
Comparison of Morphological, Physiological, and Related Gene Expression Responses to Drought Stress in Five Camellia vietnamensis Cultivars
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment
2.2. Morphological Parameters
2.3. Physiological Index
2.3.1. Determination of Membrane Permeability-Relevant Indexes, Osmoregulatory Substance Contents, and Antioxidant Enzyme Activity
2.3.2. Photosynthetic Pigment Content
2.3.3. Total Polyphenol (Pol), Total Flavonoid (Fla) and Tea Saponin (TS) Contents
- (1)
- Preparation of C. vietnamensis leaf extracts
- (2)
- Pol content determination
- (3)
- Fla content determination
- (4)
- TS content determination
2.4. RT-qPCR Analysis
2.5. Comprehensive Evaluation of Drought Resistance
2.5.1. Drought-Tolerance Coefficients
2.5.2. Principal Component Analysis
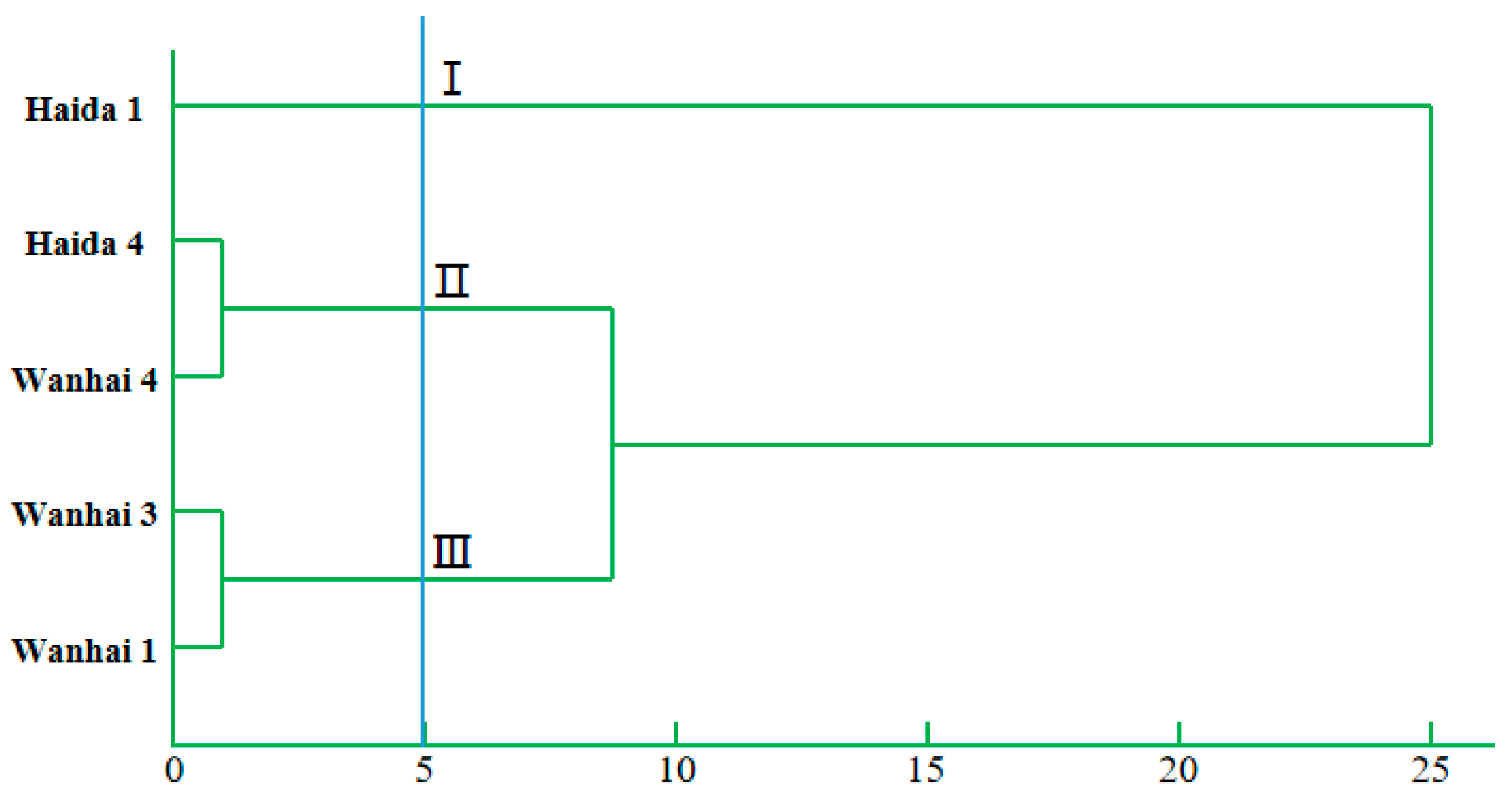
2.6. Cluster Analysis
2.7. Data statistics and Analysis
3. Results
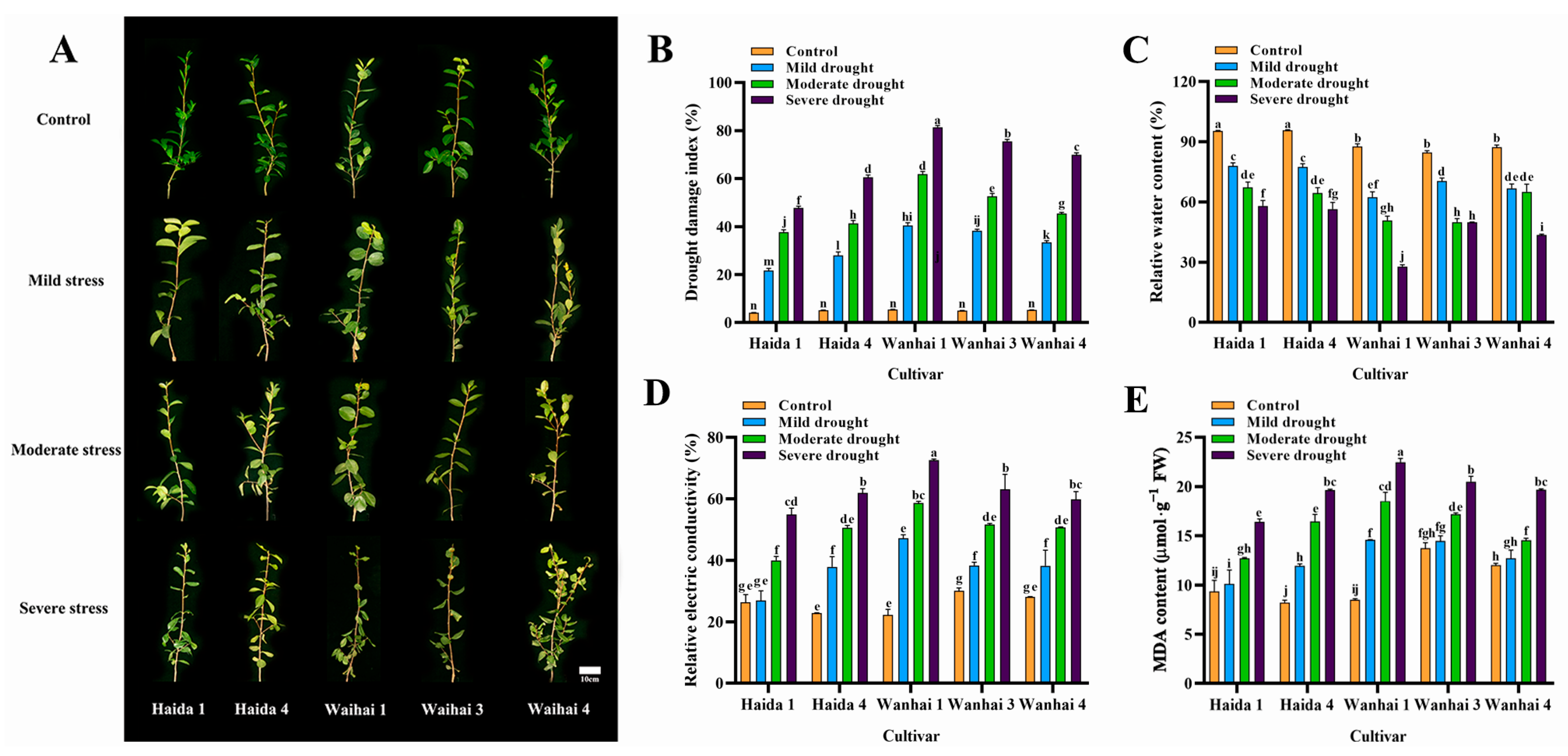
3.1. Effects of Drought Stress on the Morphological Characteristics and Membrane Permeability of Five C. vietnamensis Cultivars
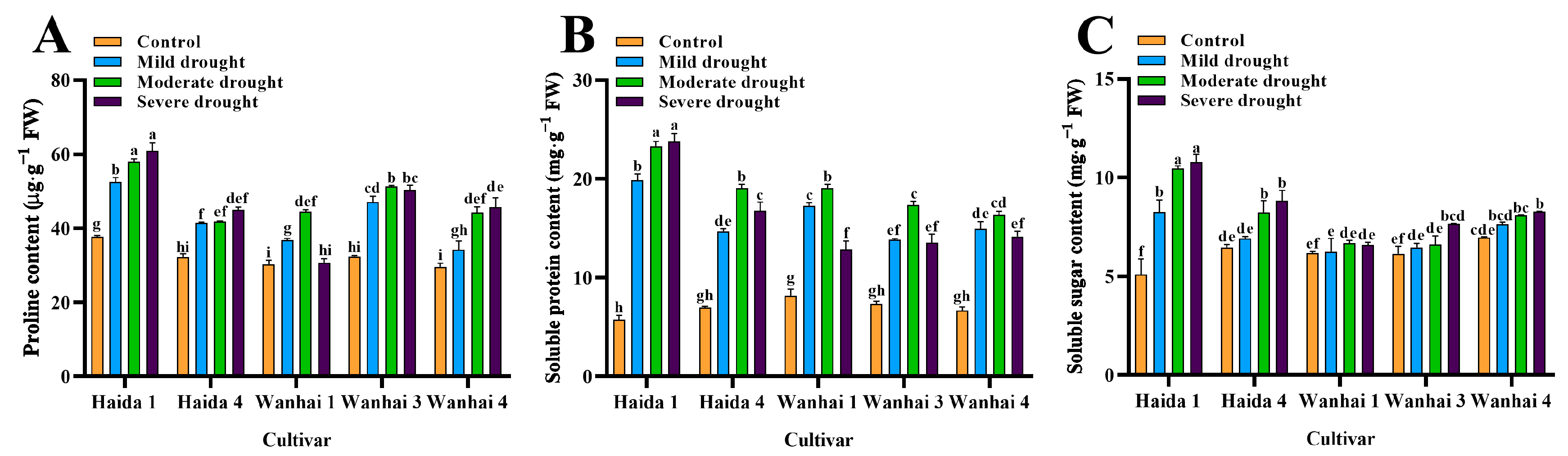
3.2. Effects of Drought Stress on the Osmotic Regulation-Substance Content of Five C. vietnamensis Cultivars
3.3. Effects of Drought Stress on the Photosynthetic Pigment Contents of Five C. vietnamensis Cultivars
3.4. Effects of Drought Stress on Antioxidant Enzyme Activities and Secondary Metabolites of Five C. vietnamensis Cultivars
3.5. Effects of Drought Stress on the Expression of Key Genes in Phenylpropane Metabolism
3.6. Comprehensive Evaluation of Drought Resistance of C. vietnamensis
4. Discussion
4.1. Plant Growth and Membrane Lipid Peroxidation
4.2. Physiological Responses and Drought Tolerance
4.3. Response of Key Genes and Metabolites in the Phenylpropane Pathway to Drought Tolerance
4.4. PCA, HCA and Multivariate Statistical Analysis of Drought Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Zeppel, M.; Anderegg, W.; Hartmann, H.; Mcdowell, N.G. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Prăvălie, R. Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 2016, 161, 259–278. [Google Scholar] [CrossRef]
- Ault, T.R. On the essentials of drought in a changing climate. Science 2020, 368, 256–260. Available online: https://www.science.org/doi/10.1126/science.aaz5492 (accessed on 20 December 2022). [CrossRef] [PubMed]
- Mondal, S.K.; Mishra, A.; Leung, R.; Cook, B. Global droughts connected by linkages between drought hubs. Nat. Commun. 2023, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Rane, J.; Singh, A.K.; Tiwari, M.; Prasad, P.V.; Jagadish, S.V. Effective use of water in crop plants in dryland agriculture: Implications of reactive oxygen species and antioxidative system. Front. Plant Sci. 2022, 12, 778270. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Zörb, C.; Geilfus, C.M. Drought and crop yield. Plant Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Hendrawan, V.S.A.; Komori, D.; Kim, W. Possible factors determining global-scale patterns of crop yield sensitivity to drought. PLoS ONE 2023, 18, e0281287. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, F.; Chen, B.; Su, E.; Chen, Y.; Cao, F. Composition, bioactive substances, extraction technologies and the influences on characteristics of Camellia oleifera oil: A review. Food Res. Int. 2022, 156, 111159. [Google Scholar] [CrossRef]
- Li, M.; Wang, G.; Zong, S.; Chai, X. Copula-Based Assessment and Regionalization of Drought Risk in China. Int. J. Environ. Res. Public Health 2023, 20, 4074. [Google Scholar] [CrossRef]
- Mao, D.; Wang, Z.; Wu, B.; Zeng, Y.; Luo, L.; Zhang, B. Land degradation and restoration in the arid and semiarid zones of China: Quantified evidence and implications from satellites. Land Degrad. Dev. 2018, 29, 3841–3851. [Google Scholar] [CrossRef]
- Gao, S.; Wang, Y.; Yu, S.; Huang, Y.; Liu, H.; Chen, W.; He, X. Effects of drought stress on growth, physiology and secondary metabolites of Two Adonis species in Northeast China. Sci. Hortic. 2020, 259, 108795. [Google Scholar] [CrossRef]
- Lim, C.W.; Baek, W.; Jung, J.; Kim, J.H.; Lee, S.C. Function of ABA in stomatal defense against biotic and drought stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic signaling during abiotic stress combination in plants. Proc. Nati. Acad. Sci. USA 2020, 117, 13810–13820. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B.S.M.A.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Balatova, Z.; Drevenakova, P.; Olsovska, K.; Kalaji, H.M.; Yang, X.; Allakhverdiev, S.I. Photosynthetic electron transport and specific photoprotective responses in wheat leaves under drought stress. Photosynth. Res. 2013, 117, 529–546. [Google Scholar] [CrossRef]
- Waititu, J.K.; Zhang, X.; Chen, T.; Zhang, C.; Zhao, Y.; Wang, H. Transcriptome analysis of tolerant and susceptible maize genotypes reveals novel insights about the molecular mechanisms underlying drought responses in leaves. Int. J. Mol. Sci. 2021, 22, 6980. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Ji, Y.; Sun, M.; Lin, C.; Zhou, P.; Ren, J.; Luo, D.; Wang, X.; Ma, C.; Zhang, X.; et al. Research on the drought tolerance mechanism of Pennisetum glaucum (L.) in the root during the seedling stage. BMC Genom. 2021, 22, 568. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Han, M.; Yang, L.M.; Li, Y.; Sun, Z.; Zhang, T. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crops Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, B.; Hu, T.; Chen, H.; Li, H.; Zhang, W.; Zhong, Y.; Hu, H. Combined action of an antioxidant defence system and osmolytes on drought tolerance and post-drought recovery of Phoebe zhennan S. Lee saplings. Acta Physiol. Plant. 2015, 37, 84. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.S.M.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of salicylic acid, zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress. Sci. Rep. 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Shawon, R.A.; Kang, B.S.; Lee, S.G.; Kim, S.K.; Lee, H.J.; Katrich, E.; Gorinstein, S.; Ku, Y.G. Influence of drought stress on bioactive compounds, antioxidant enzymes and glucosinolate contents of Chinese cabbage (Brassica rapa). Food Chem. 2020, 308, 125657. [Google Scholar] [CrossRef]
- Luo, C.; Min, W.; Akhtar, M.; Lu, X.; Bai, X.; Zhang, Y.; Tian, L.; Li, P. Melatonin enhances drought tolerance in rice seedlings by modulating antioxidant systems, osmoregulation, and corresponding gene expression. Int. J. Mol. Sci. 2022, 23, 12075. [Google Scholar] [CrossRef] [PubMed]
- Shehata, S.A.; Omar, H.S.; Elfaidy, A.G.; El-Sayed, S.S.; Abuarab, M.E.; Abdeldaym, E.A. Grafting enhances drought tolerance by regulating stress-responsive gene expression and antioxidant enzyme activities in cucumbers. BMC Plant Biol. 2022, 22, 408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, M.; Zhao, Z.; Ren, Y.; Li, Q.; Wang, W. Wheat TaPUB1 modulates plant drought stress resistance by improving antioxidant capability. Sci. Rep. 2017, 7, 7549. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Shen, S.; Yan, W.; Xie, S.; Yu, J.; Yao, G.; Xia, P.; Wu, Y.; Yang, H. Physiological and transcriptional analysis reveals the response mechanism of Camellia vietnamensis Huang to drought stress. Int. J. Mol. Sci. 2022, 23, 11801. [Google Scholar] [CrossRef]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Du, Z.; Hu, Y.; Buttar, N.A. Analysis of mechanical properties for tea stem using grey relational analysis coupled with multiple linear regression. Sci. Hortic. 2020, 260, 108886. [Google Scholar] [CrossRef]
- Gedam, P.A.; Thangasamy, A.; Shirsat, D.V.; Ghosh, S.; Bhagat, K.P.; Sogam, O.A.; Gupta, A.J.; Mahajan, V.; Soumia, P.S.; Salunkhe, V.N.; et al. Screening of onion (Allium cepa L.) genotypes for drought tolerance using physiological and yield based indices through multivariate analysis. Front. Plant Sci. 2021, 12, 600371. [Google Scholar] [CrossRef]
- Xiong, S.; Wang, Y.; Chen, Y.; Gao, M.; Zhao, Y.; Wu, L. Effects of drought stress and rehydration on physiological and biochemical properties of four oak species in China. Plants 2022, 11, 679. [Google Scholar] [CrossRef]
- Zhang, T.; Qiu, F.; Chen, L.; Liu, R.; Chang, M.; Wang, X. Identification and in vitro anti-inflammatory activity of different forms of phenolic compounds in Camellia oleifera oil. Food Chem. 2021, 344, 128660. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, H.Y.; Yang, X.N.; Zhan, J.J.; Zhou, H.; Wang, C.; Yu, Y.; Lu, X.Y.; Chen, Y.Z.; Tian, Y. Transcriptomic analysis of Camellia oleifera in response to drought stress using high throughput RNA-seq. Russ. J. Plant Physiol. 2017, 64, 728–737. [Google Scholar] [CrossRef]
- He, Z.; Liu, C.; Zhang, Z.; Wang, R.; Chen, Y. Integration of mRNA and miRNA analysis reveals the differentially regulatory network in two different Camellia oleifera cultivars under drought stress. Front. Plant Sci. 2022, 13, 1001357. [Google Scholar] [CrossRef]
- He, Z.; Cui, K.; Wang, R.; Xu, T.; Zhang, Z.; Wang, X.; Chen, Y.; Zhu, Y. Multi-omics joint analysis reveals how Streptomyces albidoflavus OsiLf-2 assists Camellia oleifera to resist drought stress and improve fruit quality. Front. Microbiol. 2023, 14, 1152632. [Google Scholar] [CrossRef]
- Dong, B. Comprehensive Evaluation of Drought Resistance and Transcriptome Analysis under Drought Stress in Camellia oleifera; South China Agricultural University: Guangzhou, China, 2018. [Google Scholar]
- Sapes, G.; Sala, A. Relative water content consistently predicts drought mortality risk in seedling populations with different morphology, physiology and times to death. Plant Cell Environ. 2021, 44, 3322–3335. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.A.M.E.L.A.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Aghaie, P.; Tafreshi, S.A.H.; Ebrahimi, M.A.; Haerinasab, M. Tolerance evaluation and clustering of fourteen tomato cultivars grown under mild and severe drought conditions. Sci. Hortic. 2018, 232, 1–12. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bouslamti, M.; El Barnossi, A.; Kara, M.; Alotaibi, B.S.; Al Kamaly, O.; Assouguem, A.; Lyoussi, B.; Benjelloun, A.S. Total polyphenols content, antioxidant and antimicrobial activities of leaves of Solanum elaeagnifolium Cav. from Morocco. Molecules 2022, 27, 4322. [Google Scholar] [CrossRef]
- Gorinstein, S.; Vargas, O.J.M.; Jaramillo, N.O.; Salas, I.A.; Ayala, A.L.M.; Arancibia-Avila, P.; Toledo, F.; Katrich, E.; Trakhtenberg, S. The total polyphenols and the antioxidant potentials of some selected cereals and pseudocereals. Eur. Food Res. Technol. 2007, 225, 321–328. [Google Scholar] [CrossRef]
- Wu, X.; Jia, L.; Wu, J.; Liu, Y.; Kang, H.; Liu, X.; Li, P.; He, P.; Tu, Y.; Li, B. Simultaneous determination and quantification of triterpene saponins from Camellia sinensis seeds using UPLC-PDA-QTOF-MS/MS. Molecules 2019, 24, 3794. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Ye, Z.; Yu, J.; Yan, W.; Zhang, J.; Yang, D.; Yao, G.; Liu, Z.; Wu, Y.; Hou, X. Integrative iTRAQ-based proteomic and transcriptomic analysis reveals the accumulation patterns of key metabolites associated with oil quality during seed ripening of Camellia oleifera. Hortic. Res. 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Li, W.; Zhao, L.; Meng, F.; Wang, Y.; Tan, H.; Yang, H.; Wei, C.; Wan, X.; et al. Tissue-specific, development-dependent phenolic compounds accumulation profile and gene expression pattern in tea plant [Camellia sinensis]. PLoS ONE 2013, 8, e62315. [Google Scholar] [CrossRef]
- Zhao, T.; Pan, X.; Ou, Z.; Li, Q. Comprehensive evaluation of waterlogging tolerance of eleven Canna cultivars at flowering stage. Sci. Hortic. 2022, 296, 110890. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.; Cervilla, L.M.; Blasco, B.; Rios, J.J.; Rosales, M.A.; Romero, l.; Ruiz, J.M. Genotypic differences in some physiological parameters symptomatic for oxidative stress under moderate drought in tomato plants. Plant Sci. 2010, 178, 30–40. [Google Scholar] [CrossRef]
- Dong, B.; Wu, B.; Hong, W.; Li, X.; Li, Z.; Xue, L.; Huang, Y. Transcriptome analysis of the tea oil camellia (Camellia oleifera) reveals candidate drought stress genes. PLoS ONE 2017, 12, e0181835. [Google Scholar] [CrossRef] [PubMed]
- Bunnag, S.; Pongthai, P. Selection of rice (Oryza sativa L.) cultivars tolerant to drought stress at the vegetative stage under field conditions. Am. J. Plant Sci. 2013, 4, 1701. [Google Scholar] [CrossRef]
- Han, C.; Chen, J.; Liu, Z.; Chen, H.; Yu, F.; Yu, W. Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress. Agronomy 2022, 12, 1461. [Google Scholar] [CrossRef]
- Li, P.; Yang, H.; Wang, L.; Liu, H.; Huo, H.; Zhang, C.; Liu, A.; Zhu, A.; Hu, J.; Lin, Y.; et al. Physiological and transcriptome analyses reveal short-term responses and formation of memory under drought stress in rice. Front. Genet. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Thu, N.B.A.; Nguyen, Q.T.; Hoang, X.L.T.; Thao, N.P.; Tran, L.S.P. Evaluation of drought tolerance of the Vietnamese soybean cultivars provides potential resources for soybean production and genetic engineering. BioMed Res. Int. 2014, 2014, 809736. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Iqbal, M.S.; Li, H.; Nazir, M.F.; Khalid, S.; Sarfraz, Z.; Hu, D.; Baojun, C.; Geng, X.; Tajo, S.M.; et al. Differential seedling growth and tolerance indices reflect drought tolerance in cotton. BMC Plant Biol. 2022, 22, 331. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef]
- Nxele, X.; Klein, A.; Ndimba, B.K. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 2017, 108, 261–266. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.; Xie, Y.; Su, L.; Zhang, R.; Wang, H.; Li, C.; Long, S. Drought resistance mechanisms of Phedimus aizoon L. Sci. Rep. 2021, 11, 13600. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Wang, X.; Li, X.; Dong, S. Physiology and metabonomics reveal differences in drought resistance among soybean varieties. Bot. Stud. 2022, 63, 8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, L.; Lai, J.; Zhao, H.; Song, W. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. BMC Plant Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Chaeikar, S.S.; Marzvan, S.; Khiavi, S.J.; Rahimi, M. Changes in growth, biochemical, and chemical characteristics and alteration of the antioxidant defense system in the leaves of tea clones (Camellia sinensis L.) under drought stress. Sci. Hortic. 2020, 265, 109257. [Google Scholar] [CrossRef]
- Mathobo, R.; Marais, D.; Steyn, J.M. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef]
- Gurumurthy, S.; Sarkar, B.; Vanaja, M.; Lakshmi, J.; Yadav, S.K.; Maheswari, M. Morpho-physiological and biochemical changes in black gram (Vigna mungo L. Hepper) genotypes under drought stress at flowering stage. Acta Physiol. Plant. 2019, 41, 42. [Google Scholar] [CrossRef]
- Stefanov, M.; Rashkov, G.; Borisova, P.; Apostolova, E. Sensitivity of the Photosynthetic Apparatus in Maize and Sorghum under Different Drought Levels. Plants 2023, 12, 1863. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Yang, S.L.; Dao, J.M.; Deng, J.; Shahzad, A.N.; Fan, X.; Li, R.D.; Quan, Y.J.; Bukhari, S.A.H.; Zeng, Z.H. Drought-induced alterations in photosynthetic, ultrastructural and biochemical traits of contrasting sugarcane genotypes. PLoS ONE 2020, 15, e0235845. [Google Scholar] [CrossRef]
- Hu, H.; Fei, X.; He, B.; Luo, Y.; Qi, Y.; Wei, A. Integrated analysis of metabolome and transcriptome data for uncovering flavonoid components of Zanthoxylum bungeanum maxim. leaves under drought stress. Front. Nutr. 2022, 8, 801244. [Google Scholar] [CrossRef]
- Pacheco, J.; Plazas, M.; Pettinari, I.; Landa-Faz, A.; González-Orenga, S.; Boscaiu, M.; Soler, S.; Prohens, J.; Vicente, O.; Gramazio, P. Moderate and severe water stress effects on morphological and biochemical traits in a set of pepino (Solanum muricatum) cultivars. Sci. Hortic. 2021, 284, 110143. [Google Scholar] [CrossRef]
- Zhang, P.; Bai, J.; Liu, Y.; Meng, Y.; Yang, Z.; Liu, T. Drought resistance of ten ground cover seedling species during roof greening. PLoS ONE 2020, 15, e0220598. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mild Drought Stress | Moderate Drought Stress | Severe Drought Stress | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Index | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Eigenvalues | 11.460 | 6.448 | 3.329 | 13.638 | 4.131 | 3.216 | 16.172 | 3.754 | 2.040 | |
| Contribution ratio/% | 49.827 | 28.035 | 14.475 | 59.295 | 17.963 | 13.983 | 70.313 | 16.322 | 8.868 | |
| Cumulative contribution ratio/% | 49.827 | 77.862 | 92.336 | 59.295 | 77.258 | 91.241 | 70.313 | 86.634 | 95.503 | |
| Eigenvector | RWC | 0.626 | −0.444 | −0.638 | 0.709 | −0.414 | −0.531 | 0.677 | −0.662 | −0.319 |
| DI | −0.707 | −0.411 | 0.008 | −0.625 | 0.253 | 0.689 | −0.867 | −0.210 | 0.228 | |
| SS | 0.952 * | 0.215 | 0.139 | 0.943 * | 0.159 | 0.289 | 0.943 * | 0.085 | −0.010 | |
| SP | 0.881 | 0.365 | 0.271 | 0.945 * | 0.172 | 0.279 | 0.977 * | 0.120 | 0.038 | |
| Pro | 0.561 | −0.758 | −0.169 | −0.032 | −0.461 | 0.781 | 0.667 | −0.727 | 0.165 | |
| REC | −0.740 | −0.087 | 0.603 | −0.609 | 0.710 | −0.325 | −0.546 | 0.774 | −0.317 | |
| MDA | −0.404 | −0.141 | 0.801 | −0.298 | 0.908 * | −0.282 | −0.306 | 0.821 | −0.454 | |
| SOD | 0.974 * | 0.077 | 0.171 | 0.908 * | 0.326 | 0.222 | 0.926 * | 0.359 | −0.103 | |
| POD | 0.756 | 0.640 | −0.021 | 0.585 | −0.395 | −0.183 | 0.306 | −0.158 | 0.881 | |
| CAT | 0.851 | 0.000 | 0.425 | 0.827 | 0.140 | 0.543 | 0.894 | 0.078 | 0.039 | |
| Pol | 0.064 | 0.797 | −0.599 | 0.727 | −0.573 | −0.321 | 0.957 * | −0.163 | −0.229 | |
| Fla | 0.791 | −0.537 | −0.221 | 0.545 | −0.354 | 0.712 | 0.948 * | −0.030 | 0.054 | |
| TS | 0.332 | −0.150 | 0.924 * | 0.225 | 0.879 | 0.236 | 0.242 | 0.720 | 0.650 | |
| ChlA | −0.266 | 0.953 * | 0.082 | 0.867 | 0.344 | −0.314 | 0.820 | 0.513 | 0.190 | |
| ChlB | −0.399 | 0.899 | 0.124 | 0.918 * | 0.363 | −0.133 | 0.952 * | 0.236 | 0.130 | |
| Car | −0.191 | 0.958 * | −0.007 | 0.813 | 0.009 | −0.386 | 0.967 * | −0.010 | −0.026 | |
| Total Chl | −0.270 | 0.914 * | 0.161 | 0.841 | 0.326 | 0.002 | 0.913 * | 0.321 | 0.222 | |
| Cv4CL1 | 0.963 * | 0.084 | 0.253 | 0.932 * | 0.310 | 0.056 | 0.926 * | 0.285 | −0.231 | |
| CvCAD1 | 0.972 * | 0.115 | 0.145 | 0.957 * | 0.028 | 0.287 | 0.981 * | 0.156 | 0.057 | |
| CvCAD2 | 0.374 | 0.643 | −0.399 | 0.654 | −0.487 | −0.303 | 0.850 | −0.438 | −0.202 | |
| CvPOX1 | 0.983 * | 0.155 | −0.022 | 0.943 * | 0.007 | 0.010 | 0.906 * | −0.032 | −0.275 | |
| CvPOX2 | 0.961 * | 0.105 | 0.170 | 0.985 * | −0.118 | −0.093 | 0.985 * | 0.070 | −0.071 | |
| CvPOX3 | 0.798 | −0.019 | 0.037 | 0.860 | 0.299 | −0.091 | 0.990 * | −0.129 | −0.043 | |
| Cultivars | X (Yak) | Ranking Value | Numerical Rank | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X (Ya1) | X (Ya2) | X (Ya3) | F Value | Mean F Value | ||||||||||
| Mild Drought | Moderate Drought | Severe Drought | Mild Drought | Moderate Drought | Severe Drought | Mild Drought | Moderate Drought | Severe Drought | Mild Drought | Moderate Drought | Severe Drought | |||
| Haida 1 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 0.00 | 1.00 | 1.00 | 0.85 | 1.00 | 0.95 | 1 |
| Haida 4 | 0.38 | 0.40 | 0.39 | 0.44 | 0.44 | 0.48 | 0.37 | 0.37 | 0.26 | 0.39 | 0.40 | 0.40 | 0.40 | 2 |
| Wanhai 1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 | 1.00 | 0.00 | 0.01 | 0.15 | 0.00 | 0.05 | 5 |
| Wanhai 3 | 0.06 | 0.03 | 0.12 | 0.03 | 0.11 | 0.15 | 0.06 | 0.79 | 0.02 | 0.05 | 0.16 | 0.11 | 0.11 | 4 |
| Wanhai 4 | 0.10 | 0.36 | 0.32 | 0.28 | 0.44 | 0.26 | 0.00 | 0.27 | 0.10 | 0.14 | 0.36 | 0.29 | 0.26 | 3 |
| Weights | 0.54 | 0.65 | 0.74 | 0.30 | 0.20 | 0.17 | 0.16 | 0.15 | 0.09 | |||||
| Stress Intensity | Principal Component Synthesis Model |
|---|---|
| Mild drought | F = −0.008V1 − 0.161V2 + 0.189V3 + 0.207V4 − 0.016V5 − 0.077V6 − 0.012V7 + 0.179V8 + 0.195V9 + 0.172V10 + 0.054V11 + 0.043V12 + 0.114V13 + 0.078V14 + 0.054V15 + 0.084V16 + 0.080V17 + 0.185V18 + 0.181V19 + 0.102V20 + 0.173V21 + 0.180V22 + 0.128V23 |
| Moderate drought | F = −0.005V1 + 0.007V2 + 0.187V3 + 0.188V4 − 0.005V5 − 0.011V6 + 0.067V7 + 0.201V8 + 0.010V9 + 0.189V10 − 0.007V11 + 0.089V12 + 0.185V13 + 0.151V14 + 0.177V15 + 0.086V16 + 0.172V17 + 0.187V18 + 0.169V19 − 0.004V20 + 0.139V21 + 0.118V22 + 0.162V23 |
| Severe drought | F = −0.005V1 + 0.007V2 + 0.187V3 + 0.188V4 − 0.005V5 − 0.011V6 + 0.067V7 + 0.201V8 + 0.010V9 + 0.189V10 − 0.007V11 + 0.089V12 + 0.185V13 + 0.151V14 + 0.177V15 + 0.086V16 + 0.172V17 + 0.187V18 + 0.169V19 − 0.004V20 + 0.139V21 + 0.118V22 + 0.162V23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, S.; Yan, W.; Xie, S.; Yu, J.; Yao, G.; Liu, Y.; Yang, D.; Wu, Y.; Yang, H. Comparison of Morphological, Physiological, and Related Gene Expression Responses to Drought Stress in Five Camellia vietnamensis Cultivars. Agronomy 2024, 14, 915. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050915
Shen S, Yan W, Xie S, Yu J, Yao G, Liu Y, Yang D, Wu Y, Yang H. Comparison of Morphological, Physiological, and Related Gene Expression Responses to Drought Stress in Five Camellia vietnamensis Cultivars. Agronomy. 2024; 14(5):915. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050915
Chicago/Turabian StyleShen, Shuaishuai, Wuping Yan, Shuao Xie, Jing Yu, Guanglong Yao, Ya Liu, Dongmei Yang, Yougen Wu, and Huageng Yang. 2024. "Comparison of Morphological, Physiological, and Related Gene Expression Responses to Drought Stress in Five Camellia vietnamensis Cultivars" Agronomy 14, no. 5: 915. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050915