S). The cells were cultured in 12-well plates. The experiment was initiated when the cell confluence reached 50%. A 2× working solution of EdU was prepared in complete GM from the 10 mM stock solution. Then, 500 μL 2× EdU working solution was added to each well, resulting in 1× EdU (10 μM). After 2 h of incubation, cells were fixed by 4% paraformaldehyde (PFA) for 15 min. Subsequently, cells were permeabilized in 0.3% Triton X-100 in PBS for 15 min and stained with Click reaction solution for 30 min. After washing, the nuclei were stained with DAPI for 10 min. Cells were imaged with a Nexcope NIB610-FL microscope. Around 1000 randomly elected cells were analyzed.
2.6. SA-β-Gal Activity
The SA-β-gal activity was assessed in cells from different passages, including pMuSCs (P1, P2, P4, and P10) and SV40 T-pMuSCs (P1, P10, P30, and P40). A senescence β-Galactosidase Staining Kit (Beyotime, Shanghai, China, Cat# C0602) was used according to the manufacturer’s instruction. The cells were cultured in 12-well plates and fixed with stationary liquid for 15 min. Subsequently, cells were washed once with PBS. A volume of 500 µL β-galactosidase staining solution was added, and the cells were incubated overnight at 37 °C in a CO2-free chamber. The resulting images were directly captured using a Nexcope NIB610-FL microscope.
2.7. RNA-seq Analysis
RNA samples were collected from P1, P2, and P4 generations of pMuSCs using TRIzol. RNA-seq libraries were generated by Nanopore Co., Ltd. (Wuhan, China). The libraries were sequenced using the Illumina NovaSeq 6000. For the data analysis, fastp (v0.23.2) was used for quality filtering of the sequencing reads, and reads with unknown sequences >10% and quality scores <20 were removed. STAR (v2.7.9) was used for a fast and accurate sequence aligned to the Suscrofa11.1 reference genome. Finally, a transcriptome gene expression count file was converted using feature Counts (v2.0.3) to obtain the gene expression profile in each sample. Differentially expressed genes were identified by DESeq2 (v.1.20). Genes with a corrected p-value < 0.05 and log2 fold changes > 1 were assigned as significantly differentially expressed.
2.8. SiRNA Transfection
The siRNAs were designed and synthesized by Gene Pharma. Cells were plated into two 12-well plates for the siRNA interference assay, with three replicates per group. The experiment was initiated when the cells reached a confluence of 50%. SV40 T-pMuSCs were transfected with either 50 nM siRNA (si-SV40 T) or control (si-NC) using Lipofectamine RNAiMAX (Invitrogen, Carlsbad, CA, USA, Cat# 13778150). Cells were collected 48 h post-transfection, and RNAs were extracted using the aforementioned method and reverse transcribed for qPCR. After 48 h of transfection, the differentiation medium was replaced, and cells were induced to differentiate for 3 days. The sequences of the siRNAs used were as follows: si-NC sense (5′-3′): UUCUCCGAACGUGUCACGUTT, si-NC antisense (5′-3′): ACGUGACACGUUCGGAGAATT si-SV40 T-antigen-1 sense (5′-3′): GGAGUUUCAUCCUGAUAAATT, si-SV40 T-antigen-1 antisense (5′-3′): UUUAUCAGGAUGAAACUCCTT; si-SV40 T-antigen-2 sense (5′-3′): GUGGUGGAAUGCCUUUAAUTT, si-SV40 T-antigen-2 antisense (5′-3′): AUUAAAGGCAUUCCACCACTT.
2.9. Statistical Analysis
All data were expressed as the mean ± standard error of mean (SEM). Statistical analysis was performed using GraphPad Prism 8. For comparisons of two treatment groups, a t-test was used. For more than two groups, one-way ANOVA with Tukey’s multiple comparison tests was used. Statistical significance was set at * p < 0.05 or ** p < 0.01.
4. Discussion
Muscle satellite cells were initially discovered by Mauro in 1961 and were believed to play a role in skeletal muscle growth and regeneration [
1]. Muscle satellite cells, responsible for muscle regeneration, serve as crucial materials for studying skeletal muscle development and regeneration. It has been reported that the regenerative capacities of mouse [
25], canine [
26], and human [
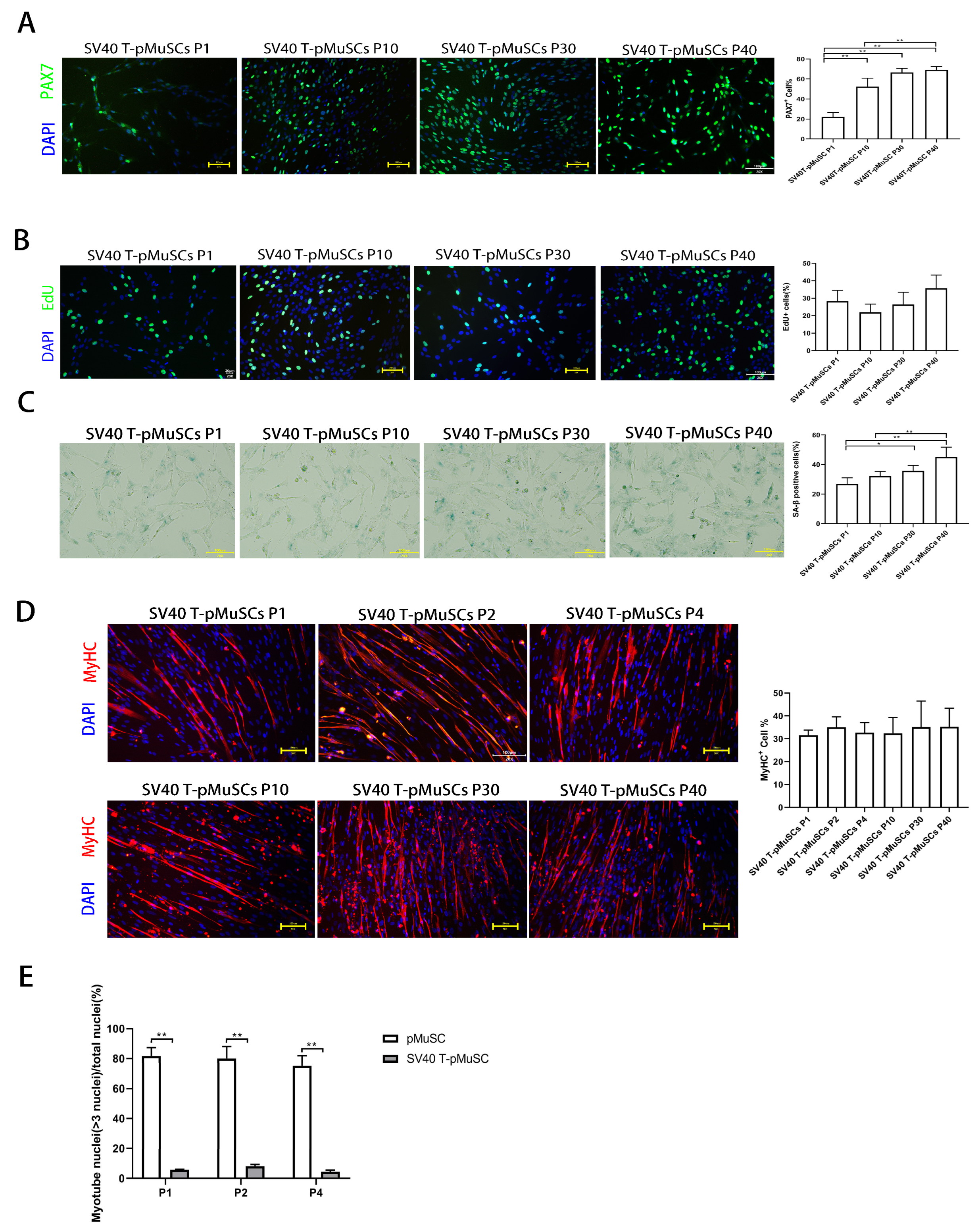
27] primary muscle satellite cells lost their stemness during the in vitro culture process. In this study, we also discovered that pig primary muscle satellite cells (pMuSCs) gradually lose their stemness during long-term in vitro passaging. By transducing lentiviral vectors expressing SV40 T-antigen, we were able to maintain the stemness of pMuSCs. However, compared to pMuSCs, SV40 T-pMuSCs exhibited a noticeable decrease in their differentiation capacity, and interference with the SV40 T-antigen gene could restore the normal differentiation ability.
Obtaining a sufficient number of functional pMuSCs requires significant labor and time, thus limiting studies involving pMuSCs. To understand the mechanism underlying the loss of stemness in satellite cells, we conducted a transcriptomic analysis of satellite cells at different passages. The transcriptional patterns were similar between primary satellite cells at P1 and P2 generations. However, a noticeable change in the transcriptional pattern was observed when comparing P4 with P1, and P4 with P2 generations (
Figure 2A–C). Moreover, there was a significant reduction of the differentiation capacity of P10 generation pMuSCs compare with other early generations (
Figure 1C). These results indicated that one should use lower passaging primary satellite cells for studying the pig’s muscle development and differentiation.
Based on the transcriptomic results, we found that, in low-passage satellite cells, were mainly enriched in pathways such as MAPK signaling, PI3K-Akt signaling, and FoxO signaling. High-passage satellite cells showed a decline in abilities related to DNA replication, base excision repair, and mismatch repair. The cellular response to external stimuli requires the integration and activation of intracellular signaling pathways. These pathways largely determine post-translational modifications of proteins. Among them, phosphorylation being particularly important. One crucial pathway in intracellular signaling is the activation of mitogen-activated protein kinases (MAPKs), which participate in most cellular signaling pathways [
28]. The forkhead box transcription factor (FoxO) family acts downstream of the PI3K/Akt pathway and regulates various physiological processes, including cell proliferation, survival, and metabolism [
29]. It has been discovered that FoxO3 regulates the Notch signaling pathway, which is an important regulator of quiescence in adult satellite cells. The loss of FoxO3 results in decreased Notch signaling levels in satellite cells, indicating the significance of the FoxO3–Notch axis in promoting quiescence during self-renewal processes in satellite cells. Moreover, p38 is a subgroup of the MAPK family, and the p38 pathway is a major regulator of satellite cell fate determination, exerting positive effects at each stage of myogenesis [
30]. The p38α/β MAPK signaling pathway orchestrates the asymmetric division of MuSCs. In this process, one daughter cell activates the p38α/β MAPK cascade, which in turn induces the expression of
MYOD and leads to the generation of proliferative myoblasts. Conversely, in the other daughter cell, the p38α/β MAPK pathway remains inactive, and
MYOD expression is not triggered, resulting in the production of quiescent MuSCs that help to preserve the stem cell reservoir [
31]. The PI3K/AKT signaling pathway mediates the response of satellite cells to growth factors such as IGF1, which promotes key events in the regeneration process, including proliferation, muscle gene expression, myoblast fusion, survival, and post-mitotic growth of myotubes [
32,
33]. In contrast, high-generation satellite cells exhibit a decline in abilities such as DNA replication, base excision repair, and mismatch repair. DNA repair systems have evolved in eukaryotes to overcome DNA damage and include processes such as homologous recombination, non-homologous end joining, base excision repair, and nucleotide excision repair [
34]. Studies have shown that factors involved in DNA repair also regulate cellular metabolism in response to DNA damage, thereby avoiding further genomic instability [
35,
36]. Ineffective DNA repair processes generate DNA fragments that enhance aging, which can be accelerated in cells lacking the homologous recombination repair system [
37]. In summary, skeletal muscle satellite cells exhibit an elevated expression of MAPK signaling, PI3K-Akt signaling, and FoxO signaling at low passage numbers, which promote cell proliferation ability. However, as the cells undergo successive passages, there is a downregulation of DNA replication, base excision repair, and mismatch repair pathways. These findings suggest that prolonged in vitro cell culture weakens DNA repair capabilities, which may contribute to the decreased proliferative and differentiation potential of satellite cells.
To overcome the decline in stemness of muscle satellite cells during long-term in vitro culture, we transduced pMuSCs with a lentiviral vector-expressing SV40 T-antigen. The results showed that the proliferation capacity of SV40 T-pMuSCs was maintained after SV40 T-antigen expression (
Figure 3B). The binding of the SV40 T-antigen to the Rb protein inhibits pathways involved in cell cycle entry and growth arrest controlled by the Rb-E2F complex. E2F is a transcription factor that, together with Rb, regulates the transcription of E2F target genes, which encoded proteins required for DNA replication, nucleotide metabolism, DNA repair, and cell cycle progression. Disruption of the inhibitory effect of the SV40 T-antigen on the Rb-E2F complex leads to the transcription of E2F-dependent genes and entry into the S-phase, enabling continuous cell proliferation [
18]. The SV40 T-antigen interacts with the tumor-suppressor protein p53, a transcriptional activator that mediates apoptosis under adverse conditions, thereby inhibiting cell death [
38].
In addition, we found that the proportion of PAX7+ cells was initially low in SV40 T-pMuSCs at P1, and it gradually approached 60% with subsequent cell passages, similar to pMuSCs at P1 (
Figure 3A). This may be due to the immediate suboptimal state of the cells upon SV40 T-antigen lentivirus infection, as the cells underwent 7 days of selection with hygromycin B, which could affect their normal gene expression patterns. Note that, when we performed the PAX7 immunostaining using P1-generation SV40 T-pMuSCs, we found that, the cell density was low, which could lead to a lower percentage of PAX7+ cells. This is consistent with the findings of Zheng et al., who demonstrated a significant increase in
PAX7 mRNA expression in satellite cells under high-density culture conditions [
39]. As the cells continued to sub-culture and the cell density increased, their cellular state recovered, and the PAX7+ rate was also restored
Figure 3A). Immunofluorescence staining of MyHC in differentiated SV40 T-pMuSCs at different generations revealed that the cells maintained their differentiation capacity (
Figure 3D). However, the number of MyHC+ cells and the myotube fusion index were lower than that of pMuSCs at P1, suggesting that the SV40 T-antigen may affect the differentiation potential of pMuSCs. This result is consistent with the findings of Klundert et al., who discovered that the transformation of primary hamster myoblasts with the SV40 T-antigen led to the inhibition of terminal differentiation. This inhibition was associated with the transcriptional blockade of
MYOD and
MYOG [
40].
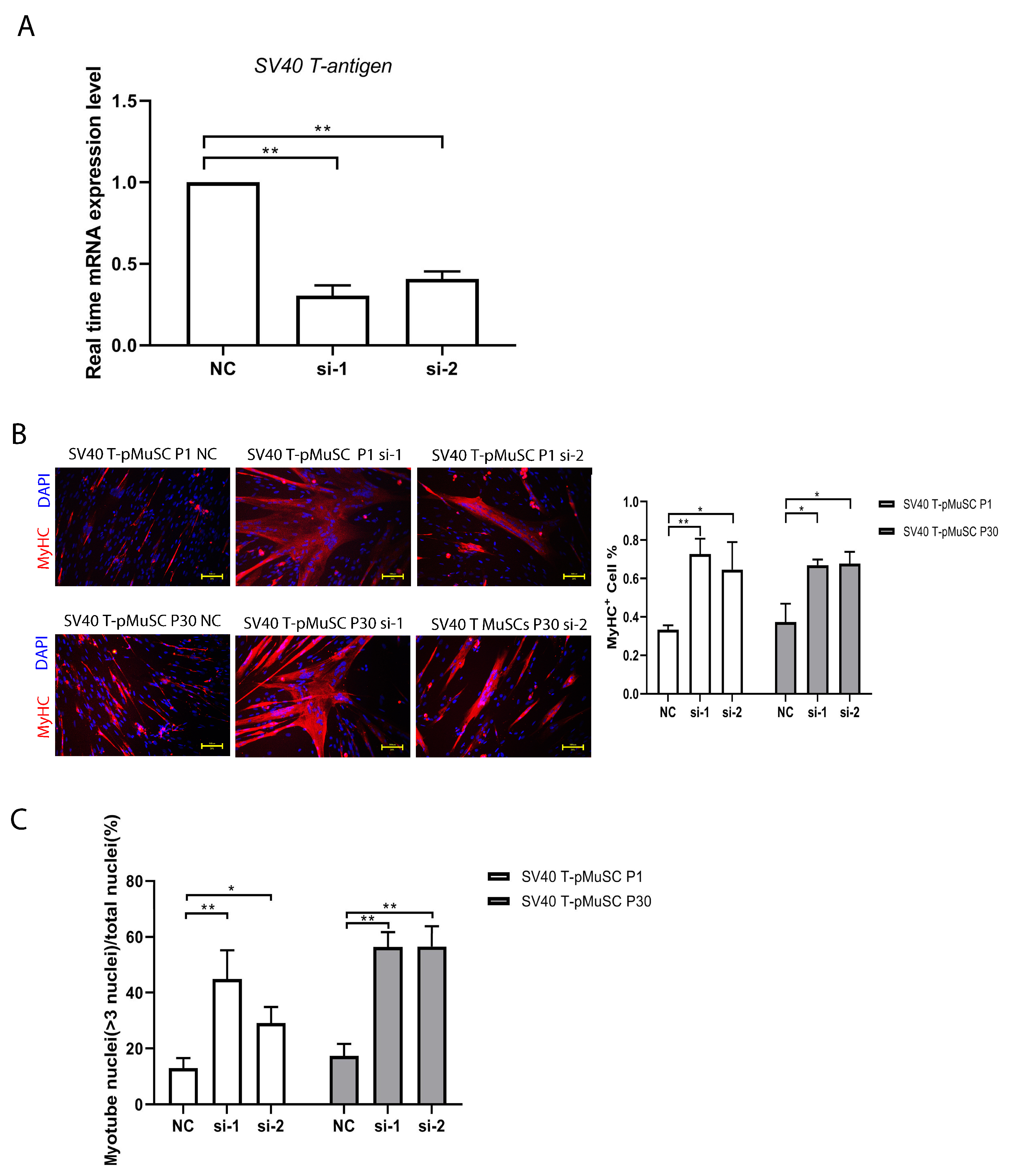
To investigate the potential effects of SV40 T-antigen on the differentiation capacity of pMuSCs, we interfered with the expression of
SV40 T-antigen and assessed the differentiation capacity of SV40 T-pMuSCs. Compared to the negative control (NC), the percentage of MyHC+ cells and fusion index were significantly increased after knockdown of SV40 T-antigen gene, although the fusion index remained slightly lower than that of pMuSCs. Activated satellite cells require cell cycle withdrawal and undergo asymmetric division, with one daughter cell undergoing differentiation [
41]. Due to the continuous promotion of satellite cell proliferation by the SV40 T-antigen, it becomes difficult for cells to exit the cell cycle, thereby affecting their differentiation potential. Telomerase and CDK4 are two biologically significant molecules closely associated with cellular immortalization. Telomerase is an enzyme that maintains the length of telomeres during cell division. CDK4 (cyclin-dependent kinase 4) is a cell-cycle protein kinase that drives the progression of the cell cycle, allowing cells to bypass senescence and apoptosis, thereby achieving immortality [
42]. Andrew et al. developed genetically immortalized bovine satellite cells (iBSCs) via the constitutive expression of bovine telomerase reverse transcriptase (TERT) and cyclin-dependent kinase 4 (CDK4), and maintain their capacity for myogenic differentiation [
43]. This may represent an effective method for immortalizing pMuSCs without damaging their capacity for differentiation.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}



