


Organic vs. Integrated-Production Agriculture Farming: Which Grapevine Stress-Responsive Genes Are Affected by the Application of Resistance Inducers and Elicitors?
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Materials
2.2. Application of Plant Resistant Inducers
2.3. Sampling Collection
2.4. RNA Extraction and Complementary DNA Synthesis (cDNA)
2.5. qPCR Conditions for Gene Expression Analysis
2.6. Statistical Analysis
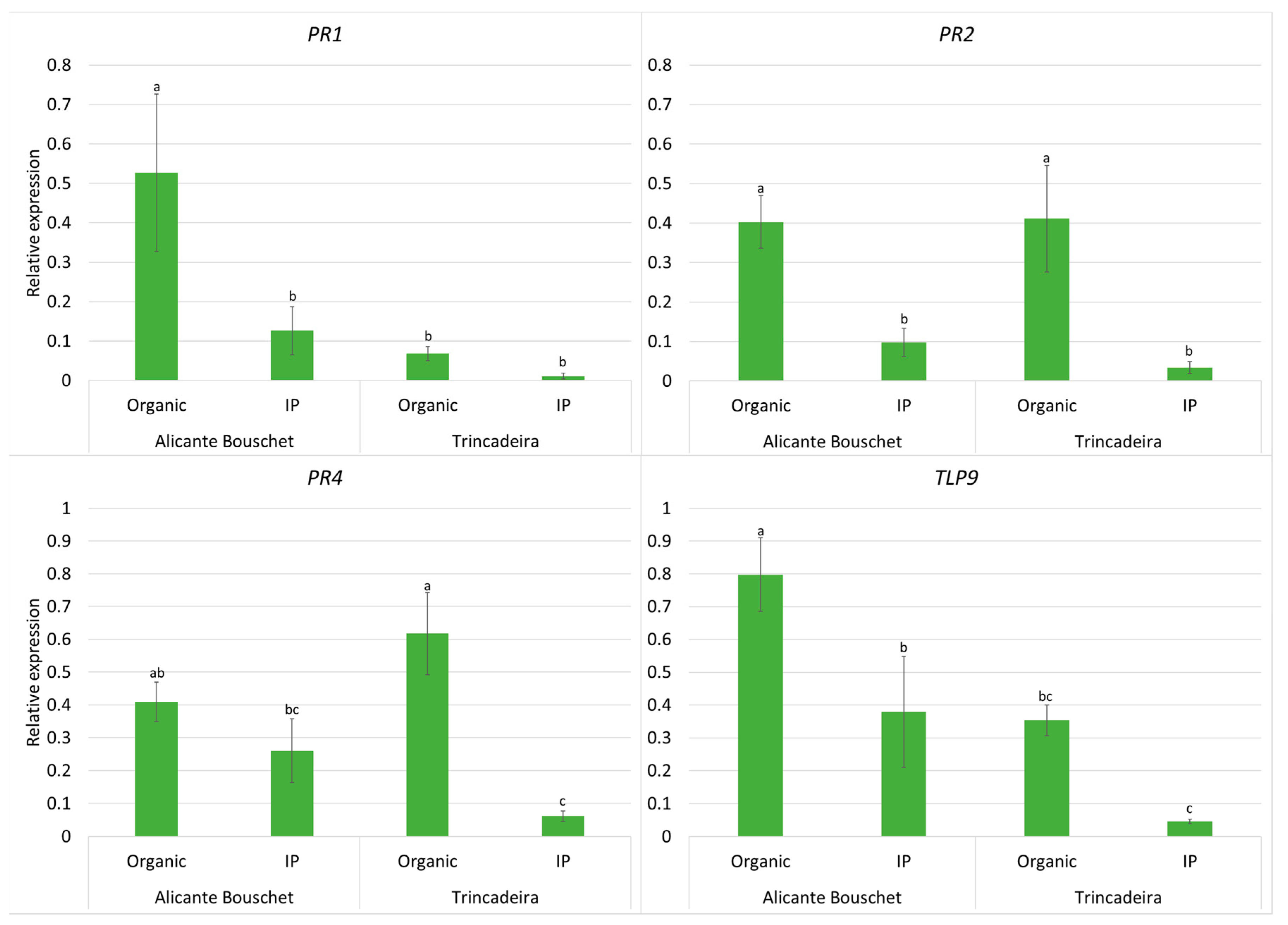
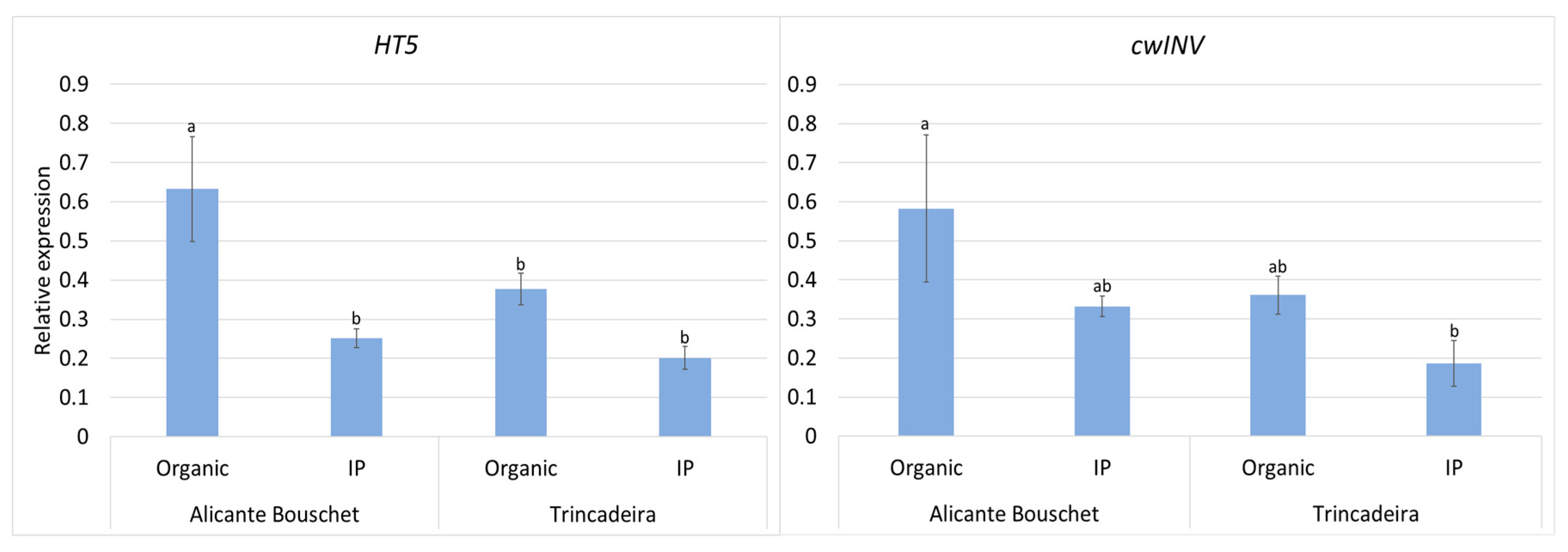
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CAP 2023-27—European Commission. Available online: https://agriculture.ec.europa.eu/common-agricultural-policy/cap-overview/cap-2023-27_en (accessed on 31 January 2024).
- Organic Action Plan—European Commission. Available online: https://agriculture.ec.europa.eu/farming/organic-farming/organic-action-plan_en (accessed on 31 January 2024).
- Sustainable Development Goals—European Commission. Available online: https://commission.europa.eu/strategy-and-policy/sustainable-development-goals_en (accessed on 13 March 2024).
- Vineyard Surface Area Worldwide 2022|Statista. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/240635/total-vineyard-areas-worldwide-and-in-europe/ (accessed on 18 October 2023).
- OIV. Focus OIV the World Organic Vineyard 2019; International Organisation of Vine and Wine: Paris, France, 2021; p. 11. [Google Scholar]
- Portal Do INE. Available online: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_indicadores&indOcorrCod=0000019&contexto=bd&selTab=tab2 (accessed on 18 October 2023).
- Portugal—Crop Trust. Available online: https://www.croptrust.org/pgrfa-hub/crops-countries-and-genebanks/countries/portugal/ (accessed on 18 October 2023).
- Wine Production Europe by Country 2022|Statista. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/445651/leading-countries-wine-production-europe/ (accessed on 18 October 2023).
- Integrated Production—Kiwa. Available online: https://www.kiwa.com/en/service2/inspection/integrated-production/ (accessed on 5 February 2024).
- Organic Agriculture: What Is Organic Agriculture? Available online: https://www.fao.org/organicag/oa-faq/oa-faq1/en/ (accessed on 5 February 2024).
- Wiesel, L.; Newton, A.C.; Elliott, I.; Booty, D.; Gilroy, E.M.; Birch, P.R.J.; Hein, I.; Bart, R.; Danforth Plant, D.; Ferrari, S. Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Front. Plant Sci. 2014, 5, 111484. [Google Scholar] [CrossRef] [PubMed]
- Reglinski, T.; Havis, N.; Rees, H.J.; de Jong, H. The Practical Role of Induced Resistance for Crop Protection. Phytopathology 2023, 113, 719–731. [Google Scholar] [CrossRef] [PubMed]
- La Torre, A.; Righi, L.; Iovino, V.; Battaglia, V. Evaluation of copper alternative products to control grape downy mildew in organic farming. J. Plant Pathol. 2019, 101, 1005–1012. [Google Scholar] [CrossRef]
- European Commission. Commission Implementing Regulation (EU) 218/1981; Official Journal of the European Union: Luxembourg, 2018; p. L317/16. [Google Scholar]
- Leonardi, F.; Mossi, G.; Camani, M. Il rame dei vigneti. Dati–Stat. E Soc. 2002, 3, 55–59. [Google Scholar]
- Komárek, M.; Čadková, E.; Chrastný, V.; Bordas, F.; Bollinger, J.C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Lencioni, V.; Grazioli, V.; Rossaro, B.; Bernabò, P. Transcriptional profiling induced by pesticides employed in organic agriculture in a wild population of Chironomus riparius under laboratory conditions. Sci. Total Environ. 2016, 557–558, 183–191. [Google Scholar] [CrossRef] [PubMed]
- La Torre, A.; Iovino, V.; Caradonia, F. Copper in plant protection: Current situation and prospects. Phytopathol. Mediterr. 2018, 57, 201–236. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/26507086 (accessed on 17 March 2024).
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Kaya, M.; Mujtaba, M.; Bulut, E.; Akyuz, B.; Zelencova, L.; Sofi, K. Fluctuation in physicochemical properties of chitins extracted from different body parts of honeybee. Carbohydr. Polymers 2015, 132, 9–16. [Google Scholar] [CrossRef]
- De Miccolis Angelini, R.M.; Angelini, M.; Rotolo, C.; Gerin, D.; Abate, D.; Pollastro, S.; Faretra, F. Global transcriptome analysis and differentially expressed genes in grapevine after application of the yeast-derived defense inducer cerevisane. Pest Manag. Sci. 2019, 75, 2020–2033. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; di Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, F.; Ippolito, A.; Di Venere, D.; Salerno, M. Effects of Pre- and Postharvest Chitosan Treatments to Control Storage Grey Mold of Table Grapes. J. Food Sci. 2002, 67, 1862–1867. [Google Scholar] [CrossRef]
- Singh, R.K.; Ruiz-May, E.; Rajput, V.D.; Minkina, T.; Gómez-Peraza, R.L.; Verma, K.K.; Shekhawat, M.S.; Pinto, C.; Falco, V.; Quiroz-Figueroa, F.R. Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices. Agriculture 2022, 12, 1369. [Google Scholar] [CrossRef]
- Llorens, E.; Mateu, M.; González-Hernández, A.I.; Agustí-Brisach, C.; García-Agustín, P.; Lapeña, L.; Vicedo, B. Extract of Mimosa tenuiflora and Quercus robur as potential eco-friendly management tool against Sclerotinia sclerotiorum in Lactuca sativa enhancing the natural plant defences. Eur. J. Plant Pathol. 2019, 153, 1105–1118. [Google Scholar] [CrossRef]
- Patanita, M.; do Rosário Félix, R.; Ribeiro, J.A.; Varanda CM, R.; Albuquerque, A.; Materatski, P.; Garrido, N.; Campos, M.D. Plant Pathogen Interaction Insights into Grapevine Defence Response Against Fungal Oomycete Diseases towards a Sustainable Plant Breeding; Verma, P.K., Mishra, S., Srivastava, V., Mehrotra, S., Eds.; Springer Nature: Singapore, 2023. [Google Scholar] [CrossRef]
- Alicante Bouschet—Vivai Cooperativi Rauscedo. Available online: https://vivairauscedo.com/contributi/en/schede/alicante-bouschet.pdf (accessed on 8 April 2024).
- Soares, B.; Pinto, T.; Leonor, P. Trincadeira. Available online: https://www.advid.pt/uploads/Trincadeira_Final_25.08.pdf (accessed on 8 April 2024).
- Idai COBRE. Available online: https://www.biotopio.gr/product/174/products-nutrition-biostimulants-idai-cobre-/eng (accessed on 22 February 2024).
- MIMETIC|Idai Nature. Available online: https://www.anagra.cl/Content/Documento/FichaTecnica/MIMETIC.pdf (accessed on 22 February 2024).
- Baslact Plus.|Hubel Verde. Available online: https://www.hubel.pt/pt/hv/products/nutricao_protecao_plantas/product/baslact-plus/ (accessed on 22 February 2024).
- Naturdai Carbobasic—Idai Nature. Available online: https://www.idainature.com/pt-pt/produtos/idai-nature/naturdai-carbobasic-sustancia-basica-mineral-com-acao-fungicida/ (accessed on 22 February 2024).
- Cardot, C.; Mappa, G.; la Camera, S.; Gaillard, C.; Vriet, C.; Lecomte, P.; Ferrari, G.; Coutos-Thévenot, P. Comparison of the Molecular Responses of Tolerant, Susceptible and Highly Susceptible Grapevine Cultivars During Interaction with the Pathogenic Fungus Eutypa lata. Front. Plant Sci. 2019, 10, 465699. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.F.; Fonseca, C.; Ferreira, R.B.; Lourenço, A.M.; Monteiro, S. Reference Gene Validation for Quantitative RT-PCR during Biotic and Abiotic Stresses in Vitis vinifera. PLoS ONE 2014, 9, 111399. [Google Scholar] [CrossRef] [PubMed]
- Mutawila, C.; Stander, C.; Halleen, F.; Vivier, M.A.; Mostert, L. Response of Vitis vinifera cell cultures to Eutypa lata and Trichoderma atroviride culture filtrates: Expression of defence-related genes and phenotypes. Protoplasma 2017, 254, 863–879. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qiao, H.; Zhang, X.; Guo, C.; Wang, M.; Wang, Y.; Wang, X. Analysis of the grape (Vitis vinifera L.) thaumatin-like protein (TLP) gene family and demonstration that TLP29 contributes to disease resistance. Sci. Rep. 2017, 7, 4269. [Google Scholar] [CrossRef]
- Hou, X.; Wei, L.; Xu, Y.; Khalil-Ur-rehman, M.; Feng, J.; Zeng, J.; Tao, J. Study on russet-related enzymatic activity gene expression in ‘Shine Muscat’ grape treated with, G.A.3. CPPU J. Plant Interact. 2018, 13, 195–202. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; Roy, N.; de Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, 341–3411. [Google Scholar] [CrossRef]
- Grape Varieties—CVRA. Available online: https://www.vinhosdoalentejo.pt/en/wines/grape-varieties/ (accessed on 13 March 2024).
- dos Santos, C.; Franco, O.L. Pathogenesis-Related Proteins (PRs) with Enzyme Activity Activating Plant Defense Responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan Oligomers and Copper Sulfate Induce Grapevine Defense Reactions and Resistance to Gray Mold and Downy Mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Mian, G.; Musetti, R.; Belfiore, N.; Boscaro, D.; Lovat, L.; Tomasi, D. Chitosan application reduces downy mildew severity on grapevine leaves by positively affecting gene expression pattern. Physiol. Mol. Plant Pathol. 2023, 125, 102025. [Google Scholar] [CrossRef]
- Repka, V. Elicitor-stimulated induction of defense mechanisms and defense gene activation in grapevine cell suspension cultures. Biol. Plant. 2001, 44, 555–565. [Google Scholar] [CrossRef]
- Chun, S.C.; Chandrasekaran, M. Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. Int. J. Biol. Macromol. 2019, 125, 948–954. [Google Scholar] [CrossRef] [PubMed]
- López-Velázquez, J.C.; García-Morales, S.; López-Sánchez, G.P.; Montero-Cortés, M.I.; Uc-Várguez, A.; Qui-Zapata, J.A. High-Density Chitosan Induces a Biochemical and Molecular Response in Coffea arabica during Infection with Hemileia vastatrix. Int. J. Mol. Sci. 2023, 24, 16165. [Google Scholar] [CrossRef]
- Yan, W.; Wu, X.; Li, Y.; Liu, G.; Cui, Z.; Jiang, T.; Ma, Q.; Luo, L.; Zhang, P. Cell wall invertase 3 affects cassava productivity via regulating sugar allocation from source to sink. Front. Plant Sci. 2019, 10, 445753. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.P.; Bonghi, C.; Varotto, S.; Battistelli, A.; Burbidge, C.A.; Castellarin, S.D.; Chen, Z.H.; Darriet, P.; Moscatello, S.; Rienth, M.; et al. Sucrose metabolism and transport in grapevines, with emphasis on berries and leaves and insights gained from a cross-species comparison. Int. J. Mol. Sci. 2021, 22, 7794. [Google Scholar] [CrossRef]
- Pagliarani, C.; Moine, A.; Chitarra, W.; Meloni, G.R.; Abbà, S.; Nerva, L.; Pugliese, M.; Gullino, M.L.; Gambino, G. The Molecular Priming of Defense Responses is Differently Regulated in Grapevine Genotypes Following Elicitor Application against Powdery Mildew. Int. J. Mol. Sci. 2020, 21, 6776. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Chen, Z. Functional Analysis of the Arabidopsis PAL Gene Family in Plant Growth, Development, and Response to Environmental Stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Ma, W.; Qayyum, Z.; Noureen, F.; Khan, M.; Amir, R.; Khan, M.; Haider, G.; Munir, F.; Gul, A. Identification and Expression Analysis of Stilbene Synthase Genes in Arachis hypogaea in Response to Methyl Jasmonate and Salicylic Acid Induction. Plants 2022, 11, 1776. [Google Scholar] [CrossRef] [PubMed]
- Ramaroson, M.L.; Koutouan, C.; Helesbeux, J.J.; le Clerc, V.; Hamama, L.; Geoffriau, E.; Briard, M. Role of Phenylpropanoids and Flavonoids in Plant Resistance to Pests and Diseases. Molecules 2022, 27, 8371. [Google Scholar] [CrossRef] [PubMed]
- Miliordos, D.E.; Alatzas, A.; Kontoudakis, N.; Kouki, A.; Unlubayir, M.; Gémin, M.P.; Tako, A.; Hatzopoulos, P.; Lanoue, A.; Kotseridis, Y. Abscisic Acid and Chitosan Modulate Polyphenol Metabolism and Berry Qualities in the Domestic White-Colored Cultivar Savvatiano. Plants 2022, 11, 1648. [Google Scholar] [CrossRef] [PubMed]
- Rühmann, S.; Pfeiffer, J.; Brunner, P.; Szankowski, I.; Fischer, T.C.; Forkmann, G.; Treutter, D. Induction of stilbene phytoalexins in grapevine (Vitis vinifera) and transgenic stilbene synthase-apple plants (Malus domestica) by a culture filtrate of Aureobasidium pullulans. Plant Physiol. Biochem. 2013, 72, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Arif, Y.; Miszczuk, E.; Bajguz, A.; Hayat, S. Specific Roles of Lipoxygenases in Development and Responses to Stress in Plants. Plants 2022, 11, 979. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kaothien, P.; Matsui, T.; Kawaoka, A.; Shinmyo, A. Molecular biology and application of plant peroxidase genes. Appl. Microbiol. Biotechnol. 2003, 60, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Ros Barceló, A.; Pomar, F.; López-Serrano, M.; Pedreño, M.A. Peroxidase: A multifunctional enzyme in grapevines. Funct. Plant Biol. 2003, 30, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan stimulates defense reactions in grapevine leaves and inhibits development of Botrytis cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Gauthier, A.; Trouvelot, S.; Kelloniemi, J.; Frettinger, P.; Wendehenne, D.; Daire, X.; Joubert, J.M.; Ferrarini, A.; Delledonne, M.; Flors, V.; et al. The Sulfated Laminarin Triggers a Stress Transcriptome before Priming the SA- and ROS-Dependent Defenses during Grapevine’s Induced Resistance against Plasmopara viticola. PLoS ONE 2014, 9, e88145. [Google Scholar] [CrossRef]
- Qin, F.; Yu, B.; Li, W. Heat shock protein 101 (HSP101) promotes flowering under nonstress conditions. Plant Physiol. 2021, 186, 407–419. [Google Scholar] [CrossRef]
- Xie, C.; Yang, L.; Gai, Y. MAPKKKs in Plants: Multidimensional Regulators of Plant Growth and Stress Responses. Int. J. Mol. Sci. 2023, 24, 4117. [Google Scholar] [CrossRef] [PubMed]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: Striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef] [PubMed]
- Micronutrientes IDAINATURE Naturdai Mimetic 250 cc—Comercial Mida. Available online: https://comercialmida.pt/products/micronutrientes-naturdai-mimetic-idainature (accessed on 14 March 2024).
- Patanita, M.; Albuquerque, A.; Campos, M.D.; Materatski, P.; Varanda CM, R.; Ribeiro, J.A.; Félix, M.D.R. Metagenomic Assessment Unravels Fungal Microbiota Associated to Grapevine Trunk Diseases. Horticulturae 2022, 8, 288. [Google Scholar] [CrossRef]
- Campos, M.D.; do Rosário Félix, R.; Patanita, M.; Materatski, P.; Varanda, C. High throughput sequencing unravels tomato-pathogen interactions towards a sustainable plant breeding. Hortic. Res. 2021, 8, 171. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.D.; do Rosário Félix, R.; Patanita, M.; Materatski, P.; Albuquerque, A.; Ribeiro, J.A.; Varanda, C. Defense Strategies: The Role of Transcription Factors in Tomato–Pathogen Interaction. Biology 2022, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.A.; Albuquerque, A.; Materatski, P.; Patanita, M.; Varanda, C.M.R.; do Rosário Félix, R.; Campos, M.D. Tomato Response to Fusarium spp. Infection under Field Conditions: Study of Potential Genes Involved. Horticulturae 2022, 8, 433. [Google Scholar] [CrossRef]
- Zhu, F.; Cao, M.Y.; Zhang, Q.P.; Mohan, R.; Schar, J.; Mitchell, M.; Chen, H.; Liu, F.; Wang, D.; Fu, Z.Q. Join the green team: Inducers of plant immunity in the plant disease sustainable control toolbox. J. Adv. Res. 2024, 57, 15–42. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | AL (bp) | NCBI Accession ID | E (%) | Ref. |
|---|---|---|---|---|---|
| GAPDH | Fw: CCACAGACTTCATCGGTGACA Rv: TTCTCGTTGAGGGCTATTCCA | 70 | XM_002263109.3 | 96.3 | [33] |
| PEP | Fw: CCTCCTCCTCCAGATTGC Rv: GGCTTGCTTGATTCCATTATC | 198 | AF236126.1 | 95.0 | [34] |
| UBC | Fw: CATAAGGGCTATCAGGAGGAC Rv: TGGCGGTCGGAGTTAGG | 161 | EE253706 | 106.6 | [34] |
| Gene | Primer Sequence (5′→3′) | AL (bp) | NCBI Accession ID | E (%) | Ref. |
|---|---|---|---|---|---|
| PR1 | Fw: GCAACTATATCGGACAACGTCCTT Rv: TCACCATGCTCTAACAGTACCCA | 80 | XM_002273752 | 97.4 | [35] |
| PR2 | Fw: GCAGTCGGGAACGAAGTGAG Rv: ATGGAGGGTAGGAGTTGCCC | 172 | NM_001280967.2 | 116.9 | [33] |
| PR4 | Fw: GCCCAGAGCGCCAGCAATGT Rv: CGCCATGCCAAGGGCTTGCT | 125 | XM_002264684 | 104.7 | [35] |
| TLP9 | Fw: TGCAGCAACCTTCAACATC Rv: GTGGCGGCCTTCACAT | 120 | XM_002276395.4 | 110.1 | [36] |
| HT5 | Fw: TAGTGATGCGTCCCTCTACTC Rv: CTTCCAGCAAGAGCAATCGAC | 113 | NM_001281278.1 | 108.3 | [33] |
| cwINV | Fw: ACGAATCATCTAGTGTGGAGCAC Rv: CTTAAACGATATCTCCACATCTGC | 236 | NM_001281279.1 | 92.2 | [33] |
| PAL | Fw: TGCTGACTGGTGAAAAGGTG Rv: CGTTCCAAGCACTGAGACAA | 114 | XM_003635609.3 | 107.2 | [33] |
| STS1 | Fw: AGGGAAGCAGCATTGAAGGC Rv: CGGGCATTTCTACACCGGAG | 97 | XM_002263845.4 | 94.8 | [33] |
| LOX | Fw: TGCTCTACCCCACAAGCGAA Rv: AGCAGTGTGCTCATGATTTTCCAG | 95 | NM_001281249.1 | 109.3 | [33] |
| PER42 | Fw: CTTGTGAGAGGTATGAAGATG Rv: ACCATAACGCCATTGTAAC | 193 | XM_002274733.3 | 95.1 | [37] |
| HSP101 | Fw: AATGAGACTCTTGCTGGGGC Rv: CAGCACCGATTATGGCTTGC | 130 | NM_001280893.1 | 106.8 | This study |
| MAPKKK17 | Fw: ACCTTAGGCTCTGGCTCCTC Rv: CACACCCCTTGTAGCCAACT | 169 | XM_002269624.3 | 116.5 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, T.; Patanita, M.; Félix, M.d.R.; Albuquerque, A.; Ribeiro, J.A.; Santos, F.; Basaloco, M.; da Rosa, A.M.; Campos, M.D. Organic vs. Integrated-Production Agriculture Farming: Which Grapevine Stress-Responsive Genes Are Affected by the Application of Resistance Inducers and Elicitors? Agronomy 2024, 14, 892. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050892
Monteiro T, Patanita M, Félix MdR, Albuquerque A, Ribeiro JA, Santos F, Basaloco M, da Rosa AM, Campos MD. Organic vs. Integrated-Production Agriculture Farming: Which Grapevine Stress-Responsive Genes Are Affected by the Application of Resistance Inducers and Elicitors? Agronomy. 2024; 14(5):892. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050892
Chicago/Turabian StyleMonteiro, Tomás, Mariana Patanita, Maria do Rosário Félix, André Albuquerque, Joana A. Ribeiro, Filipa Santos, Margarida Basaloco, Augusto Maria da Rosa, and Maria Doroteia Campos. 2024. "Organic vs. Integrated-Production Agriculture Farming: Which Grapevine Stress-Responsive Genes Are Affected by the Application of Resistance Inducers and Elicitors?" Agronomy 14, no. 5: 892. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14050892