
An Evaluation of Bacterial Wilt (Ralstonia solanacearum) Resistance in a Set of Tomato Germplasm from the United States Department of Agriculture †


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Bacterial Wilt Pathogen
2.2. Greenhouse Experiment for Pathogen Tests
2.3. Phenotypic Data Analysis
2.3.1. Statistical Model
2.3.2. ANOVA, Distribution, Descriptive Statistics, Pearson’s Correlation, and Broad-Sense Heritability
2.4. Genetic Diversity Analysis
2.4.1. DNA Extraction: Genotyping by Sequencing (GBS) and SNP Discovery
2.4.2. Principal Component Analysis (PCA) and Genetic Diversity
3. Results
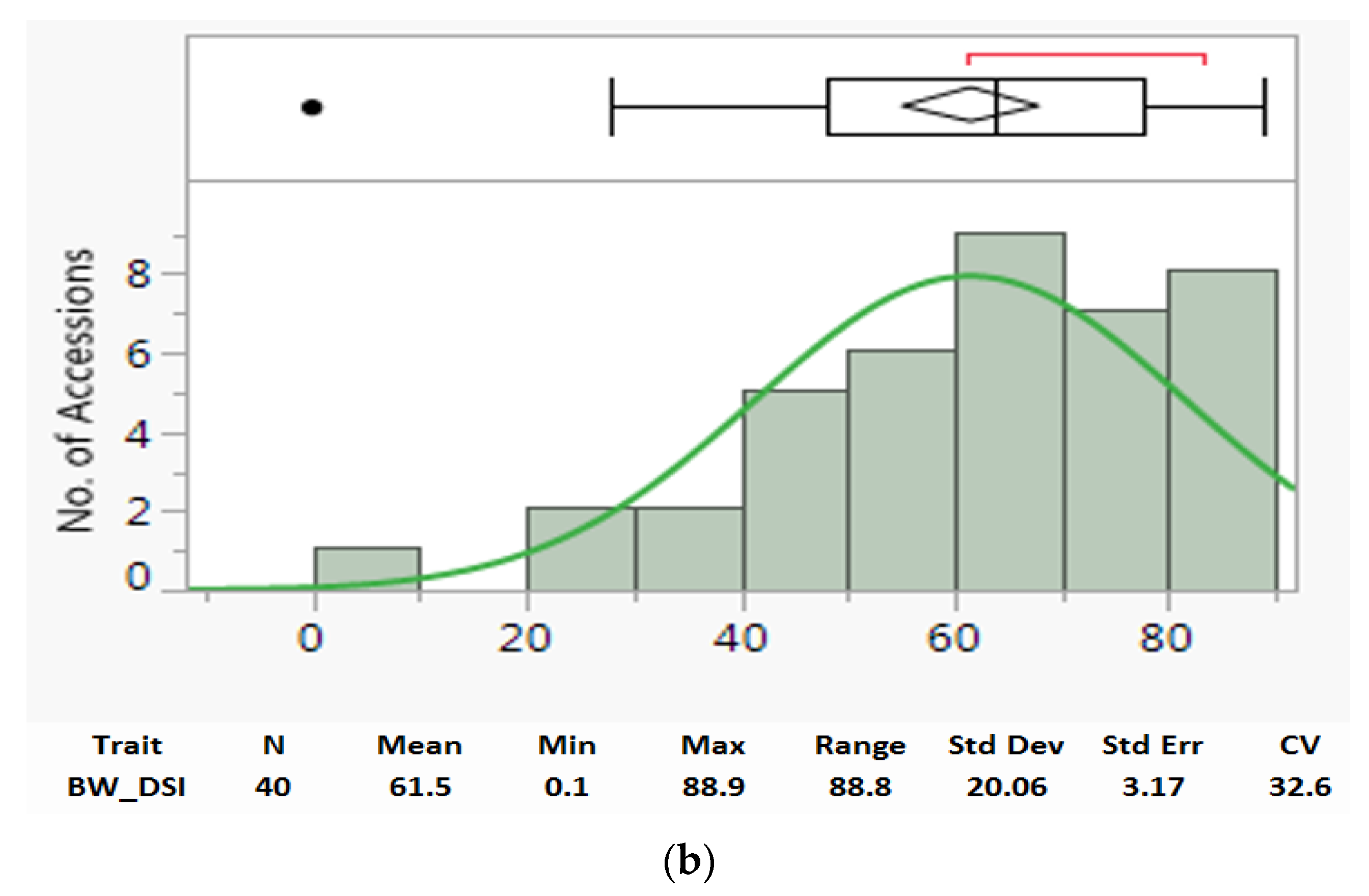
3.1. Parameters and Distributions of Bacterial Wilt Resistance
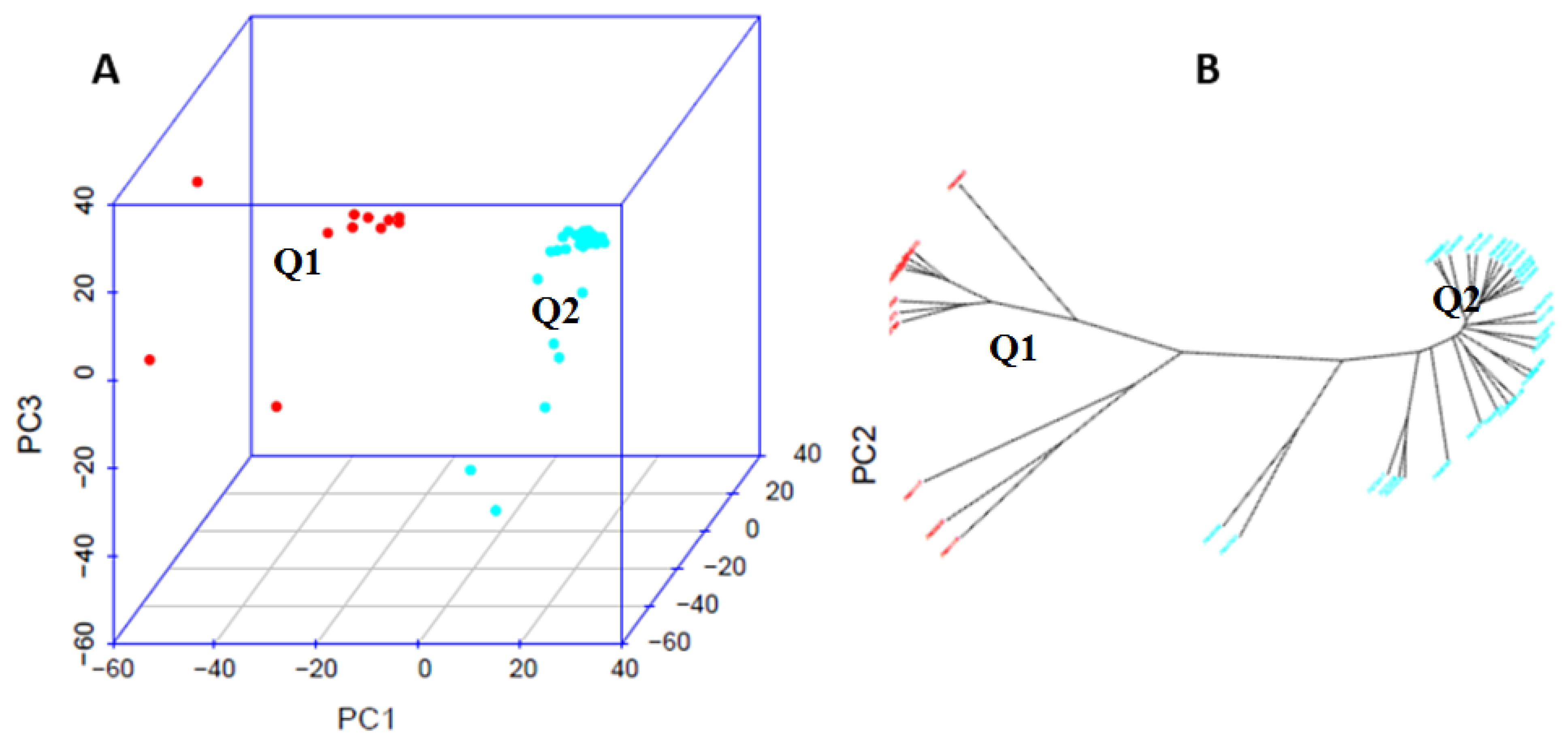
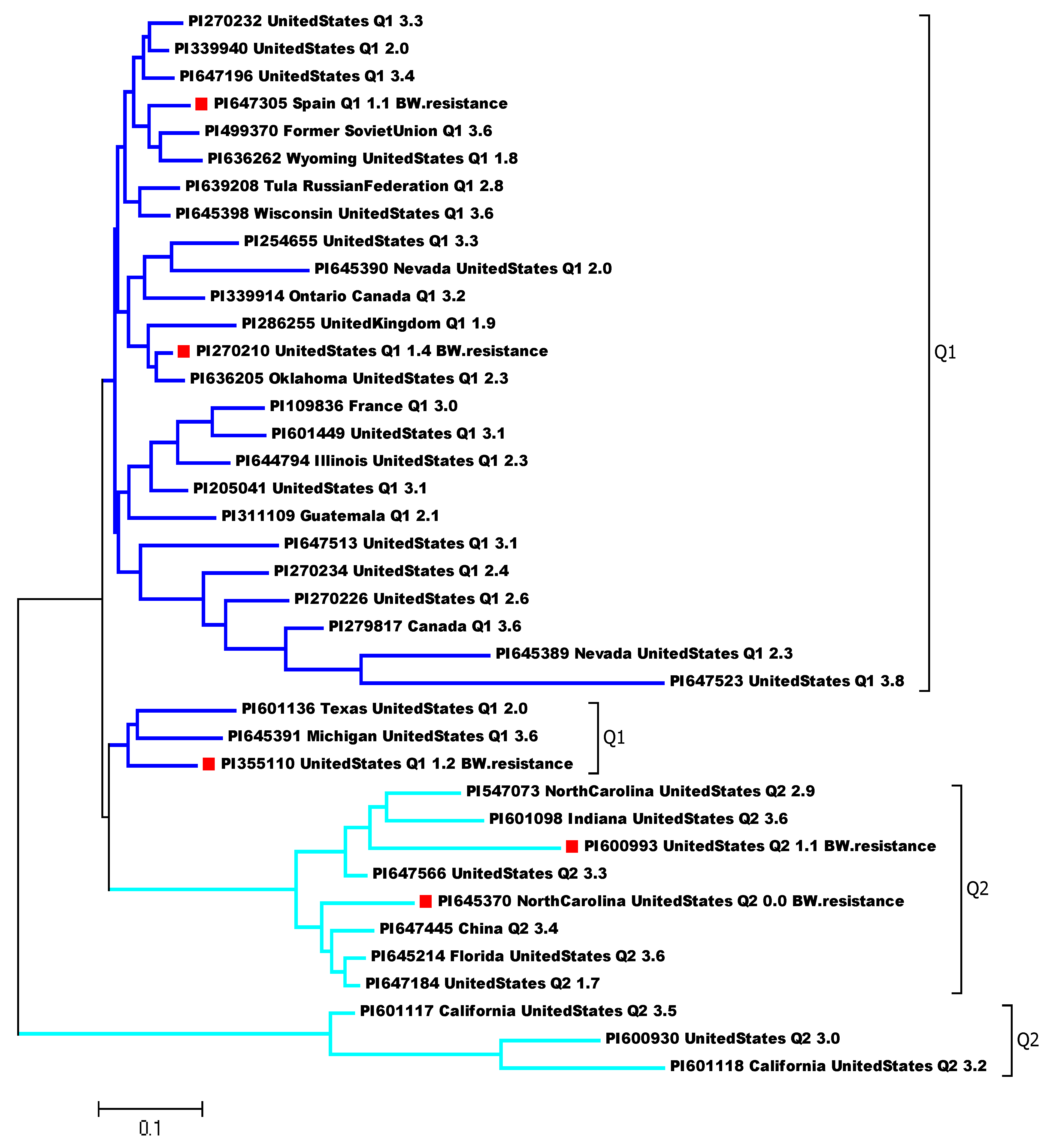
3.2. Principal Component Analysis (PCA) and Genetic Diversity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mohan, V.; Gupta, S.; Thomas, S.; Mickey, H.; Charakana, C.; Chauhan, V.S.; Sharma, K.; Kumar, R.; Tyagi, K.; Sarma, S.; et al. Tomato fruits show wide phenomic diversity but fruit developmental genes show low genomic diversity. PLoS ONE 2016, 11, e0152907. [Google Scholar] [CrossRef] [PubMed]
- Perveen, R.; Suleria HA, R.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Ahmad, S. Tomato (Solanum lycopersicum) carotenoids and lycopenes chemistry; metabolism, absorption, nutrition, and allied health claims—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2015, 55, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Hur, O.S.; Ro, N.Y.; Ko, H.C.; Rhee, J.H.; Sung, J.S.; Ryu, K.Y.; Lee, S.Y.; Baek, H.J. Evaluation of resistance to Ralstonia solanacearum in tomato genetic resources at seedling stage. Plant Pathol. J. 2016, 32, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H.; Rosado, P.; Roser, M. Agricultural Production. Our World in Data. 2023. Available online: https://ourworldindata.org/agricultural-production (accessed on 22 December 2023).
- Collins, E.J.; Bowyer, C.; Tsouza, A.; Chopra, M. Tomatoes: An extensive review of the associated health impacts of tomatoes and factors that can affect their cultivation. Biology 2022, 11, 239. [Google Scholar] [CrossRef] [PubMed]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.J.; Tucker, E.J.; Tester, M. Genetic analysis of abiotic stress tolerance in crops. Curr. Opin. Plant Biol. 2011, 14, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, M.R.; Ayyub, C.M.; Amjad, M.; Waraich, E.A. Morpho-physiological evaluation of tomato genotypes under high-temperature stress conditions. J. Sci. Food Agric. 2016, 96, 2698–2704. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, Z.; Wang, X.; Rosenqvist, E.; Wang, Y.; Zhao, T.; Ottosen, C.O. Evaluation of temperature stress tolerance in cultivated and wild tomatoes using photosynthesis and chlorophyll fluorescence. Hortic. Environ. Biotechnol. 2018, 59, 499–509. [Google Scholar] [CrossRef]
- Hong, J.K.; Jang, S.J.; Lee, Y.H.; Jo, Y.S.; Yun, J.G.; Jo, H.; Park, C.J.; Kim, H.J. Reduced bacterial wilt in tomato plants by bactericidal peroxyacetic acid mixture treatment. Plant Pathol. J. 2018, 34, 78–84. [Google Scholar] [CrossRef]
- Lee, J.H.; Jang, K.S.; Choi, Y.H.; Kim, J.-C.; Choi, G.J. Development of an efficient screening system for resistance of tomato cultivars to Ralstonia solanacearum. Res. Plant Dis. 2015, 21, 290–296. [Google Scholar] [CrossRef]
- Yuliar; Asi Nion, Y.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Gautam, R.K.; Singh, D.R.; Sharma, T.V.R.S.; Sakthivel, K.; Roy, S.D. Genetic approaches for mitigating losses caused by bacterial wilt of tomato in tropical islands. Eur. J. Plant Pathol. 2015, 143, 205–221. [Google Scholar] [CrossRef]
- Bindal, S.; Srivastava, S. Bacterial wilt of solanaceous crops: Sign, symptoms and management. Agrica 2019, 8, 134. [Google Scholar] [CrossRef]
- Álvarez, B.; López, M.M.; Biosca, E.G. Influence of native microbiota on survival of Ralstonia solanacearum phylotype II in river water microcosms. Appl. Environ. Microbiol. 2007, 73, 7210–7217. [Google Scholar] [CrossRef] [PubMed]
- Prior, P. How Complex Is the “Ralstonia solanacearum Species Complex? 2005. Available online: https://www.researchgate.net/publication/37628297 (accessed on 22 December 2023).
- Lebeau, A.; Daunay, M.C.; Frary, A.; Palloix, A.; Wang, J.F.; Dintinger, J.; Chiroleu, F.; Wicker, E.; Prior, P. Bacterial wilt resistance in tomato, pepper, and eggplant: Genetic resources respond to diverse strains in the Ralstonia solanacearum species complex. Phytopathology 2011, 101, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Kong, J.; Cui, D.; Zhao, H.; Niu, Y.; Xu, M.; Jiang, G.; Zhao, Y.; Wang, W. Resistance against Ralstonia solanacearum in tomato depends on the methionine cycle and the γ-aminobutyric acid metabolic pathway. Plant J. 2019, 97, 1032–1047. [Google Scholar] [CrossRef] [PubMed]
- Muthoni, J.; Shimelis, H.; Melis, R. Management of bacterial wilt [Rhalstonia solanacearum] of potatoes: Opportunity for host resistance in Kenya. J. Agric. Sci. 2012, 4, 621–625. [Google Scholar] [CrossRef]
- Aslam, M.N.; Mukhtar, T.; Hussain, M.A.; Raheel, M. Assessment of resistance to bacterial wilt incited by Ralstonia solanacearum in tomato germplasm. J. Plant Dis. Prot. 2017, 124, 585–590. [Google Scholar] [CrossRef]
- Champoiseau, P.G.; Jones, J.B.; Allen, C. Ralstonia solanacearum Race 3 Biovar 2 causes tropical losses and temperate anxieties. Plant Health Progress 2009, 10. [Google Scholar] [CrossRef]
- Hong, J.C.; Norman, D.J.; Reed, D.L.; Timur Momol, M.; Jones, J.B. Diversity among Ralstonia solanacearum strains isolated from the southeastern United States. Phytopathology 2012, 102, 924–936. [Google Scholar] [CrossRef]
- Ji, P.; Allen, C.; Sanchez-Perez, A.; Yao, J.; Elphinstone, J.G.; Jones, J.B.; Momol, M.T. New diversity of Ralstonia solanacearum strains associated with vegetable and ornamental crops in Florida. Plant Dis. 2007, 91, 195–203. [Google Scholar] [CrossRef]
- Du, H.; Wen, C.; Zhang, X.; Xu, X.; Yang, J.; Chen, B.; Geng, S. Identification of a major qtl (Qrrs-10.1) that confers resistance to Ralstonia solanacearum in pepper (Capsicum annuum) using slaf-bsa and qtl mapping. Int. J. Mol. Sci. 2019, 20, 5887. [Google Scholar] [CrossRef]
- Mcavoy, T.; Freeman, J.H.; Rideout, S.L.; Olson, S.M.; Paret, M.L. Evaluation of grafting using hybrid rootstocks for management of bacterial wilt in field tomato production. Hortscience 2012, 47, 621–625. [Google Scholar] [CrossRef]
- Lu, Y.; Rao, S.; Huang, F.; Cai, Y.; Wang, G.; Cai, K. Effects of biochar amendment on tomato bacterial wilt resistance and soil microbial amount and activity. Int. J. Agron. 2016, 2016, 2938282. [Google Scholar] [CrossRef]
- Huet, G. Breeding for resistance to Ralstonia solanacearum. Front. Plant Sci. 2014, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Asati, R.; Patel, V.; Sikarwar, R.S.; Payasi, D.K. Breeding and genomic approaches towards the development of fusarium wilt resistance in chickpea. Life 2023, 13, 988. [Google Scholar] [CrossRef] [PubMed]
- Barik, S.; Reddy, A.C.; Ponnam, N.; Kumari, M.; DC, L.R.; Petikam, S.; Petikam, S.; Gs, S. Breeding for bacterial wilt resistance in eggplant (Solanum melongena L.): Progress and prospects. Crop Prot. 2020, 137, 105270. [Google Scholar] [CrossRef]
- Fock, I.; Collonnier, C.; Purwito, A.; Luisetti, J.; Souvannavong, V.; Vedel, F.; Servaes, A.; Ambroise, A.; Kodja, H.; Ducreux, G.; et al. Resistance to bacterial wilt in somatic hybrids between Solanum tuberosum and Solanum phureja. Plant Sci. 2000, 160, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.U.; Gopal, J.; Singh, B.P. Improvement for bacterial wilt resistance in potatoes by conventional and biotechnological approaches. Agric. Res. 2012, 1, 299–316. [Google Scholar] [CrossRef]
- Hanson, P.M.; Licardo, O. Diallel Analysis of Bacterial Wilt Resistance in Tomato Derived from Different Sources. 1998. Available online: https://apsjournals.apsnet.org/doi/pdf/10.1094/PDIS.1998.82.1 (accessed on 22 December 2023).
- Chellemi, D.O.; Dankers, H.A.; Olson, S.M.; Hodge, N.C.; Scott, J.W. Evaluating bacterial wilt-resistant tomato genotypes using a regional approach. J. Amer. Soc. Hort. Sci 1994, 119, 325–329. Available online: https://journals.ashs.org/jashs/view/journals/jashs/119/2/article-p325.xml (accessed on 22 December 2023). [CrossRef]
- Zohoungbogbo, H.; Quenum, A.; Honfoga, J.; Chen, J.R.; Achigan-Dako, E.; Kenyon, L.; Hanson, P. Evaluation of resistance sources of tomato (Solanum lycopersicum L.) to phylotype I strain of Ralstonia solanacearum species complex in Benin. Agronomy 2021, 11, 1513. [Google Scholar] [CrossRef]
- Newton B-Mensah, I.; Osei, K.; Prempeh, R.N.A. Screening tomato genotypes for bacterial wilt disease (Ralstonia solanacearum) resistance in Ghana. Eur. J. Agric. Food Sci. 2021, 3, 1–8. [Google Scholar] [CrossRef]
- Kumar, S.; Ramanjini Gowda, P.H.; Saikia, B.; Debbarma, J.; Velmurugan, N.; Chikkaputtaiah, C. Screening of tomato genotypes against bacterial wilt (Ralstonia solanacearum) and validation of resistance-linked DNA markers. Australas. Plant Pathol. 2018, 47, 365–374. [Google Scholar] [CrossRef]
- Makawa Phiri, T. Screening a Set of Tomato Genotypes for Resistance to Bacterial Wilt and a Genomic Wide Association Study and Genomic Prediction for Yield-Related Traits in ‘L aits In ‘LCP 85-384′. I. Sugar CP 85-384′. I. Sugarcane. Master’s Thesis, University of Arkansas, Fayetteville, AR, USA, 2023. Available online: https://scholarworks.uark.edu/etd/4912 (accessed on 22 December 2023).
- Bocsanczy, A.M.; Espindola, A.S.; Norman, D.J. Whole-genome sequences of Ralstonia solanacearum strains P816, P822, and P824, emerging pathogens of blueberry in Florida. Microbiol. Resour. Announc. 2019, 8, e01316-18. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.; Jones, J.B.; Harmon, C.; Momol, T.M. R. solanacearum/Culture Media. 2007. Available online: https://plantpath.ifas.ufl.edu/rsol/Culturemedia_PrinterText.html (accessed on 22 December 2023).
- Kalpage, M.D.; De Costa, D.M. Isolation of bacteriophages and determination of their efficiency in controlling Ralstonia solanacearum causing bacterial wilt of tomato. Trop. Agric. Res. 2014, 26, 28. Available online: https://pdfs.semanticscholar.org/33c3/492b6bd7c418b060695ed4ea61fafb0b69ff.pdf (accessed on 22 December 2023). [CrossRef]
- Bi-Hao, C.; Jian-Jun, L.; Yong, W.; Guo-Ju, C. Inheritance and identification of SCAR marker linked to bacterial wilt-resistance in eggplant. Afr. J. Biotechnol. 2009, 8, 5201–5207. Available online: http://www.academicjournals.org/AJB (accessed on 22 December 2023).
- Kwon, J.S.; Nam, J.Y.; Yeom, S.I.; Kang, W.H. Leaf-to-whole plant spread bioassay for pepper and Ralstonia solanacearum interaction determines the inheritance of resistance to bacterial wilt for further breeding. Int. J. Mol. Sci. 2021, 22, 2279. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.S.; Liu, H.I.; Bock, C.H. A discussion on disease severity index values. Part I: Warning on inherent errors and suggestions to maximize accuracy. Ann. Appl. Biol. 2017, 171, 139–154. [Google Scholar] [CrossRef]
- Holland, J. Estimating and interpreting heritability for plant breeding: An update. Plant Breed. Rev. 2003, 22, 9–112. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e0019379. [Google Scholar] [CrossRef]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A High Capacity Genotyping by Sequencing Analysis Pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef]
- Rochette, N.C.; Rivera-Colón, A.G.; Catchen, J.M. Stacks 2: Analytical methods for paired-end sequencing improve RADseq-based population genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. BLINK: A package for the next level of genome-wide association studies with both individuals and markers in the millions. GigaScience 2019, 8, giy154. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hong Hai, T.T.; Esch, E.; Wang, J.F. Resistance to Taiwanese Race 1 strains of Ralstonia solanacearum in wild tomato germplasm. Eur. J. Plant Pathol. 2008, 122, 471–479. [Google Scholar] [CrossRef]
- Morel, A.; Peeters, N.; Vailleau, F.; Barberis, P.; Jiang, G.; Berthomé, R.; Guidot, A. Plant pathogenicity phenotyping of Ralstonia solanacearum strains. Methods Mol. Biol. 2018, 1734, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Deslandes, L.; Olivier, J.; Theulières, F.; Hirsch, J.; Feng, D.X.; Bittner-Eddy, P.; Beynon, J.; Marco, Y. Resistance to Ralstonia solanacearum in Arabidopsis thaliana is conferred by the recessive RRS1-R gene, a member of a novel family of resistance genes. Proc. Natl. Acad. Sci. USA 2002, 99, 2404. [Google Scholar] [CrossRef] [PubMed]
- Grimault, V.; Prior, P.; Anaïs, G. A monogenic dominant resistance of tomato to bacterial wilt in hawaii 7996 is associated with plant colonization by Pseudomonas solanacearum. J. Phytopathol. 1995, 143, 349–352. [Google Scholar] [CrossRef]
- Kaushik, S.K. Genetics of fruit yield and its contributing characteristics in tomato (Solanum lycopersicom). J. Agric. Biotechnol. Sustain. Dev. 2011, 3, 209. [Google Scholar] [CrossRef]
- Boakye-Mensah, N.I. Evaluation of Tomato (Solanum lycopersicum L.) Genotypes for Bacterial Wilt (Ralstonia solanacearum) Resistance in Ghana. 2020. Available online: https://ir.ucc.edu.gh/xmlui (accessed on 22 December 2023).
- Schmidt, P.; Hartung, J.; Bennewitz, J.; Hans-Peter, P. Heritability in plant breeding on a genotype-difference basis. Genetics 2019, 212, 991–1008. [Google Scholar] [CrossRef]
- Maji, A.T. Application of principal component analysis for rice germplasm characterization and evaluation. J. Plant Breed. Crop Sci. 2012, 4, 87–93. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ACCESSION | NAME | ORIGIN | 2_Cluster | BW_Score (Order by Lower Scale, Higher Resistant) | BW Score Significant at p = 0.05 Level | BW_DSI % | BW DSI Significant at p = 0.05 Level |
|---|---|---|---|---|---|---|---|
| PI645370 | Venus | North Carolina, United States | Q2 | 0.0 | I | 0.0 | G |
| PI600993 | Liberator | United States | Q2 | 1.1 | HI | 27.8 | FG |
| PI647305 | Rosa o Monserrat | Spain | Q1 | 1.1 | HI | 27.8 | FG |
| PI355110 | Napoli | United States | Q1 | 1.2 | GHI | 30.6 | EFG |
| PI270210 | Sioux | United States | Q1 | 1.4 | FGHI | 36.1 | DEFG |
| PI647184 | Creole | United States | Q2 | 1.7 | EFGH | 41.7 | CDEFG |
| PI636262 | Favorite | Wyoming, United States | Q1 | 1.8 | DEFGH | 44.4 | BCDEF |
| PI286255 | Moneymaker | United Kingdom | Q1 | 1.9 | CDEFGH | 47.2 | ABCDEF |
| PI339940 | Chalks Early Jewel | United States | Q1 | 2.0 | BCDEFGH | 50.0 | ABCDEF |
| PI601136 | Baxters Early Bush Cherry | Texas, United States | Q1 | 2.0 | BCDEFGH | 50.0 | ABCDEF |
| PI645390 | CMVF 232 | Nevada, United States | Q1 | 2.0 | BCDEFGH | 41.7 | CDEFG |
| PI311109 | Tomate Jocotillo | Guatemala | Q1 | 2.1 | BCDEFGH | 44.4 | BCDEF |
| PI636205 | T039 | Oklahoma, United States | Q1 | 2.3 | ABCDEFGH | 58.3 | ABCDEF |
| PI644794 | Winsall | Illinois, United States | Q1 | 2.3 | ABCDEFGH | 58.3 | ABCDEF |
| PI645389 | H 2990 | Nevada, United States | Q1 | 2.3 | ABCDEFGH | 58.3 | ABCDEF |
| PI270234 | Loran Blood | United States | Q1 | 2.4 | ABCDEFGH | 50.0 | ABCDEF |
| PI270226 | Early Santa Clara Canner | United States | Q1 | 2.6 | ABCDEFGH | 63.9 | ABCDEF |
| PI639208 | Black from Tula | Tula, Russian Federation | Q1 | 2.8 | ABCDEFG | 61.1 | ABCDEF |
| PI547073 | NC 8276 | North Carolina, United States | Q2 | 2.9 | ABCDEF | 72.2 | ABCDE |
| PI109836 | Precoce des Halles | France | Q1 | 3.0 | ABCDEF | 66.7 | ABCDEF |
| PI600930 | Moran 3053 | United States | Q2 | 3.0 | ABCDEF | 75.0 | ABCD |
| PI205041 | P.A. Young T162 FS-1 | United States | Q1 | 3.1 | ABCDE | 69.4 | ABCDEF |
| PI601449 | Bealls Gourmet | United States | Q1 | 3.1 | ABCDE | 77.8 | ABCD |
| PI647513 | Red Pear | United States | Q1 | 3.1 | ABCDE | 77.8 | ABCD |
| PI601118 | VF 9209 | California, United States | Q2 | 3.2 | ABCDE | 61.1 | ABCDEF |
| PI279565 | Caro Red | United States | no data | 3.2 | ABCDE | 63.9 | ABCDEF |
| PI339914 | Coldset | Ontario, Canada | Q1 | 3.2 | ABCDE | 80.6 | ABC |
| PI254655 | Ker-1-M | United States | Q1 | 3.3 | ABCD | 63.9 | ABCDEF |
| PI270232 | Homestead | United States | Q1 | 3.3 | ABCD | 83.3 | ABC |
| PI647566 | Flora-dade | United States | Q2 | 3.3 | ABCD | 83.3 | ABC |
| PI647196 | Rutgers | United States | Q1 | 3.4 | ABC | 66.7 | ABCDEF |
| PI647445 | Zhongza No. 4 | China | Q2 | 3.4 | ABC | 86.1 | AB |
| PI601117 | Peelmech | California, United States | Q2 | 3.5 | AB | 69.4 | ABCDEF |
| PI279817 | Scotia | Canada | Q1 | 3.6 | AB | 72.2 | ABCDE |
| PI499370 | Prevoskhodnyi 176 | Former, Soviet Union | Q1 | 3.6 | AB | 77.8 | ABCD |
| PI601098 | Indiana 812 | Indiana, United States | Q2 | 3.6 | AB | 88.9 | A |
| PI645214 | Floradel | Florida, United States | Q2 | 3.6 | AB | 88.9 | A |
| PI645391 | Florida MH-1 | Michigan, United States | Q1 | 3.6 | AB | 88.9 | A |
| PI645398 | 466 Jungs Improved Wayahead | Wisconsin, United States | Q1 | 3.6 | AB | 80.6 | ABC |
| PI647523 | VFNT Cherry | United States | Q1 | 3.8 | A | 72.2 | ABCDE |
| BW | Source | DF | Sum of Squares | Mean Square | F Ratio | Prob > F | EMS | H2% |
|---|---|---|---|---|---|---|---|---|
| BW Score | Genotype | 39 | 92.584 | 2.374 | 2.493 | 0.0003 | σ2e + rσ2G | 59.9 |
| Error | 80 | 76.185 | 0.952 | σ2e | ||||
| C. Total | 119 | 168.769 | ||||||
| BW_DSI | Genotype | 39 | 47,138.31 | 1208.67 | 1.7507 | 0.0177 | σ2e + rσ2G | 42.8 |
| Error | 80 | 55,231.48 | 690.39 | σ2e | ||||
| C. Total | 119 | 102,369.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phiri, T.M.; Bhattarai, G.; Chiwina, K.E.; Fan, Q.; Xiong, H.; Alatawi, I.; Dickson, R.; Joshi, N.K.; Rojas, A.; Ling, K.-S.; et al. An Evaluation of Bacterial Wilt (Ralstonia solanacearum) Resistance in a Set of Tomato Germplasm from the United States Department of Agriculture. Agronomy 2024, 14, 350. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14020350
Phiri TM, Bhattarai G, Chiwina KE, Fan Q, Xiong H, Alatawi I, Dickson R, Joshi NK, Rojas A, Ling K-S, et al. An Evaluation of Bacterial Wilt (Ralstonia solanacearum) Resistance in a Set of Tomato Germplasm from the United States Department of Agriculture. Agronomy. 2024; 14(2):350. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14020350
Chicago/Turabian StylePhiri, Theresa Makawa, Gehendra Bhattarai, Kenani Edward Chiwina, Qiurong Fan, Haizheng Xiong, Ibtisam Alatawi, Ryan Dickson, Neelendra K. Joshi, Alejandro Rojas, Kai-Shu Ling, and et al. 2024. "An Evaluation of Bacterial Wilt (Ralstonia solanacearum) Resistance in a Set of Tomato Germplasm from the United States Department of Agriculture" Agronomy 14, no. 2: 350. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14020350