1. Introduction
Machado-Joseph disease/spinocerebellar ataxia type 3 (MJD/SCA3) (OMIM#109150) is the most frequent autosomal dominant inherited ataxia worldwide [
1]. MJD is caused by an expansion of a polyglutamine (polyQ)-encoding CAG repeat in the
ATXN3 gene [
2]. Ataxin-3 (ATXN3), the protein encoded by
ATXN3, is ubiquitously expressed in different cell types of peripheral and central nervous system tissues [
3]. Neuropathological complexity is a characteristic of this disease, which especially affects the cerebellum, brainstem, basal ganglia, some cranial nerves, and spinal cord [
4]. In average, the age at onset of MJD has been reported around 40 years old, and the mean survival time is estimated to be 21 years [
5]. In MJD, overt disease is preceded by a preclinical phase in which neuropathological and molecular alterations are already present [
6,
7,
8,
9].
Several studies using cellular and animal models of MJD, including the YACMJDQ84.2 (Q84) transgenic mice that are frequently used in pre-clinical studies of MJD [
10,
11,
12,
13,
14], have been revealing that mutant ATXN3 harboring an expanded polyQ tract is involved in the impairment of transcription, mitochondrial function, mechanisms regulating oxidative stress, and apoptosis [
3,
15,
16]. Several of these changes, including transcriptional dysregulation, have also been observed in blood samples from MJD carriers [
17,
18].
Mitochondrial apoptosis is a complex mechanism regulated by several proteins including those belonging to the BCL2 family, such as the anti-apoptotic protein BCL2 (B-cell lymphoma-2) and the pro-apoptotic protein BAX (BCL2 Associated X) [
19]. Another crucial player in apoptosis is the transcription factor P53 (Tumor protein p53) which, among other roles, regulates the levels and the activity of BCL2 family proteins [
20]. In fact, evidence of a direct link between mutant ATXN3 and apoptosis has been provided by Liu and colleagues [
21] who reported that p53 is a substrate of ATXN3 and that mutant ATXN3, exhibiting a stronger deubiquitinase activity than native ATXN3, increases p53 stability and its consequent cellular accumulation leading to increased p53-dependent apoptosis in a MJD zebrafish model and in HCT116 cells. Moreover, our group has formerly shown that peripheral blood cells of MJD carriers display decreased
BCL2/
BAX transcript ratio [
8] resulting from a reduction of blood
BCL2 transcript levels [
18], indicating a higher vulnerability to an apoptotic stimulus in MJD subjects compared with controls. Similarly, at the protein level, abundance of Bcl2 and Bcl2/Bax ratio has been found to be decreased in human neuronal SK-N-SH cells expressing expanded-polyQ ATXN3 compared with the parental cells [
22]. Although some studies using animal and cellular models have shown the involvement of neuronal apoptosis in MJD [
22,
23], little is known about the expression patterns of apoptotic-related genes in the brain and other tissues of individuals with MJD [
24].
Hence, the levels of the apoptosis related BCL2, BAX and TP53 transcripts in cross-sectional and longitudinal samples of peripheral blood from MJD carriers (patients and pre-clinical subjects) were assessed to gather further insights into the dysregulation of mitochondrial apoptosis in MJD and to evaluate their capacity of being used as transcriptional biomarkers of MJD. Additionally, the expression patterns of BCL2, BAX and TP53 genes were investigated in samples from post-mortem brains of MJD patients to better elucidate the involvement of the mitochondrial apoptosis pathway in MJD pathogenesis and to evaluate possible conserved changes of apoptosis markers in the brain and peripheral blood of MJD patients. Finally, the transcript and protein levels of mouse Bcl2, Bax and Trp53 genes were evaluated in samples from the peripheral blood and the brain of Q84 transgenic mice, expressing the full-length human disease ATXN3 gene, to determine whether this widely used MJD mouse model replicates the findings observed in MJD subjects.
2. Materials and Methods
2.1. Human Samples and Clinical Data
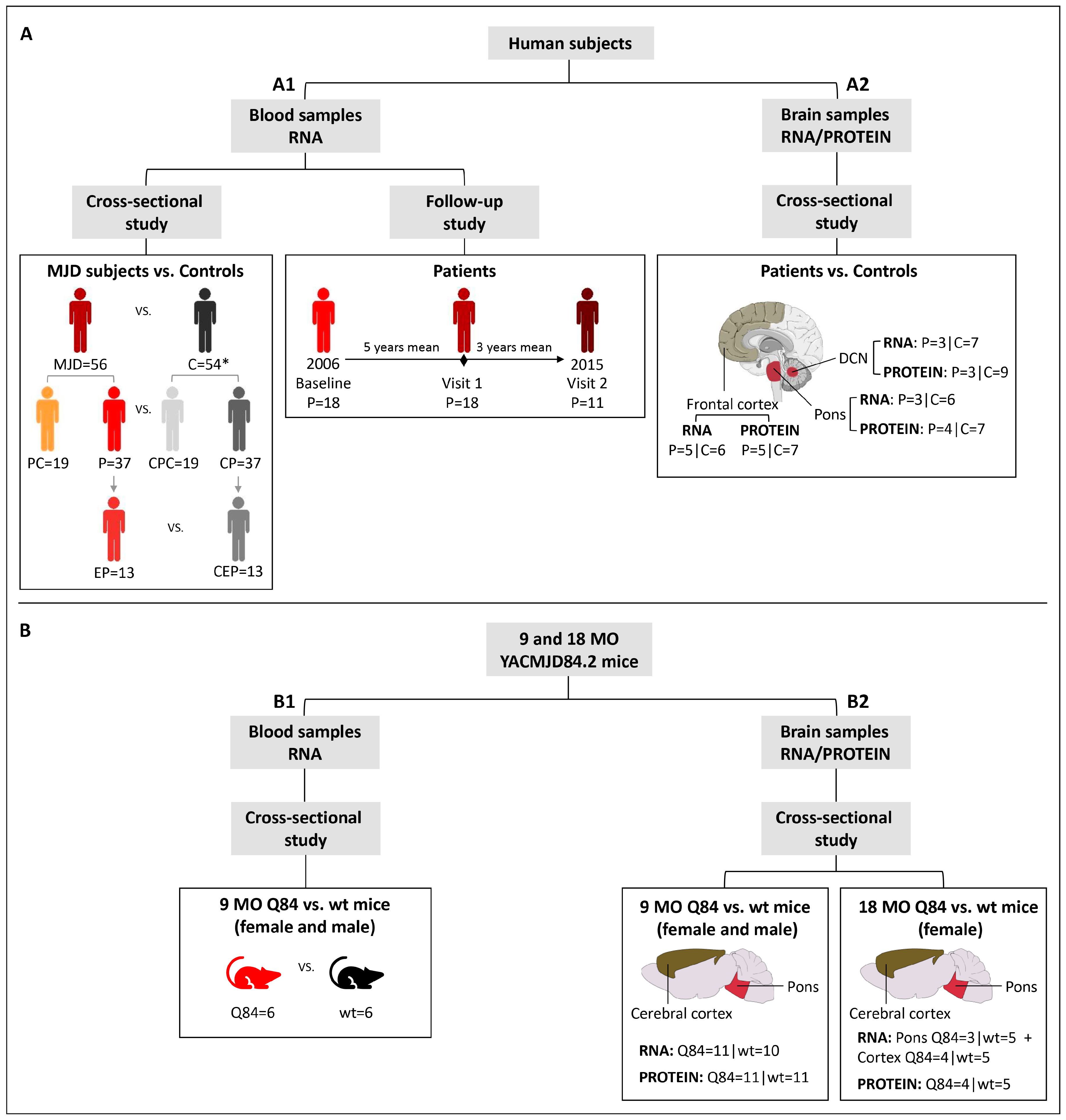
A total of 19 preclinical (no ataxia or diplopia) subjects, 42 MJD patients and 63 healthy controls were included in this study. A summarized clinical, genetic, and demographic characterization of the individuals included in this study is provided in
Supplementary Table S1.
Blood samples: baseline samples from 19 preclinical subjects, 37 patients and 54 age (±3 years) and sex-matched paired controls were used in a cross-sectional study to evaluate the expression levels of
BCL2,
BAX and
TP53 transcripts (
Figure 1(A1);
Supplementary Table S1). A subset of 13 patients with five or less years of disease duration was used to represent the early stage of the disease (early patients) (
Figure 1(A1)). The cut-off used to define the early patients was based on the natural history data for MJD that shows that the mean score of disease progression (defined by gait impairment) evolves very slowly during the first five years [
25]. Additionally, for a subset of 18 MJD patients, blood samples collected at one or two additional observational moments were used in a follow-up study (
Figure 1(A1);
Supplementary Table S2). All MJD subjects underwent a standard neurological evaluation by the same single neurologist. Age at onset was defined as the age of appearance of the first symptoms (gait ataxia and/or diplopia) reported by the patient or a close relative. Disease duration was defined as the time elapsed between age at onset and age at neurological evaluation/blood collection. Preclinical subjects were enrolled in the study after being confirmed as carriers of the
ATXN3 mutation, in the context of the Azorean Genetic Counseling and Predictive Test Program offered by the regional health system to patients and families. For preclinical subjects, the number of years between the age at neurological evaluation/blood collection and the predicted age at disease onset (years to onset, time) was calculated as previously described by Raposo and colleagues [
26]. The determination of the number of CAG repeats at the
ATXN3 locus was performed for all MJD subjects as described by Bettencourt and colleagues [
27]. In addition to the lack of family history of MJD, most of the controls were molecularly excluded for the
ATXN3 mutation using the above-mentioned protocol [
27]. The use of blood samples and clinical data of MJD carriers for the purposes of this study was approved by the Ethics Committee of the University of the Azores (Parecer 5/2017 and Parecer 26/2018). All subjects provided written informed consent for research.
Brain samples: frozen samples from post-mortem brain regions of deidentified and molecularly confirmed MJD patients (n = 5) and molecularly excluded control subjects (n = 9) were obtained from the University of Michigan Brain Bank (
Supplementary Table S1; Supplementary Table S3) and were used in a cross-sectional study (
Figure 1(A2)). Samples from two brain regions severely affected in MJD (dentate cerebellar nucleus (DCN) and pons) [
4] and from a region with a lower degree of disease affectation (frontal cortex) [
28,
29], were used in this study. The
ATXN3 CAG repeat number of all post-mortem brain samples was determined at Laragen Inc. (Culver City, CA, USA) by DNA fragment analysis (
Supplementary Table S3). The use of post-mortem brain samples for the purposes of this study was approved by the University of Michigan Biosafety Committee (IBCA00000265) and by the Ethics Committees of the University of the Azores (Parecer 26/2018).
2.2. Mouse Samples
Hemizygous YACMJD84.2 (Q84) and wild-type (wt) littermate mice [
30] were generated and maintained at the University of Michigan in a C57BL/6J background strain [
10,
14]. Mice were housed in cages with a maximum of five animals (range 3–5) and maintained in a standard 12 h light/dark cycle with food and water ad libitum. Hemizygous Q84 mice show a reduced locomotor and exploratory activity on an open-field test at least at 75 weeks of age (≈17 months-old-MO) [
10]. At least at eight weeks of age, hemizygous Q84 transgenic mice show some neuropathological features of MJD compared to non-transgenic littermates, such as progressive intranuclear accumulation of ATXN3 in neurons of several brain regions that are known to be also affected in MJD patients [
10]. Pre-symptomatic 9 months-old (MO) Q84 (n = 11) and wt mice (n = 11), and symptomatic 18 MO Q84 (n = 4) and wt mice (n = 5) were used in the present study (
Figure 1B;
Supplementary Table S4). Mouse genotyping for the presence of the human disease
ATXN3 transgene was performed using DNA isolated from tail biopsy at the time of weaning, as previously described by Costa and colleagues [
10] and confirmed using DNA extracted from the tail collected post-mortem. The mouse
ATXN3 CAG repeat size was determined as above mentioned for human brain samples. Animal procedures were approved by the University of Michigan Committee on Use and Care of Animals (Protocol PRO00006371). The use of animal samples for the purposes of this study was approved by the Ethics Committees of the University of the Azores (Parecer 26/2018).
Blood samples: mice were anesthetized with ketamine/xylazine, blood was collected from a subset of the 9 MO mice (Q84 = 6 and wt = 6) (
Figure 1(B1)) with a sterile syringe by cardiac puncture, and RNAlater Stabilization Solution (Thermo Fisher Scientific, Waltham, MA, USA) was added to the blood and stored at −20 °C. Mice were subsequently perfused transcardially with phosphate-buffered saline.
Brain samples: 9 MO (Q84 = 11 and wt = 11) and 18 MO (Q84 = 4 and wt = 5) mouse brains (
Figure 1(B2)) were collected after saline perfusion, dissected in the two hemispheres, and the pons and cerebral cortex (isocortex) regions were macro dissected and stored at −80 °C. Brain tissues from the right hemisphere were used for RNA studies and tissues from the left hemisphere were used for protein studies.
2.3. RNA Extraction and cDNA Synthesis
Human samples: total RNA was extracted from blood samples according to the procedures described in Raposo and colleagues [
17]. cDNA was generated from 0.5 µg of total RNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA). For post-mortem brain samples, total RNA was extracted using Trizol (Invitrogen, Waltham, MA, USA), and purified using the RNeasy mini kit (Qiagen, Hilden, Germany) as previously described [
10]. RNA concentration and integrity were assessed using the Agilent 2100 Bioanalyzer (
Supplementary Table S3). cDNA was generated from 1.25 µg of total RNA using the iScript cDNA synthesis kit (BIORAD, Hercules, CA, USA).
Mouse samples: total RNA from mouse blood samples was extracted using the RiboPure RNA Purification Kit (Invitrogen). Total RNA from brain regions was extracted, quantified, and evaluated for integrity as described above for post-mortem human brain samples.
2.4. Quantitative Real-Time PCR
Human samples: qPCR experiments were conducted using the SensiFAST Probe Hi-ROX Master Mix (Bioline, Saint Petersburg, Russia) and the TaqMan Gene Expression Assays (Applied Biosystems) for
BCL2 (Hs00608023_m1),
BAX (Hs00180269_m1) and
TP53 (Hs01034249_m1) using an ABI StepOnePlus Real-Time PCR System (Applied Biosystems). Each sample was run in triplicate (blood) or quadruplicate (brain) alongside with the respective reference gene in the same plate. Experiments including the subset of patients used in the follow-up study were performed by including the samples from a given subject corresponding to all available collection moments in the same plate. In qPCR experiments using RNA from blood samples and post-mortem brain samples,
TRAP1 (TNF receptor associated protein 1; Hs00972326_m1) [
31] and
GAPDH (glyceraldehyde 3-phosphate dehydrogenase; Hs02758991_g1) were, respectively, used as reference genes. The relative expression values were calculated using the 2
−ΔCt method [
32] in DataAssist v3.0 software (Applied Biosystems).
Mouse samples: the qPCR experiments using mouse blood and brain samples were conducted as above described for human blood samples, using the following TaqMan Gene Expression Assays (Applied Biosystems) for Bcl2 (B cell leukemia/lymphoma 2; Mm00477631_m1), Bax (BCL2-associated X protein; Mm00432051_m1) and Trp53 (transformation related protein 53; Mm01731287_m1). For both blood and brain samples, Hmbs (hydroxymethylbilane synthase; Mm01143545_m1) was used as the reference gene.
2.5. Western Blotting
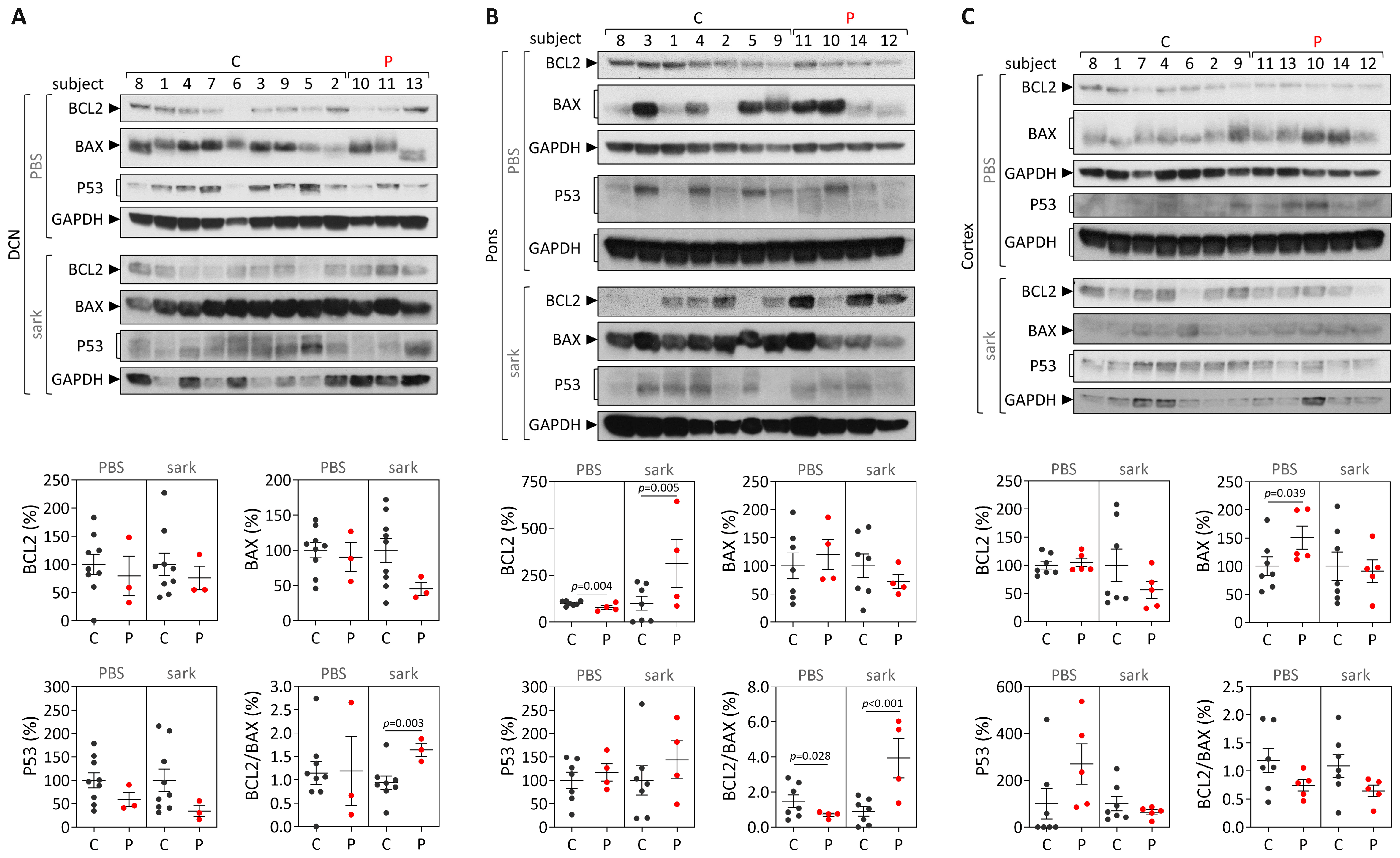
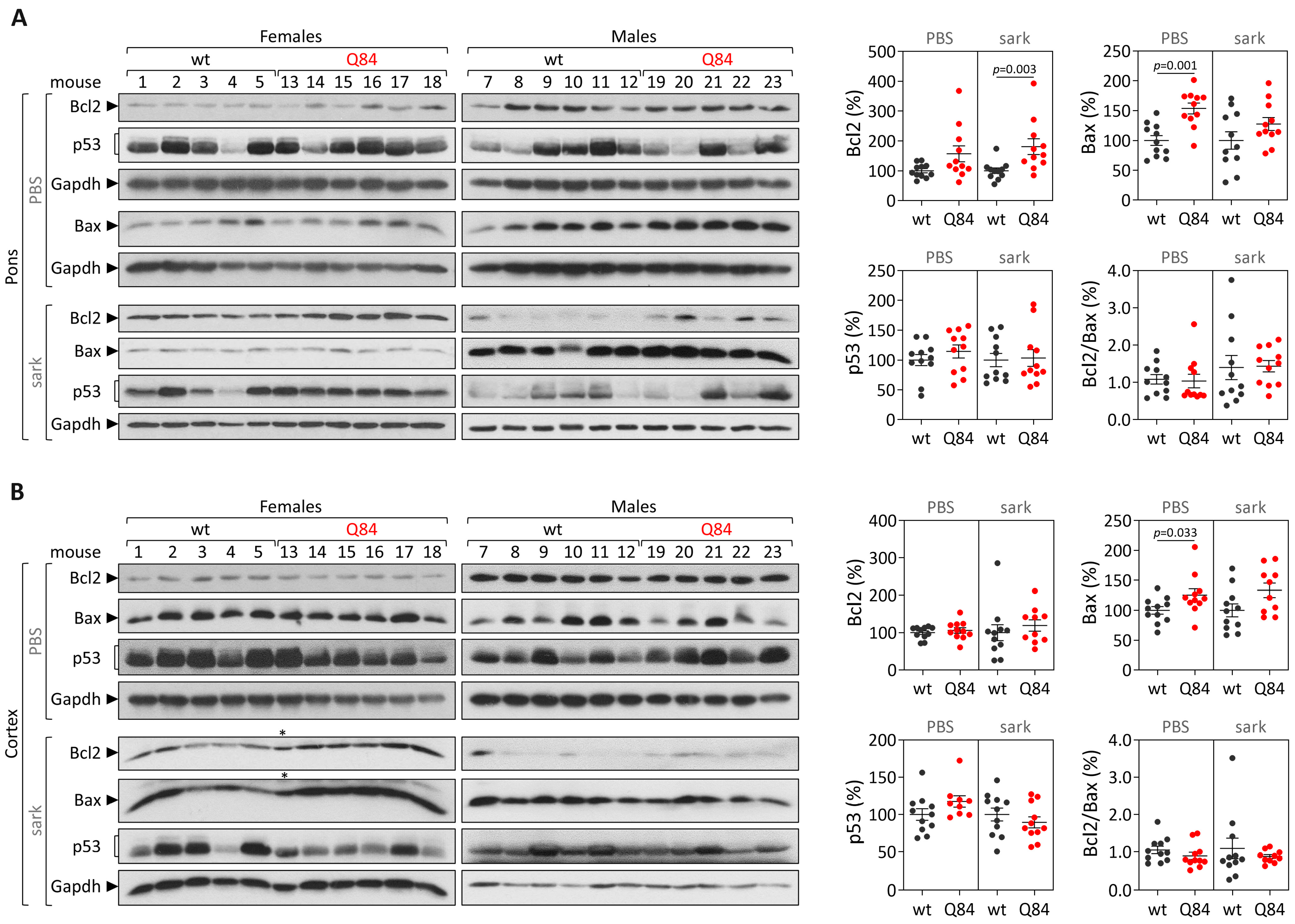
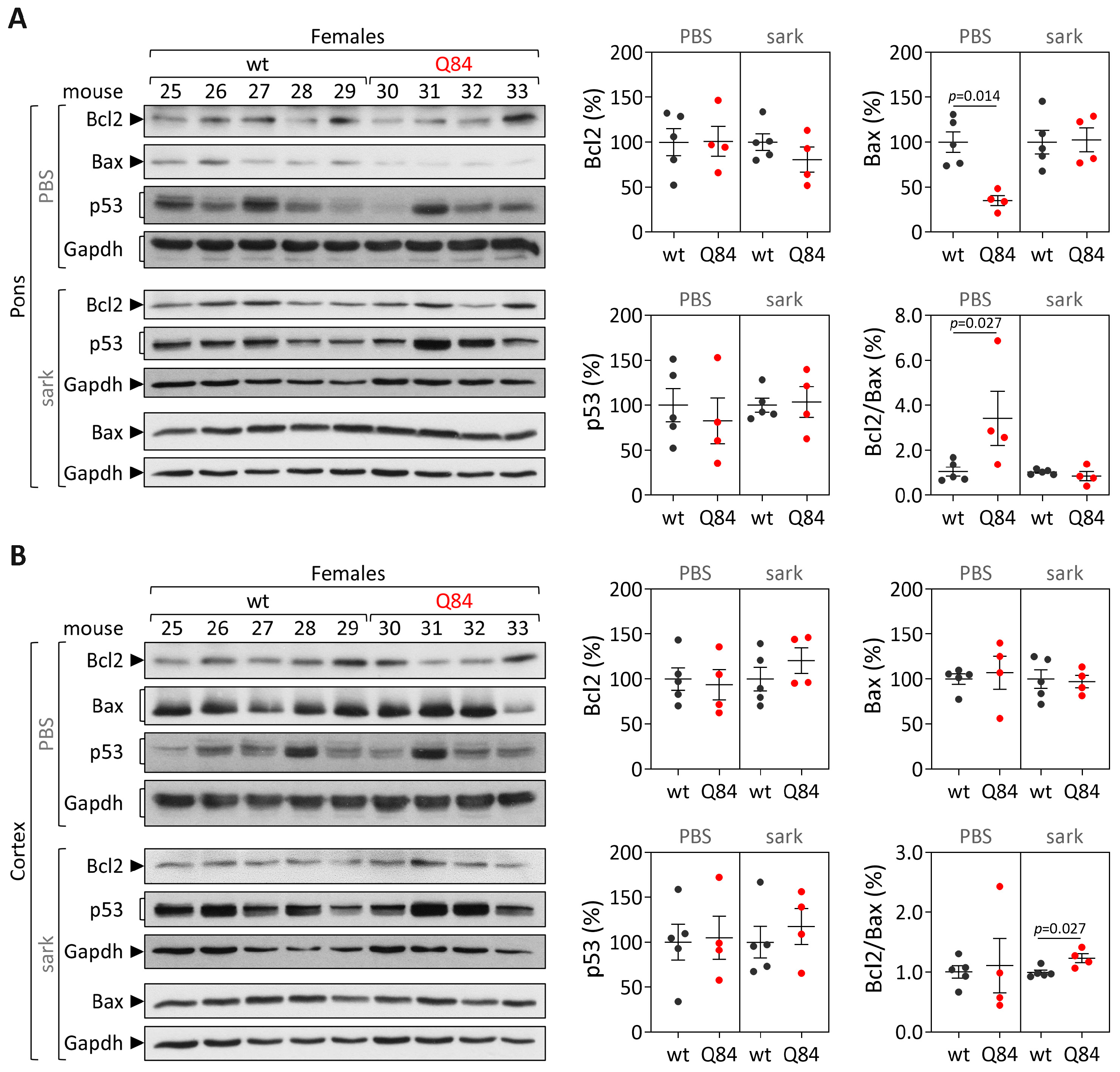
Human samples: protein extracts from DCN, the pons and frontal cortex tissues were obtained by homogenization in cold phosphate-buffered saline (PBS) containing protease inhibitor cocktail (cOmplete Protease Inhibitor Cocktail, Roche) and phosphatase inhibitors (PhosSTOP, Roche, Basel, Switzerland), followed by sonication and centrifugation. The supernatant was collected (PBS-soluble fraction) and stored at −80 °C. The pellet was resuspended in 1% sodium lauroyl sarcosinate (sarkosyl, Sark)/PBS, sonicated, centrifuged and the supernatant (sarkosyl-soluble fraction) was stored at −80 °C. Total protein concentrations from PBS-soluble (soluble) and sarkosyl-soluble (insoluble) fractions were assessed using the BCA method (Pierce BCA Protein Assay Kit; Thermo Fisher Scientific). Total protein lysates from PBS-soluble fractions and sarkosyl-soluble fractions (50 µg) were resolved on 12% SDS-PAGE gels, and corresponding polyvinylidene difluoride (PVDF) membranes were incubated overnight at 4 °C with various antibodies: rabbit anti-Bcl-2 (1:500; #3498, Cell Signaling, Danvers, MA, USA), rabbit anti-Bax (1:500; #2772, Cell Signaling) mouse anti-p53 (1:1000; #2524, Cell Signaling), mouse anti-GAPDH (1:10,000; MAB374, Millipore, Burlington, MA, USA), and mouse anti-ATXN3 (1H9) (1:2000; MAB5360, Millipore). Bound primary antibodies were visualized by incubation with peroxidase affiniPure goat anti-rabbit or anti-mouse secondary antibody (1:10,000; Jackson Immuno Research Laboratories, West Grove, PA, USA), followed by reaction with ECL-plus reagent (Western Lighting, PerkinElmer, Waltham, MA, USA) and subsequent exposure to autoradiography films. Film band intensity was quantified by densitometry on ImageJ.
Mouse samples: protein extraction and quantification from the pons and cerebral cortex from 9 and 18 MO Q84 and wt mice was conducted as previously described for post-mortem human brain samples. Total protein lysates from PBS-soluble and sarkosyl-soluble fractions (40–75 µg) of each brain region were resolved on 12% SDS-PAGE gels.
2.6. Statistical Analysis
2.6.1. Cross-Sectional Studies
Human samples: blood transcript levels of MJD subjects were compared to those of controls using a T-test. When considering the groups of preclinical subjects and patients individually, blood transcript levels were compared to age- and sex-matched paired controls using a Wilcoxon Signed-rank test (two-tailed). The sex of preclinical subjects and patients was compared using a Chi-square test, and age at blood collection and the expanded CAG repeat size were compared using a Mann–Whitney U test (two-tailed). The ability for blood transcript levels to discriminate MJD subjects from healthy controls was assessed using the receiver operating characteristics (ROC) curve analysis; ROC curve accuracy was measured by the area under de curve (AUC) (95% confidence interval (CI), p-value). Correlations between blood transcript levels and clinical features of MJD carriers were conducted using the Spearman’s rank correlation (two-tailed); a partial correlation (two-tailed) was used to adjust for covariates, whenever necessary. Post-mortem brain transcripts and protein levels were compared between biological groups (patients and controls) using a generalized linear model adjusting to the age at death.
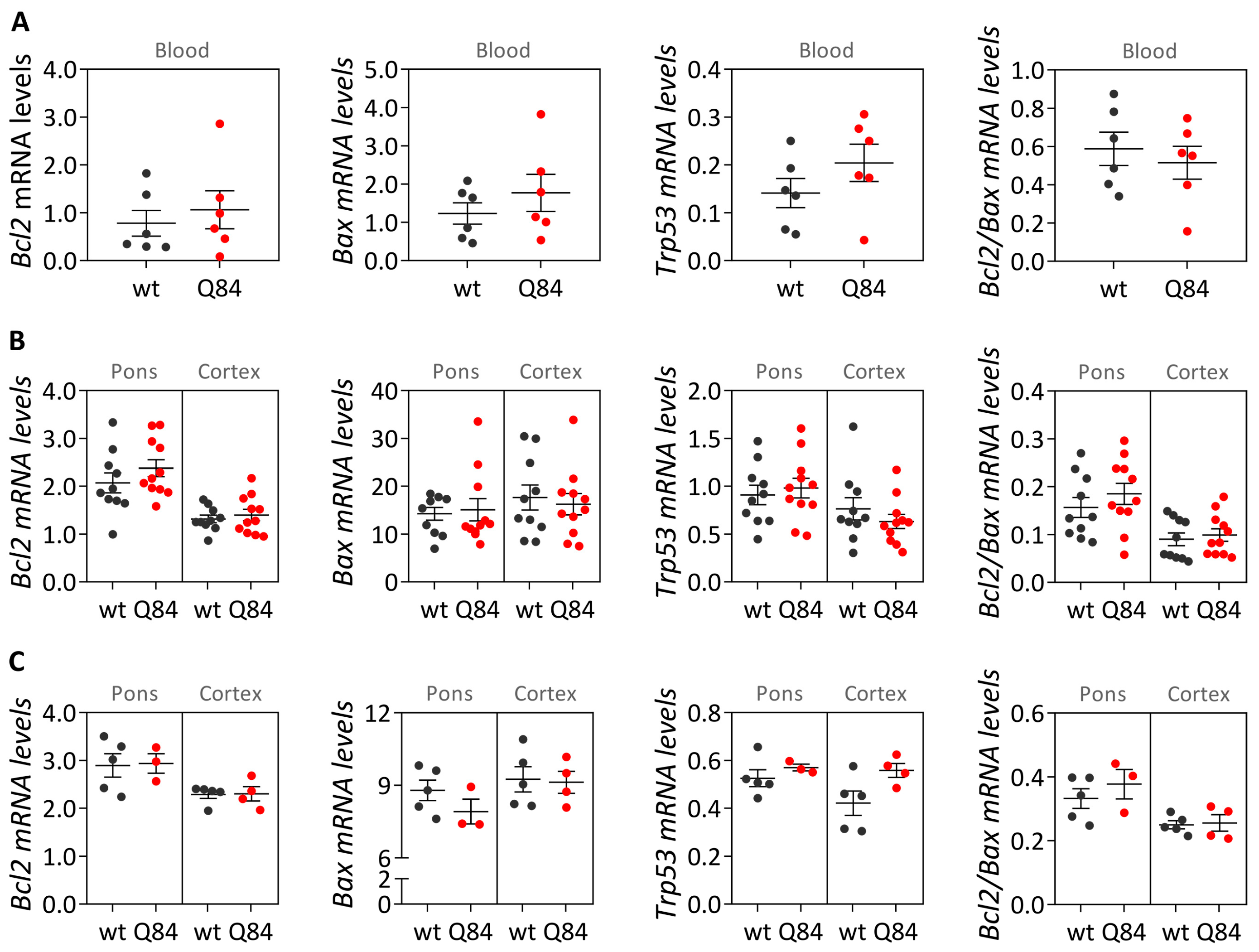
Mouse samples: blood and brain transcript and protein levels of Q84 and wt mice (9 MO males and females combined) were compared using a Mann–Whitney U test (two-tailed).
GraphPad Prism version 8.0.1 for Windows (GraphPad Software, San Diego, CA, USA) was used to identify extreme outliers (ROUT method Q = 1%), which were excluded from data analysis, and to generate the data cross-sectional graphs. Statistical analysis was performed using IBM SPSS Statistics for Windows, version 25 (IBM Corp., Armonk, NY, USA). Statistical significance was set at p < 0.05.
2.6.2. Follow-Up Study of Human Blood Samples
A total of 18 MJD patients were included in the follow-up study. Since repeated observations were performed on the same subjects over a period of 9 years (2006 to 2015), a linear mixed model was considered. The time metric used in the level 1 model was the baseline year 2006. The information about age at blood collection, sex, expanded CAG repeat length, age at onset and disease duration were used in the level 2 model to determine the relationship between blood transcript levels of
BCL2,
BAX and
TP53 as well as the
BCL2/
BAX ratio and the above-mentioned variables. The statistical analysis was performed using R software building on the longitudinal data analysis described by Garcia and colleagues [
33]. Statistical significance was set at
p < 0.05.
4. Discussion
In this study, the expression behavior of the apoptosis-related genes
BCL2,
BAX and
TP53 and the
BCL2/
BAX ratio, an indicator of susceptibility to apoptosis [
34], was evaluated in peripheral blood and post-mortem brain samples from MJD subjects and Q84 transgenic mice. Like previous reports [
8,
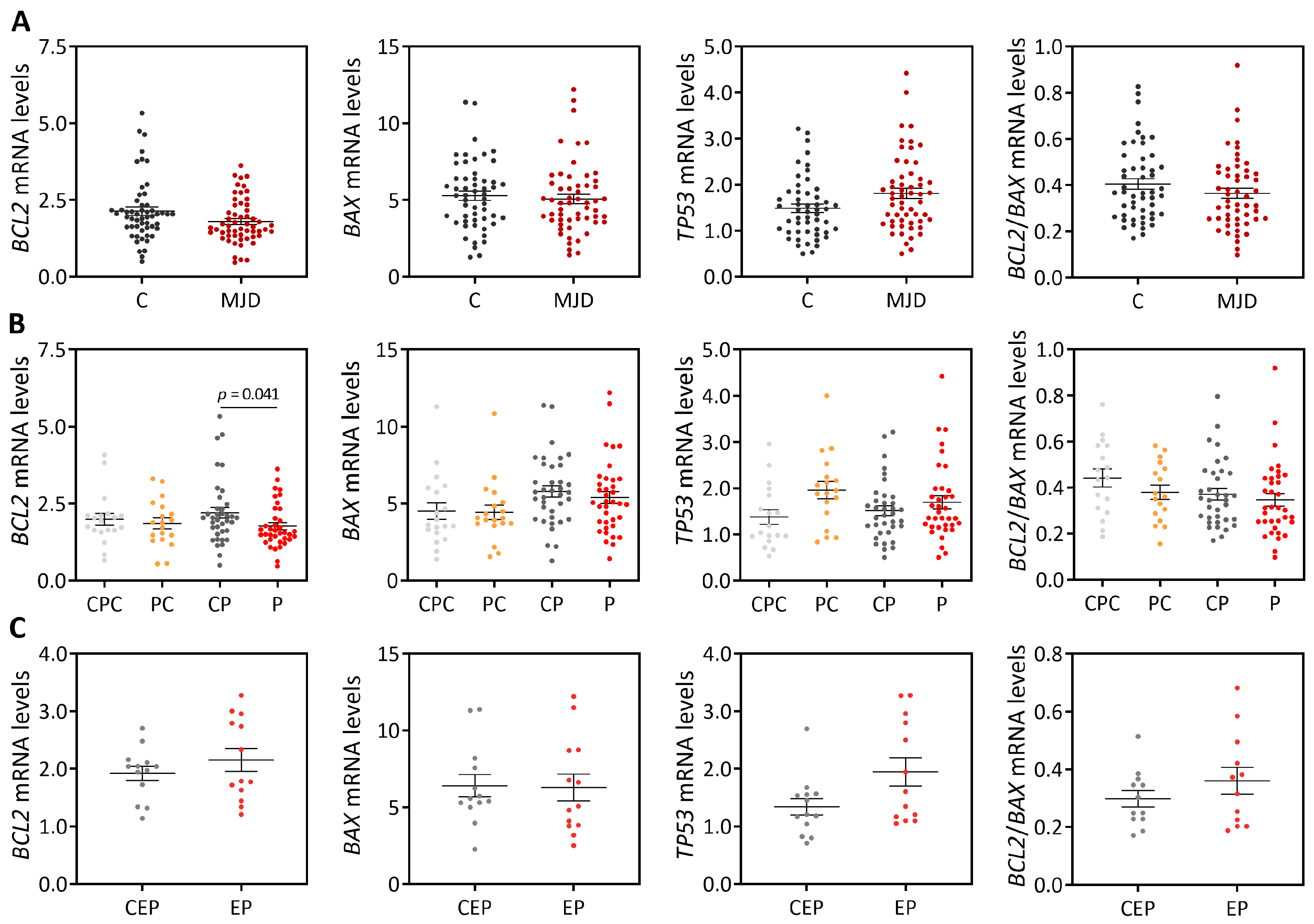
18], our cross-sectional analysis showed evidence of reduced
BCL2 transcript levels in blood of MJD patients. However, the abundance of blood
BCL2 transcripts revealed a low capacity to discriminate these two biological groups, thus precluding the use of
BCL2 levels as a biomarker of MJD. Similar blood
BCL2/
BAX transcript ratio was further found in MJD patients and matched controls, which contrasts with the previous report describing a reduced
BCL2/
BAX transcript ratio in MJD patients [
8]. The partial replication of the published results [
8] could perhaps be explained by the analysis of a small number of patients (n = 37) and the use of age- and sex-matched paired controls in this study comparing with the study by Raposo and colleagues analyzing a higher number of patients (n = 74) but non-matched controls [
8]. In addition, similar blood abundance of
BCL2 transcripts and the
BCL2/
BAX transcript ratio was found here in MJD preclinical subjects and matched controls (n = 19), also contrasting with previous reports showing reduced
BCL2 levels [
18] and the
BCL2/
BAX transcript ratio in the blood of preclinical subjects (n = 16) [
8]. Again, the different observations in MJD preclinical subjects described here and in the previous reports [
8,
18] could be due to the use of different experimental designs, namely the use of age- and sex-matched paired controls in this study versus non-matched controls used in the published reports [
8,
18]. Noteworthy, in this study, increased levels of blood pro-apoptotic
BAX transcripts and a decreased
BCL2/
BAX ratio were found to be associated with earlier age at onset, indicating that these transcriptional alterations may be associated with MJD pathogenesis. Consistently with these findings, and although global
TP53 levels are similar in patients and healthy controls, our follow-up analysis of blood samples from 18 MJD patients collected at distinct moments of disease progression over a maximum period of nine years showed that the abundance of pro-apoptotic
TP53 transcripts increases with disease progression. In contrast, and while in a lower extent than
TP53 transcript changes, levels of anti-apoptotic
BCL2 transcripts also increased with disease progression. The increased abundance of blood
TP53 and
BCL2 transcripts with MJD progression may indicate that pro-apoptotic and anti-apoptotic signs increase concomitantly in this tissue with the disease course.
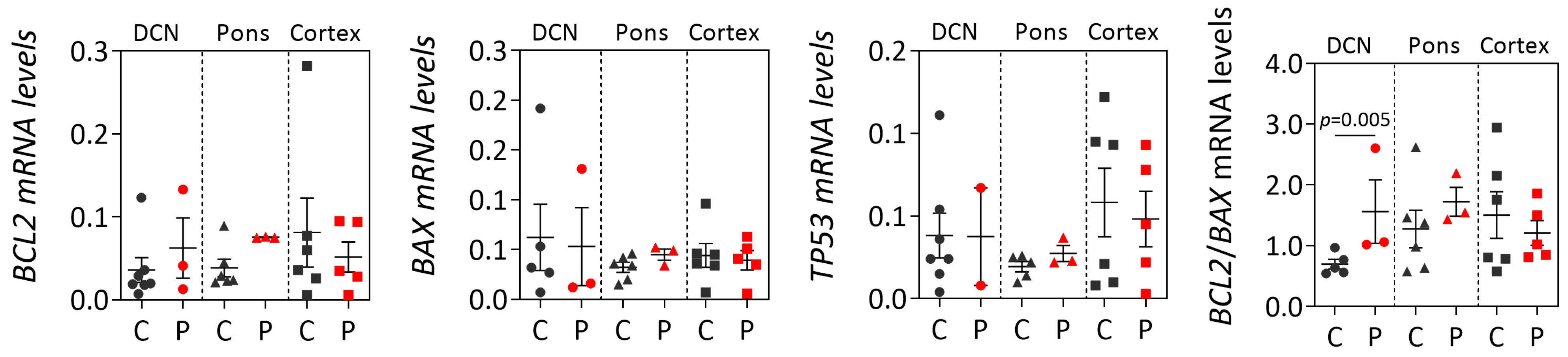
While post-mortem brains from MJD patients are very rare, dispersed throughout several brain banks in the world and studies using such materials usually show a low power, the analysis of human MJD brain samples contributes to the elucidation of altered cellular mechanisms at the end stage of disease that may be associated with MJD pathogenesis. Overall, our cross-sectional findings of the increased
BCL2/
BAX transcript ratio in DCN and the increased insoluble protein BCL2/BAX ratio in DCN and the pons of post-mortem MJD brains suggest that, contrarily to what would be expected, cells in the DCN and pons, which are brain areas severely affected by degeneration in MJD [
4,
28,
29] might be more prone to survive compared with controls. Interestingly, this behavior was not observed in the less-affected frontal cortex of MJD patients that only showed significantly increased soluble BAX levels, suggesting that this brain region displays similar susceptibility to apoptosis in MJD patients and in control individuals. The evidence of increased abundance of insoluble BCL2/BAX ratio observed in the DCN and pons of MJD patients may indicate increased abundance of the BCL2 protein in the mitochondrial outer membrane thus preventing apoptosis activation by inhibiting the activity of pro-apoptotic BCL2 family members [
35]. However, the possibility that some of these insoluble proteins could instead be recruited to ATXN3 aggregates or be localized in other cellular membranes cannot be excluded. Yet, the results observed in the DCN and pons of MJD patients agree with previous reports of absence of TUNEL-positive cells and changes of expression of apoptosis-related proteins (BCL2, P53, BAX or CPP32) in post-mortem DCN from MJD patients [
24]. MJD patients globally show a long survival time (time elapsed from onset to death) [
36] and thus post-mortem MJD brain samples most likely represent end stages of the disease. Hence, the most affected neurons and other cells in the DCN and pons of MJD patients [
4] are most probably dead and absent at the end stage of disease and, therefore, the increased anti-apoptotic signs (BCL2/BAX ratio) observed in both brain regions could indicate that the surviving glial and neuronal cells in these two affected regions activate survival protection mechanisms at the end-stage of MJD. Similarly, Satou et al. [
37] reported that the abundance of BCL2 protein within neurons of post-mortem brains from Alzheimer’s disease patients increased with disease severity, suggesting that BCL2 protein may have a protective role at the end-stage of the disease.
In parallel with the study in human subjects, mouse
Bcl2,
Bax and
Trp53 transcript and protein levels were assessed in blood of pre-symptomatic 9 MO Q84 transgenic mice, and in the pons and cerebral cortex of both pre-symptomatic 9 MO and symptomatic 18 MO Q84 mice [
10,
30] to evaluate whether this widely used MJD mouse model replicates the findings observed in MJD subjects. Our cross-sectional analysis of Q84 mice, showed similar
Bcl2,
Bax and
Trp53 transcript levels in the blood of pre-symptomatic 9 MO Q84 mice and controls. These results in the pre-symptomatic Q84 mice agree with the observed lack of differences between the abundance of
BCL2,
BAX and
TP53 transcripts in peripheral blood samples from preclinical MJD subjects and age- and sex-matched paired controls. Additionally, similar transcript levels of
Bcl2,
Bax and
Trp53 were found in the pons and cerebral cortex of pre-symptomatic 9 MO or symptomatic 18 MO Q84 transgenic mice and wt controls. Interestingly, the similar abundance of the three assessed transcripts in the pons and cerebral cortex of symptomatic Q84 mice and controls mimics the observations in post-mortem brains from MJD patients further indicating that the Q84 mice replicate several aspects of the human disease. In contrast to the observed similar abundance of
Bcl2,
Bax and
Trp53 transcripts in pre-symptomatic and symptomatic Q84 and wt mice, altered levels of Bcl2 and/or Bax proteins were found in the two assessed brain regions of pre-symptomatic and symptomatic Q84 transgenic mice compared with wt littermates. Despite the altered abundance of these two proteins in the pons and cerebral cortex of pre-symptomatic 9 MO Q84 mice, the cells present in these brain regions show similar susceptibility to apoptosis as in controls, as shown by the similar insoluble Bcl2/Bax ratio. Likewise, cells in the pons of symptomatic 18 MO Q84 transgenic mice and controls show a similar susceptibility to apoptosis, while the cells in the less-affected cerebral cortex of symptomatic Q84 mice display increased levels of insoluble Bcl2/Bax ratio possibly indicating higher apoptosis resistance. In summary, pre-symptomatic and symptomatic Q84 transgenic mice show altered abundance of Bcl2 and Bax proteins with disease progression in both brain regions. Additionally, the similar results observed in the less-affected cerebral cortex of symptomatic 18 MO Q84 mice and in the severely affected the DCN and pons of MJD patients suggest that symptomatic Q84 mice are at different stages of the disease compared to post-mortem MJD brains. Globally, the mitochondrial apoptosis mechanisms in cells present in the highly affected and less-affected brain regions seem to respond differently in an age-dependent way to the damage elicited by the expanded
ATXN3 CAG repeat in MJD.
Overall, our study displays some limitations such as the limited statistical power of the analysis of post-mortem brain samples due to the small sample size, and the lack of blood transcript levels over disease progression in Q84 transgenic mice due to the lack of peripheral blood samples from symptomatic 18 MO transgenic Q84 mice. The fact that no transcriptional differences were found in the Q84 mouse model suggests that this animal model may replicate the human disease, yet future studies using mouse samples from several points along disease progression would need to be performed to test this hypothesis. Importantly, future evaluation of the abundance of a wider number of apoptotic-related molecules including markers of the late phase of apoptosis in peripheral blood samples from MJD subjects and post-mortem MJD brains could reveal novel biomarkers of MJD and could help elucidate the apoptotic signaling cascades that are majorly involved in MJD. Lastly, because mutant ATXN3 stabilizes P53 leading to an increased abundance of p53 in MJD cellular models [
21], future quantification of the P53 protein in peripheral blood from MJD carriers could reveal a new progression biomarker for MJD.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






