Extracellular Vesicles for Childhood Cancer Liquid Biopsy
by
 , , and
, , and
Nilubon Singhto
1,†, 
Pongpak Pongphitcha
2,3,†,
Natini Jinawath
4,5,6,
Suradej Hongeng
3 and
Somchai Chutipongtanate
7,8,* 1
Ramathibodi Comprehensive Cancer Center, Faculty of Medicine Ramathibodi Hospital, Mahidol University, Bangkok 10400, Thailand
2
Bangkok Child Health Center, Bangkok Hospital Headquarters, Bangkok 10130, Thailand
3
Division of Hematology and Oncology, Department of Pediatrics, Faculty of Medicine Ramathibodi Hospital, Mahidol University, Bangkok 10400, Thailand
4
Program in Translational Medicine, Faculty of Medicine Ramathibodi Hospital, Mahidol University, Bangkok 10400, Thailand
5
Chakri Naruebodindra Medical Institute, Faculty of Medicine Ramathibodi Hospital, Mahidol University, Samut Prakan 10540, Thailand
6
Integrative Computational Biosciences Center, Mahidol University, Nakon Pathom 73170, Thailand
7
MILCH and Novel Therapeutics Laboratory, Division of Epidemiology, Department of Environmental and Public Health Sciences, University of Cincinnati College of Medicine, Cincinnati, OH 45267, USA
8
Extracellular Vesicle Working Group, University of Cincinnati College of Medicine, Cincinnati, OH 45267, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cancers 2024, 16(9), 1681; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16091681
Submission received: 9 April 2024
/
Revised: 22 April 2024
/
Accepted: 24 April 2024
/
Published: 26 April 2024
(This article belongs to the Special Issue Advances in Multi-Omics of Pediatric, Adolescence and Young Adult Cancers)
Abstract
:Simple Summary
Liquid biopsy is a technique that uses minimally invasive or noninvasive methods to detect disease biomarkers in biofluids, such as blood and urine. For childhood cancers, this approach is promising, especially when tissue biopsies are challenging. By analyzing tiny particles called extracellular vesicles, the small particles in biofluids which carry genetic and molecular information from cancer cells, this liquid biopsy technique can aid early diagnosis, treatment monitoring, and outcome prediction, potentially improving the outcomes of childhood cancers. This review article introduces the concept of extracellular vesicle liquid biopsy, summarizes the progress that is being made in the diagnosis of multiple types of pediatric malignancies, and provides prospects for using this extracellular vesicle method to guide the development of novel cancer therapeutics.
Abstract
Liquid biopsy involves the utilization of minimally invasive or noninvasive techniques to detect biomarkers in biofluids for disease diagnosis, monitoring, or guiding treatments. This approach is promising for the early diagnosis of childhood cancer, especially for brain tumors, where tissue biopsies are more challenging and cause late detection. Extracellular vesicles offer several characteristics that make them ideal resources for childhood cancer liquid biopsy. Extracellular vesicles are nanosized particles, primarily secreted by all cell types into body fluids such as blood and urine, and contain molecular cargos, i.e., lipids, proteins, and nucleic acids of original cells. Notably, the lipid bilayer-enclosed structure of extracellular vesicles protects their cargos from enzymatic degradation in the extracellular milieu. Proteins and nucleic acids of extracellular vesicles represent genetic alterations and molecular profiles of childhood cancer, thus serving as promising resources for precision medicine in cancer diagnosis, treatment monitoring, and prognosis prediction. This review evaluates the recent progress of extracellular vesicles as a liquid biopsy platform for various types of childhood cancer, discusses the mechanistic roles of molecular cargos in carcinogenesis and metastasis, and provides perspectives on extracellular vesicle-guided therapeutic intervention. Extracellular vesicle-based liquid biopsy for childhood cancer may ultimately contribute to improving patient outcomes.
1. Introduction
The evolution of pathological measures from histomorphology and immunohistochemistry to molecular diagnostics has dramatically enhanced the accuracy of cancer diagnosis and classification. This advancement offers more precise prognostication and tailors therapeutic plans, including targeted therapy and immunotherapy, for individual cancer patients [1]. To obtain tumor tissues for diagnosis, invasive procedures are required, including surgical removal or biopsy. These procedures come with risks of anesthesia complications, bleeding, and infection [2]. Furthermore, the inherent heterogeneity of tumor cells and their clonal evolution over time often necessitate repeated biopsies from primary, metastatic, or relapsed tumors, adding additional risk to patients [3]. Tissue biopsies may also introduce sampling bias, leading to an incomplete analysis of tumor subpopulations. Moreover, tumors in some areas, such as the brain stem, are inaccessible for biopsy due to the unacceptable risk of mortality [4].
Liquid biopsy has emerged as a minimally invasive approach supporting diagnosis, making therapeutic decisions, and monitoring various types of cancers when surgical or biopsy procedures are unavailable or unacceptable [5]. This approach also holds potential for early screening and monitoring of minimal residual disease [6]. Liquid biopsy captures tumor fragments or circulating tumor contents from body fluids such as blood [7], cerebrospinal fluid [8], ascites [9], saliva [10], bronchoalveolar lavage [11], and urine [12]. This allows for repeated assessments during diagnosis, treatment, and follow-up periods with less invasiveness and minimized complications [13,14]. Therefore, liquid biopsy allows the possibility of detecting overall subclonal tumor heterogeneity throughout the clinical course [15].
Blood and plasma are reliable sources for most clinical investigations, and the same is true for cancer liquid biopsy [16]. For example, peripheral blood liquid biopsy-next-generation sequencing tests have been approved by the Food and Drug Administration (FDA) since 2020 to identify BRCA1/2, ALK, PIK3CA, and ATM pathogenic variants as part of the diagnosis of breast, ovarian, and lung cancers and treatment guidance (https://www.fda.gov/) [17]. Studies have demonstrated the potential applicability of blood-based liquid biopsies in childhood cancers [18], i.e., neuroblastoma, osteosarcoma, Ewing sarcoma, and hematologic malignancies [18]. Urine collection is generally less uncomfortable than taking blood and does not require trained medical staff. Urine liquid biopsies have shown promise in tracking somatic mutations in pediatric patients with Wilms tumors and other childhood renal tumors for diagnosis, risk stratification, and monitoring treatment response [19]. The blood–brain barrier blocks the transmigration of tumor tissue fragments, e.g., cell-free DNA (cfDNA), between systemic circulation and the central nervous system (CNS), thus making CSF a viable option for developing liquid biopsies for primary brain tumors and CNS metastases [20]. Various techniques and platforms have been developed for CSF liquid biopsy, for instance, detecting histone H3 mutation [21] and cfDNA [22,23] in CSF for diagnosis and monitoring of gliomas and medulloblastoma [20]. Ascites and pleural effusion can be good resources for liquid biopsies to determine treatment responses in advanced lung and gastrointestinal malignancies [24], while saliva is an invaluable source for head and neck cancers [25,26].
Cancer liquid biopsy platforms mainly rely on cell-related extracellular components, including circulating tumor cells (CTCs), circulating tumor DNA (ctDNA), and tumor-derived extracellular vesicles (EVs), each of which has unique advantages and challenges (Table 1).
CTCs are cancer cells that detach from the primary tumor and invade systemic circulation [27]. CTCs can appear as individual cells or clusters, with the latter being associated with a higher metastasis potential [28]. Various methods have been developed to detect CTCs [29], including immunocytochemical methods [30], fluorescence-activated cell sorting (FACS) [31], reverse transcription-quantitative polymerase chain reaction [32], fluorescence in situ hybridization (FISH) [33], and mass spectrometry [27]. Nonetheless, the main challenge of CTC liquid biopsy is based on the scarce number of CTCs in circulation, i.e., patients with advanced metastasis show up to 10 CTCs per 1 mL of peripheral blood [28]. In addition, CTC detection provides insights into cancer-related invasiveness and metastasis rather than information on genetic mutations, translocations, or rearrangements [27,34].
CtDNA is typically shorter and more fragmented than non-tumor DNA, representing 0.01–10% of total cell-free DNA in the bloodstream [34] and carrying various cancer-specific molecular markers, such as single-nucleotide mutations, methylation, and cancer-derived viral sequences [35]. A unique advantage of ctDNA is that it requires a smaller biofluid sample for assessing tumor mutational burden. Studies indicated that 1 mL of plasma is sufficient for ctDNA monitoring for tumor response to treatment [36,37], making it particularly useful for pediatric patients, where obtaining large sample volumes can be difficult. CtDNA-based liquid biopsy has been investigated in various types of pediatric solid tumors, e.g., neuroblastoma [38], Ewing sarcoma [39], osteosarcoma [40], medulloblastoma [14], renal tumors [41], rhabdomyosarcoma [42], and retinoblastoma [43]. However, ctDNA has a short half-life of two hours since it is degraded by extracellular DNase enzymes and rapidly cleared from systemic circulation by various organs, including the kidneys and liver [44]. The low abundance of cancer-specific ctDNA and the corresponding high abundance of background DNA in circulation also pose a considerable challenge in early tumor stage detection [45].
Recent studies increasingly point to EVs as a critical player in liquid biopsy diagnostics [46,47,48]. EVs have gained attention due to their unique characteristics of carrying various molecular cargos, i.e., lipids, proteins, and nucleic acids of origin cells [46,49]. Thus, EVs play a crucial role in cell-to-cell communication mediated through ligand–receptor interaction and membrane fusion to release molecular contents, e.g., growth factors, kinases, enzymes, and non-coding RNAs, all of which can elicit changes and reprogram intracellular activities of the recipient cells [50]. Moreover, EVs can reflect the altered physiological and pathological states of their parent cells and organs. This fact opens new possibilities for circulating EVs in cancer liquid biopsies [46,49]. Note that EVs presented in biofluids are heterogeneous, consisting of multiple subpopulations released from various cell sources overlapping particle size and density. Thus, a challenge in employing EVs for liquid biopsy lies in the complexities of EV isolation and characterization, necessitating meticulous optimization at the initial phase of any project. However, when compared with CTCs and ctDNA, EVs offer several advantages for liquid biopsies. First, EVs are abundant in biofluids, approximately 1 × 109 particles/mL, making it relatively easier to collect sufficient quantities for analysis [51]. Second, EVs are actively secreted by living cells, including tumor cells, and carry a diverse range of information reflective of ongoing activities within their parent cells [51]. In contrast, ctDNA primarily releases from apoptotic or dead tumor cells and may provide less accurate information [51]. Finally, the lipid bilayer-enclosed EV structure provides protection to molecular cargos from proteases, DNases, and RNases in biofluids, allowing them to circulate in the bloodstream from 30 min to 6 h without degradation [49,52].
In this review, according to the promising characteristics of EVs for liquid biopsy, we summarize the EV definition and essential methods for isolation and characterization and then extensively explored the research involving the potential of EVs as biomarkers for diagnosis, treatment monitoring, and prognostic prediction in pediatric malignancies. Additionally, we discusse the mechanistic roles of EVs in carcinogenesis and metastasis, and provide our perspectives on future directions of EV-based liquid biopsy for childhood cancer.

{kind=link}
{kind=link}
Table 1.
Tissue fragments detectable by cancer liquid biopsy.
| Tissue Fragment | Description | Analytical Techniques | Advantages | Limitations | References |
|---|---|---|---|---|---|
| Circulating tumor cells (CTCs) | Cancer cells detach from the primary tumor or metastatic lesion and circulate in the bloodstream. |
|
|
| [27,28,29,31,32,33] |
| Circulating tumor DNA (ctDNA) | ctDNA is shed by primary tumor cells and subsequently enters the bloodstream. As such, ctDNA carries the genetic alterations present in the original tumor. |
|
|
| [35,36,37,41,45,53,54,55] |
| Extracellular vesicles (EVs) | Tiny membranous structures enclosed by a lipid bilayer are secreted by both normal and cancerous cells. EVs contain a diverse assortment of molecular constituents, including DNA, RNA, non-coding RNA, and proteins. |
|
|
| [51,56,57,58] |
2. EV Definition and Methods: A Brief Summary
The International Society for Extracellular Vesicles (ISEV) has recently launched minimal information for studies of extracellular vesicles 2023 (MISEV2023). In these new guidelines, the definition of EVs has been updated as “the particles that are released from cells, are delimited by a lipid bilayer, and cannot replicate on their own” [58]. Since it is likely that a broad population of EVs is being studied rather than a solitary subtype, biogenesis-related terms, including exosomes [59,60] and ectosomes [61,62], should not be used unless their subcellular origins can be demonstrated in their studies [58]. Ambiguous terms, i.e., exosome-like vesicles and microvesicles, are discouraged to avoid any nomenclature confusion in the field [58]. To address EV subtypes, operational terms are recommended based on the diameter of the separated particles: small EVs (sEVs; <200 nm) and large EVs (LEVs; >200 nm), where cautions should be made since the measured diameter is related to the specific characterization method [58]. In this review, we follow the MISEV2023 guideline and only use one umbrella term, EVs, when referring to any types of vesicles investigated in the original studies.
For EV isolation, the single-step procedure, i.e., high-speed ultracentrifugation (UC), polymer/precipitation agents (e.g., polyethylene glycol), and ultrafiltration (UF), offers high EV recovery from biofluids [63,64]. However, these techniques may lead to reduced specificity due to the contaminations of non-vesicular components (e.g., soluble proteins and lipoprotein micelles). In contrast, size-exclusion chromatography (SEC), asymmetrical flow field-flow fractionation, and immunoaffinity bead capture offer better specificity while ensuring reasonable recovery yields [63]. Nonetheless, when superior EV specificity is paramount, especially in studies focused on specific EV functions or subpopulations, a multi-step combinatorial method is recommended [65], for example, two or three-step combinations between a form of centrifugation, polymer precipitation, UF, and SEC [64,65,66,67]. These multi-step protocols ensure minimal contamination, yielding samples optimal for direct downstream analysis and data interpretation. However, their scalability and reproducibility must be addressed in the early phase of the project to ensure sufficient recovery yields with the high specificity of EVs for further characterizations.
After EV isolation, various techniques are used to characterize the purity of EVs, commonly based on morphology, hydrodynamic size, and protein markers [68]. Transmission electron microscopy (TEM) provides a high-resolution image of the nano-size EVs morphology [56]. However, morphology imaging may be affected by swelling, shrinking, or flattening from sample preparation steps, e.g., fixation, dehydration, embedding, and imaging conditions [56]. To overcome the above limitation, atomic force microscopy (AFM) and cryo-electron microscopy provide detailed information and solve the variable effects of TEM, but these techniques tend to be relatively low-throughput and resource-intensive [56]. In addition, EVs can be determined by depending on biophysical approaches. Several techniques, including nanoparticle tracking analysis (NTA), resistive pulse techniques, and dynamic light scattering, are used for determining the distribution of EV size [56]. However, these techniques have been reported to be sensitive to the size of protein or glycan extending from the EV membrane, and they do not provide the phenotype of vesicles. Protein evidence of EV presence in the isolates is usually demonstrated by three positive EV markers and one negative marker, including (i) surface proteins and CD9, CD63, and CD81 tetraspanins; (ii) proteins involved in EV biogenesis (such as the Tsg101, Alix, and Rab families); (iii) cytosolic proteins (e.g., Hsp70 and Gapdh); (iv) absence of proteins representing intracellular organelles (i.e., calnexin, GM130, and histone).
3. EV-Based Liquid Biopsy for Childhood Cancers
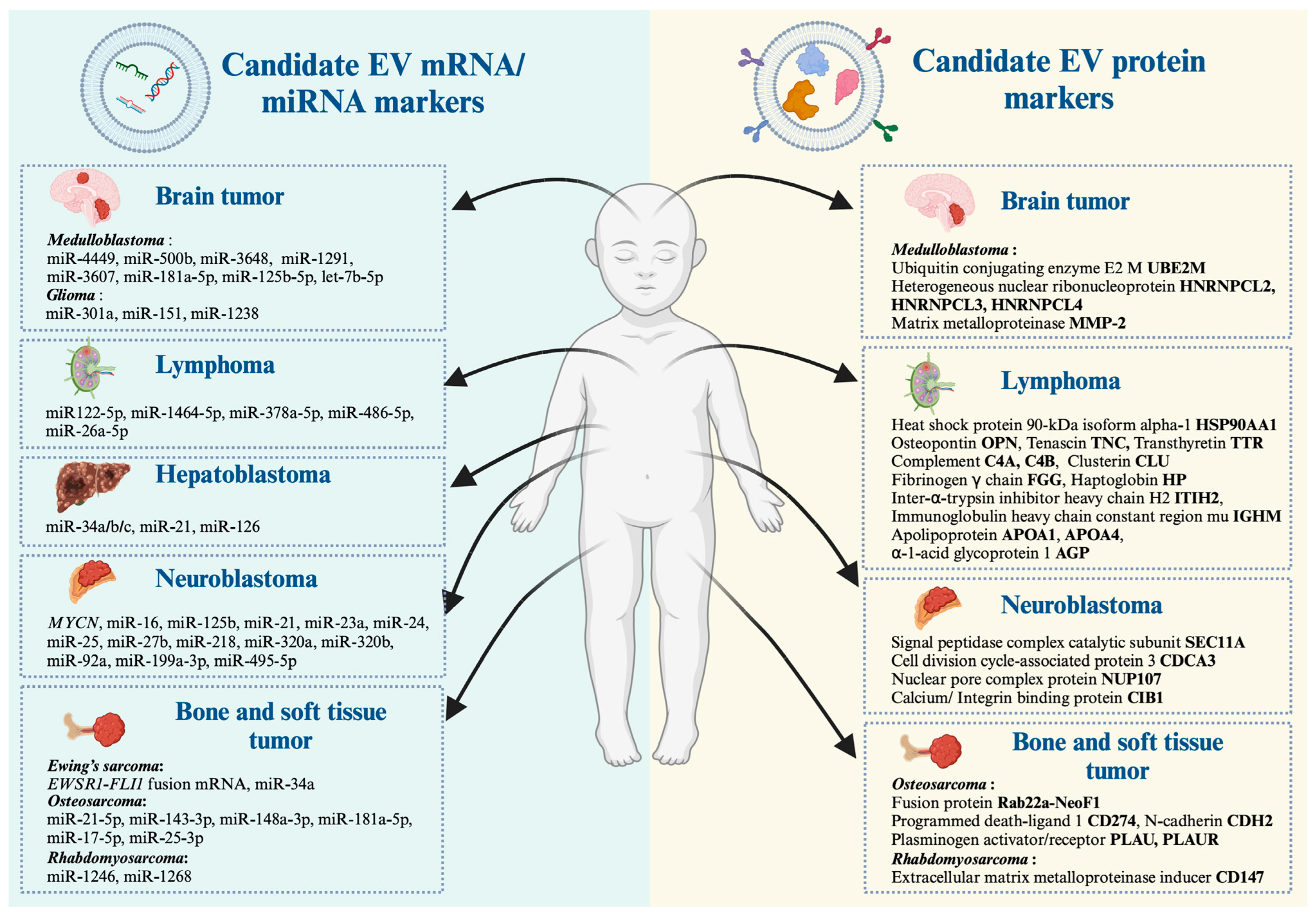
In the context of cancer, EVs play a role in facilitating metastasis by promoting the formation of a pre-metastatic niche (PMN) [69], which is the microenvironment at a distant organ that disseminates tumor cells from the primary tumor colonized and grows into metastasis [69]. Tumor-derived EVs prepare the PMN by modulating extracellular matrix formation, immune suppression, angiogenesis, and facilitating the recruitment of other stromal cells to support tumor growth and survival [69]. These EV molecular cargos serve as invaluable resource to discover biomarker candidates for several types of childhood cancer, including neuroblastoma, medulloblastoma, hepatoblastoma, osteosarcoma, Ewing’s sarcoma, rhabdomyosarcoma, and lymphoma. In the following section, we provide current evidence of EV studies conducted over recent decades with a special emphasis on transcripts and protein candidates for the development of childhood cancer liquid biopsy (summarized in Figure 1 and Table 2).
3.1. Pediatric Neuroblastoma
Neuroblastoma is the most common extracranial pediatric solid tumor, which arises from neural crest cells [99]. The clinical spectrum of neuroblastoma can vary from localized disease and spontaneous regression to highly malignant and extensive metastasis disease [99]. Genetic aberrations include recurrent segmental chromosome aberration (losses of chromosome 1p, 3p, 4p, 11q and gains of 1q, 2p, 17q), amplification of the proto-oncogene MYCN, and epigenetic modifications [99]. Although there have been significant improvements in the prognosis of neuroblastoma patients, up to 40% of high-risk cases continue to pose a substantial challenge, with a considerable portion experiencing tumor relapse driven by minimal residue disease (MRD) [100].
Ongoing research fields put effort into improving the management of high-risk neuroblastoma through early detection of MRD by liquid biopsy [100]. Interestingly, cancer-specific modification DNA and RNA can be detected in EVs. Therefore, it is theoretically presumed that MRD could be detected through cancer EVs [101]. The extensive studies of molecular cargos of EVs derived from neuroblastoma in different disease states should be more elucidated.
For example, our group recently reported that transcripts of the MYCN gene, the strongest molecular risk of neuroblastoma, were detectable in LEVs isolated from patient-derived bone marrow plasma, where the presence and absence of MYCN-LEVs were associated with MYCN-amplification status and treatment–relapse states [46]. Haug et al. [70] reported that neuroblastoma cell lines with MYCN amplification release EVs containing oncogenic miRNAs, including 11 types of miRNAs (e.g., miR-16, 125b, 21, 23a, 24, 25, 27b, 218, 320a, 320b, and 92a) [70], where miR-92a was the highest miRNA expression [70]. This study implied that these EVs might contribute to the aggressive behaviors of MYCN-amplified neuroblastoma [70]. Similarly, Ma et al. [71] performed next-generation sequencing on miRNAs containing EVs isolated from human plasma. This study revealed that high numbers of miRNA were changed by either upregulated or downregulated in patients with neuroblastoma and ganglioneuroblastoma compared to healthy donors [71]. Among these altered miRNAs, two, including miR-199a-3p and miR-495-5p, overlap in neuroblastoma and ganglioneuroblastoma patients [71]. After validation by RT-qPCR, they discovered that only miR-199a-3p expression was significantly higher expressed in neuroblastoma patients [71]. Interestingly, miR-199-3p represents an independent risk factor since its expression in the plasma was more elevated in neuroblastoma patients with unfavorable histology than in those with a good prognosis [71].
Using proteomic analysis, Coletti et al. [72] compared EVs derived from neuroblastoma cell lines representing primary tumors and bone marrow metastasis. Overall, EV proteins of primary tumors are involved in neuronal development and function, suggesting a connection between the neuronal characteristics of neuroblastoma and EV cargo. In contrast, proteins exclusively present in EVs derived from neuroblastoma–bone marrow metastatic models, e.g., signal peptidase complex catalytic subunit SEC11, cell division cycle-associated protein 3, nuclear pore complex protein Nup107, calcium, and integrin-binding protein 1, are associated with cell survival, proliferation, and progression [72]. These proteins could be further developed as potential biomarkers of neuroblastoma tumor staging. Collectively, these studies suggested that EV-based liquid biopsy is useful for diagnosis, monitoring of treatment response/relapsed disease, and prognosis prediction in pediatric neuroblastoma [46,70,71,72].
3.2. Medulloblastoma
Medulloblastoma is the predominant malignant brain tumor in children, accounting for approximately 20% of all pediatric brain tumors. It is classified as an embryonal neuroepithelial tumor of the cerebellum [102]. According to the fifth edition of the WHO classification of tumor of the CNS, medulloblastoma incorporates both molecular subgroups and traditional variants to provide a more comprehensive understanding of the disease biology [103], including the wingless (WNT)-activated subgroup, sonic hedgehog (SHH)-activated subgroup, Group 3, and Group 4 [103]. In addition, the SHH-activated subgroup can be subdivided by the detection of TP53 gene mutation, which includes SHH-activated, TP53 wild-type, and SHH-activated TP53 mutants [103]. Nonetheless, the heterogeneity of the clinical course and outcome in each patient poses challenges regarding the lack of early diagnostic biomarkers for high-risk patients with recurrent and metastatic disease.
Kaid et al. [73] performed proteomic and miRNA analysis and revealed that 464 proteins and 10 microRNAs were exclusively detected in LEVs released from highly aggressive medulloblastoma cell lines. The interactome analysis of distinct proteins and miRNA suggested that ERK, PI3K/AKT/mTOR, EGF/EFGR, and stem cell self-renewal are the main oncogenic signaling pathways altered in aggressive medulloblastoma [73]. Four proteins (UBE2M, HNRNPCL2, HNRNPCL3, and HNRNPCL4) and five miRNAs (miR-4449, 500b, 3648, 1291, and 3607) were identified as the candidate biomarkers of aggressive medulloblastoma with potential applications in diagnosis, patient stratification, and early detection of relapsed disease [73]. In this direction, Zhu et al. [74] reported that increased levels of EV-derived miR-181a-5p, miR-125b-5p, and let-7b-5p were associated with in vitro invasion and migratory abilities of SHH medulloblastoma cells through the activation of ERK in the Ras/MAPK pathway [74].
Furthermore, a recent study by Jackson et al. [75] demonstrated that metastatic medulloblastoma cells released a higher amount of EVs compared to non-metastatic cells, and metastatic EVs significantly increased surface matrix metalloproteinase-2 (MMP-2). Importantly, this study found a high level of MMP-2 activity in CSF from three of four patients associated with tumor progression [75], suggesting that MMP-2-expressed EVs may modulate the PMN to drive medulloblastoma metastasis [75].
3.3. Pediatric Gliomas
Gliomas are the most common histologic type of brain tumor, can arise in all areas of the brain hemisphere, and are rarely seen at the spinal cord level [104]. Regarding the WHO Classification for CNS neoplasm, a comprehensive molecularly refined diagnosis is crucial for therapeutic plans and prognoses [104]. However, tissue biopsies in some areas, such as deep hemispheric lesions, optic pathways, brain stem, and spinal cord, are limited due to unacceptable risk for morbidity and mortality [104]. In this regard, liquid biopsy can make a significant contribution to glioma molecular diagnosis despite brain tumors in difficult-to-access areas [104]. Molecular alteration targets, such as H3, BRAF, EGFR, and IDH1 mutations, and other markers that could be used for diagnosis, monitoring treatment response, or developing novel therapeutics, have been investigated [104].
H3 mutations in diffuse high-grade gliomas are important for diagnosis and prognosis, while BRAF alterations in low-grade gliomas are valuable in both diagnosis and treatment. García-Romero et al. [105] demonstrated the feasibility of the detection of BRAF V600E targetable mutation by digital-PCR from cell-free-DNA and EV-derived DNA in serum, plasma, and CSF isolated from a cohort of 29 CNS pediatric patients. Chen et al. [76] used a novel approach that combines biofluid EV-RNA and BEAMing RT-PCR to detect and quantify mutant and wild-type IDH1 RNA transcripts in the CSF of patients with gliomas.
Several studies suggested that EV-derived miRNA could be the biomarker for patient classification, predict response to treatment, or serve as a novel therapeutic target for pediatric gliomas. Xiao et al. [77] detected the EV-derived miR-301a from glioblastoma multiforme (GBM) cells under hypoxic conditions that promoted radiation resistance in GBM. MiR-301a is involved in regulating the Wnt/β-catenin pathway and targeting anti-oncogene TCEAL7. Therefore, these findings might facilitate the search for a novel target to overcome radiotherapy resistance in GBM patients. Ailiang et al. [78] demonstrated that a lower expression of miR-151a was related to temozolomide (TMZ) resistance in GBM; the restoration of miR-151a expression sensitized TMZ-resistant GBM cells by inhibiting XRCC4-mediated DNA repair. The detection of EV-derived miR-151a might predict chemotherapy response but also represents a promising therapeutic target for therapy-refractory GBMs. Additionally, Jianxing et al. [79] revealed that overexpression of miR-1238 led to the required resistance against TMZ in GBM patients, while another EV-derived miRNA, miR-148a, was associated with a risk of GBM progression according to The Cancer Genome Atlas [106].
3.4. Hepatoblastoma
Hepatoblastoma is a rare and aggressive type of liver cancer that primarily affects infants and children younger than three years old [107]. This malignancy is reported to arise from abnormal developments in the liver during embryonic stages [107]. The pathogenesis of hepatoblastoma involves genetic and molecular alterations that disrupt the normal processes of liver organogenesis, leading to the uncontrolled growth of cells and the formation of tumors [107]. Some genetic syndromes have been associated with an increased predisposition to hepatoblastoma, which includes Beckwith–Wiedemann syndrome, hemihypertrophy, and familial adenomatous polyposis [107]. The early diagnosis of hepatoblastoma in patients with a genetic predisposition to the disease is crucial for improving treatment outcomes, allowing for prolonged intervention and potentially more effective treatment strategies.
In this direction, Jiao et al. [80] conducted a transcriptomic study to identify EV-derived miRNAs as potential biomarkers for hepatoblastoma. As a result, EV-derived miR-34a/b/c was significantly lower in the serum of patients with hepatoblastoma compared to healthy control groups, where the downregulation of miR-34 was associated with tumorigenesis in the lungs, skin, breast, urinary bladder, and kidneys [108]. In hepatoblastoma, decreased levels of EV-derived miR-34a/b/c might be considered as the diagnostic and prognostic biomarker. Likewise, Liu et al. [81] revealed that the elevated level of miR-21 containing EV in hepatoblastoma patients might be another biomarker for hepatoblastoma, while Hu et al. [82] demonstrated EV-derived miR-126 was upregulated in hepatoblastoma cells. These findings suggest that EV-derived microRNA may be involved in liver cancer tumorigenesis.
3.5. Osteosarcoma
Osteosarcoma is the most common primary bone tumor affecting adolescents and young adults. It is characterized by a heterogenous malignant spindle cell tumor and the formation of immature osteoid tissue or osteoid and accounts for 1% of all cancers [109]. The current treatment approach for osteosarcoma involves a combination of neoadjuvant chemotherapy, surgical removal of the tumor, and adjuvant chemotherapy [110]. Despite intensive treatment regimens, the 5-year survival rate for patients with metastatic and recurrent osteosarcoma is less than 20 percent [111]. Therefore, osteosarcoma research is mainly focused on identifying new therapeutic targets to improve outcomes and early detection of metastasis.
Jarez et al. [87] investigated EV-miRNA expression in osteosarcoma cell lines with varying degrees of metastatic potential. The results showed four miRNAs (miR-21-5p, 143-3p, 148a-3p, and 181a-5p) were highly expressed in EVs derived from metastatic osteosarcoma cells compared to non-metastatic cells [87]. Gene ontology analysis of predicted miRNA target genes suggested that EVs may regulate the metastatic potential of osteosarcoma by inhibiting several genes (e.g., MAPK1, NRAS, FRS2, PRCKE, BCL2, and QKI) involved in apoptosis and cell adhesion [87]. In this direction, Araki et al. [86] revealed that sEVs derived from malignant human osteosarcomas contained miR-146a-5p and could inhibit osteoblastogenesis. Ucci et al. [112] revealed that osteosarcoma EVs had increased pro-osteoclastogenic proteins, inflammatory cytokines, and metalloproteinase, with decreased proteins involved in the cell cycle and pro-osteoblastogenesis. Moreover, Zhong et al. [89] reported that osteosarcoma with the RAB22a-NeoF1 fusion gene produced EVs containing the Rab22a-NeoF1 protein to promote PMN formation and M2 macrophage induction, eventually enhancing lung metastases [89]. Furthermore, Endo-Munoz et al. [90] demonstrated that elevation of the urokinase plasminogen activator (uPA) and the uPA receptor (uPAR) in both soluble form and osteosarcoma-secreted EVs were exclusive in metastatic osteosarcoma cells, where the uPA inhibitor significantly mitigated osteosarcoma metastasis in an orthotopic mouse model [90]. These findings suggested that osteosarcoma EVs contain molecular cargos with abilities to modulate the PMN, which contributes to osteosarcoma progression [86,89,90,112], thus serving as the basis for developing osteosarcoma liquid biopsy in future EV studies.
3.6. Ewing’s Sarcoma
Ewing’s sarcoma is the second most common primary bone malignancy affecting children and young adults between 10 and 20 years old. It typically arises in long bones and the pelvis and sees around 200 new cases annually in the United States [113]. The underlying driven pathogeneses are characterized by specific rearrangements of one of five alternative ETS family member genes, i.e., FLI1 (90%), ERG (5–10%), ETV1 (<1%), E1AF (<1%), and FEV1 (<1%) fusion with EWSR1 [113]. Approximately 60–70% of patients with the localized disease have 3–5 years overall survival, while for those with metastasis or recurrence disease, the survival remains dismal due to limited therapeutic options [113]. Aside from clinical symptomatology and radiographic imaging, no suitable tumor markers or biomarkers exist to identify and monitor patients with a high risk of recurrence, resistance to standard treatment, or early disease progression [113]. Unique pathognomonic fusions or their related transcriptome may be the detection targets that are most likely to be detected.
Ewing’s sarcoma-derived EVs were first investigated by Miller et al. [83] using RT-qPCR to detect ES-specific transcripts such as EWSR1-FLI1. This discovery opened the possibility of exploring the role of EV-based liquid biopsy in Ewing’s sarcoma diagnosis and monitoring of the MRD. Then, Tsugita et al. [84] discovered that EWS/FLI1 fusion mRNA, resulting from t(11;22)(q24;q12) translocation, could be identified in LEVs secreted from Ewing sarcoma cells using RT-qPCR. These fusion mRNA-LEVs were also detectable in plasma from xenografted mice [84]. This finding suggested that the EWS/FLI1 mRNA containing LEVs could be further developed as a non-invasive liquid biopsy for precise diagnosis of Ewing’s sarcoma.
3.7. Rhabdomyosarcoma
Rhabdomyosarcoma is the most common soft tissue sarcoma, accounting for 5–10% of sarcomas in children, adolescents, and young adults [114]. Rhabdomyosarcoma can be divided based on histology into two main types: alveolar (25%) and embryonal (75%) [114]. Alveolar rhabdomyosarcoma is characterized by chromosomal translocation t(2;13)(q35;q14), which results in a fusion between the PAX3 gene on chromosome 2 and the FOXO1 gene on chromosome 13, or in some cases, t(1;13)(p36;q14), which leads to fusion between the PAX7 gene on chromosome 1 and the FOXO gene [114]. Embryo rhabdomyosarcoma comprises cells resembling immature skeletal muscle and tends to have a more favorable prognosis than alveolar rhabdomyosarcoma [114]. The genetic abnormalities associated with embryonal rhabdomyosarcoma involve muscle development and differentiation [114].
Gayad et al. [91] conducted miRNA profiling to compare EV-derived miRNA from embryonal vs. alveolar rhabdosarcoma cell lines. The findings suggested that miRNAs in alveolar rhabdosarcoma-derived EVs expressed a distinct set of 62 miRNAs, while embryonal rhabdomyosarcoma-derived EVs were enriched with another set of 34 miRNAs [91]. Only two miRNAs (miR-1246 and 1268) were universally presented in EVs of all cell lines examined [91]. Rammal et al. [93] performed proteomic analysis between alveolar rhabdomyosarcoma-derived EVs and those of embryonal type. As a result, 81 proteins were commonly presented in EVs from both subtypes, which involved cell signaling, cell movement, and cancer progression through the integrin signaling pathway, chemokine and cytokine signaling pathway, and angiogenesis [93]. Another study reported that CD147 was exclusively expressed in metastatic tumors of human rhabdomyosarcoma tissue, which contributed to tumor cell aggressiveness, and was involved in modulating the microenvironment through rhabdomyosarcoma-derived EVs [92]. Targeted inhibition of CD147 reduced its expression in isolated EVs and suppressed their capability to enhance rhabdomyosarcoma cell invasive properties [92].
3.8. Pediatric Lymphoma
Lymphoma is a hematologic malignancy affecting the lymphatic system, which occurs in approximately 15% of childhood cancer [115]. According to the WHO system, lymphoma is classified as Hodgkin (HL) and non-Hodgkin lymphoma (NHL) [115]. HL is characterized by Reed–Sternberg cells and specific abnormal B lymphocytes and is primarily found in early and late adulthood but rarely in children younger than five years of age [115]. NHL originates from either abnormal B or T cells, accounts for approximately 5% of childhood cancers, and is more likely to present in younger children than HL [115]. NHL can be classified into various subtypes based on genetic markers, including Burkitt lymphoma, diffuse large B-cell lymphoma, primary mediastinal large B-cell, anaplastic large-cell lymphoma, and lymphoblastic lymphoma [116]. Pediatric lymphoma treatment typically includes the combination of chemotherapy and radiation therapy and, in some cases, may require stem cell transplantation. Early diagnosis and appropriate treatment are critical for successfully managing pediatric lymphomas.
Damanti et al. [94] investigated the role of EV-miRNAs as disease biomarkers and their functions in pediatric anaplastic large cell lymphoma (ALCL) compared to healthy donors [94]. They found miR-122-5p, a critical player in tumor cell dissemination and aggressiveness, was elevated in plasma EVs derived from ALCL patients. Of note, miR-122-5p may be a therapeutic target for ALCL pediatric patients [94]. Likewise, small RNA-sequencing analysis in plasma EVs isolated from 20 children with pediatric ALCL showed that miR-146a-5p might be useful for prognosis prediction in high-risk patients [95] since miR-146a-5p plays a role in promoting macrophage infiltration and M2-like polarization, thereby promoting ALCL aggressiveness and dissemination [95].
Lovisa et al. [97] performed a proteomics study to compare plasma EV derived from pediatric ALCL patients and healthy donors and identified up to 50 proteins that were significant in EVs derived from ALCL patients [97]. Functional enrichment analysis revealed that these proteins are related to cell adhesion, glycosaminoglycan metabolic process, extracellular matrix organization, collagen fibril organization, and acute phase response, as well as the PI3K/AKT pathway [97]. This study suggested that these ALCL-derived EV proteins can be used as diagnostic and possibly prognostic parameters during diagnosis and ALCL disease monitoring [97]. Another proteomic study identified 11 unique proteins, including five upregulated in non-relapsed HL (e.g., isoform 2 preproprotein of complement C4-A, complement C4-B, fibrinogen γ chain, inter-α-trypsin inhibitor heavy chain H2, and immunoglobulin heavy chain constant region mu) and six upregulated in relapsed HL (e.g., apolipoprotein A-I, apolipoprotein A-IV, clusterin, haptoglobin, α-1-acid glycoprotein 1, and transthyretin) [98]. Taken together, this unique set of EV proteins might be used as the biomarker to discriminate the non-relapse and relapse condition of HL patients [98].
4. Future Prospects
A growing number of research studies have demonstrated that EVs relate to tumorigenesis and mediate intercellular communication, which eventually leads to progression and metastasis (as summarized in Figure 2 and Table 3). EVs have been implicated in the pathogenesis and dissemination of several malignancies, as they facilitate the reciprocal communication between malignant cells and their surrounding microenvironment, hence contributing to both the advancement of the illness and the formation of metastases [117]. From this point of view, EV-based liquid biopsy with deep molecular characterization may offer mechanistic insights into tumorigenesis and progression, which will facilitate the development of personalized and precise treatment strategies.
For instance, a preclinical study unraveled a new mechanism of dinutuximab resistance in high-risk neuroblastoma: sEV-mediated inhibition of NK cell infiltration and tumor-associated macrophage (TAM) recruitment [118]. Neuroblastoma-derived sEVs suppressed splenic NK cell maturation in vivo and inhibited antibody-dependent cellular cytotoxicity mediated by dinutuximab-induced NK cells in vitro [118]. Here, RNA-sequencing of neuroblastoma sEVs identified molecular alterations involving immune effector cells, i.e., myeloid, dendritic, and NK cells, and predicted a negative regulation of leukocyte activation, thereby contributing a groundwork for neuroblastoma EV liquid biopsy to detect the immunotherapeutic resistant phenotype. Chromosomal 17q21-ter is commonly gained in neuroblastoma, where the insulin-like growth factor-2 mRNA-binding protein 1 (IGF2BP1) gene is located (17q21.32) [129]. IGF2BP1 upregulation in neuroblastoma was associated with lower overall survival and positively correlated with MYCN mRNA, even in patients with MYCN-non-amplified tumors [129]. In vivo, Dhamdhere et al. [126] demonstrated the neuroblastoma EV-mediated pro-metastatic effect of IGF2BP1, whereas EVs from IGF2BP1 knockdown NB cells did not trigger the metastatic process. Proteomic analysis revealed that two IGF2BP1-regulated proteins, SHMT2 and SEMA3A, were increased in metastatic neuroblastoma EVs. A validation study revealed the increased plasma EV levels of SHMT2 and SEMA3A proteins in patient-derived xenograft-bearing mice [126], supporting the clinical significance of IGF2BP1/SEMA3A-SHMT2 axis in neuroblastoma metastasis, which warrants further clinical studies of neuroblastoma liquid biopsy to inform therapeutic regimens against potential metastasis. Mancarella et al. [127] reported that Ewing’s sarcoma secreted EVs loaded with insulin-like growth factor 2 mRNA binding protein 3 (IGF2BP3), which promoted migration but not the proliferation of the recipient cells [127]. EV-miRNA profiling revealed that IGF2BP3 silencing of Ewing’s sarcoma cells altered specific miRNAs associated with the PI3K/Akt pathway in recipient cells [127], including miR-223-3p which target IGF1R, a major driver of EWS aggressiveness and a previously reported target of IGF2BP3. This finding indicated that IGF2BP3 containing EVs may have a role in regulating phenotypic heterogeneity and promoting cell migration of Ewing’s sarcoma [127].
For osteosarcoma, Macklin et al. [128] demonstrated that EVs released by cells originating from a clonal variant of the osteosarcoma cell line with high metastatic potential can be taken up by a clonal variant of the same cell line with low metastatic ability [128]. This uptake of EVs leads to the induction of a migratory and invasive phenotype [128]. The observed transfer of phenotypic traits in a horizontal manner is unidirectional, suggesting that the ability to metastasize may be developed through interclonal communication [128]. Here, proteomic analysis of EVs released by osteosarcoma clones with high metastatic potential identified several proteins relating to G-protein coupled receptor signaling pathways [128]. These findings support further investigation of EV liquid biopsy to guide precise treatment in osteosarcoma patients.
The natural ability of EVs to transport diverse bioactive compounds demonstrates their potential utility as a drug delivery system. One remarkable advantage is using EVs as biocompatible carriers for gene therapy and RNA therapeutics. Genetic engineering techniques may facilitate the encapsulation of tumor suppressor proteins into EVs. Investigating these approaches may significantly advance therapeutic strategies for pediatric cancers.
Few studies explored EV liquid biopsy and translated their findings to develop therapeutic approaches for pediatric cancers. For instance, Xue et al. [130] discovered miR-101-3p and miR-423-5p by comparing plasma EVs derived from medulloblastoma patients and healthy individuals. They found that miR-101-3p and miR-423-5p function as tumor suppressors through the direct regulation of a common gene, FOXP4, which is responsible for encoding a transcription factor that plays a crucial role in embryonic development and tumorigenesis [130]. Additionally, miR-101-3p exhibited targeting capabilities towards EZH2 (histone methyltransferase), enhancing its suppressive impact on tumor growth [130]. The use of miR-101-3p and miR-423-5p mimics significantly suppressed the proliferation, colony-forming ability, migratory ability, and invasive capability of medulloblastoma cells [130]. In addition, a xenograft nude mouse model of medulloblastoma was used to investigate the effects of overexpression miR-101-3p and miR-423-5p on tumorigenesis, which suppressed the progression of tumors [130]. In addition, Huang et al. found an elevated level of miR-130b-3p in plasma EVs obtained from medulloblastoma patients compared to healthy controls [131]. Interestingly, the miR-130b-3p mimic inhibited medulloblastoma cell proliferation by targeting serine/threonine-protein kinase 1 (SIK1), promoting the activity of the p53 signaling pathway, thus serving as a tumor suppressor miRNA in medulloblastoma [131]. These findings highlight a possibility that EV liquid biopsy may capture self-defense responses against tumorigenesis, thereby opening a new venue to investigate novel miRNA therapeutic approaches for pediatric cancers.
It should be emphasized that EVs have several advantages compared to other liquid biopsy platforms for pediatric cancer. For instance, EVs contain a variety of biomolecules, reflect tumor heterogeneity, preserve stability, and reflect the real-time change in intracellular signaling pathways that lead to identifying therapeutic targets. It is anticipated that EV liquid biopsy will make a significant contribution as a diagnostic platform for pediatric solid tumors. Nonetheless, prospective studies and validations of specific markers are required for clinical applications. To enable clinical integration of non-invasive EV liquid biopsies, especially for brain tumors where EV liquid biopsies offer transformative potential, it is essential to benchmark their diagnostic performance against established gold standard methods.
5. Conclusions
This review summarizes the recent progress of EV liquid biopsies in pediatric cancers and provides perspectives to explore EV biological roles, which can lead to the development of novel therapeutic approaches. The cumulative evidence suggests that EV cargo is less susceptible to degradation due to the protective role of lipid membranes surrounding vesicles. As a result, EV cargo is more stable than ctDNA and soluble proteins in bodily fluids [51,132,133]. Due to their abundance and ability to protect cargo, EVs have demonstrated useful detection sensitivities and have often been found to be superior to techniques employing ctDNA or soluble proteins as tumor antigens [51,132,133]. EV-based liquid biopsies have demonstrated promising outcomes in clinical trials, specifically for the early stage detection of cancer and the monitoring of minimal-residual disease in post-therapy follow-up patients [101,133]. Furthermore, EVs offer a similar benefit to CTCs in that they can serve as a versatile platform for detecting and measuring various types of cancer-related biomolecules. CTCs are limited by their small level in the blood, while EVs are plentiful in all human bodily fluids and offer a wide range of diagnostic possibilities [28,133]. Therefore, the analysis of EV cargo has become a promising tool for prognosis/diagnosis and is now attracting significant interest. However, there is no “gold standard” for EV isolation or characterization protocols. EV nomenclature remains inconclusive. Additionally, the challenges also affect pre-analytical procedures, including storage duration, number of freeze-and-thaw cycles and factors during transportation, which should be considered as sources of technical variations. In addition, the majority of clinical investigations of EV studies have been conducted with a relatively small cohort of patients, which lack the statistical strength of studies and the accuracy of corresponding findings [133]. In order to address the challenge mentioned earlier, the International Society for Extracellular Vesicles (ISEV) has developed the MISEV guidelines. These guidelines aim to establish a common framework for conducting studies on EVs using different protocols [57,58]. By accomplishing this, the guidelines can help minimize variations in EV detection and enhance their potential for clinical diagnosis. Prospective validation studies are needed to ensure the accuracy and reliability of EV liquid biopsies for precise diagnosis, monitoring disease progression, and guiding therapeutic intervention, which ultimately improves patient outcomes.
Author Contributions
Conceptualization, S.C.; methodology, N.S., P.P. and S.C.; resources, N.J., S.H. and S.C.; data curation, N.S. and P.P.; writing—original draft preparation, N.S. and P.P.; writing—review and editing, N.J., S.H. and S.C.; supervision, N.J., S.H. and S.C.; funding acquisition, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was partially supported by the Genomic Thailand Project of the Health Systems Research Institute, grant number HSRI64-130 (to S.H.), and Ramathibodi Foundation (to S.H.).
Acknowledgments
All figures were created with Biorender.com.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Malone, E.R.; Oliva, M.; Sabatini, P.J.B.; Stockley, T.L.; Siu, L.L. Molecular profiling for precision cancer therapies. Genome Med. 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Alieva, M.; van Rheenen, J.; Broekman, M.L.D. Potential impact of invasive surgical procedures on primary tumor growth and metastasis. Clin. Exp. Metastasis 2018, 35, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Gilson, P.; Merlin, J.L.; Harle, A. Deciphering Tumour Heterogeneity: From Tissue to Liquid Biopsy. Cancers 2022, 14, 1384. [Google Scholar] [CrossRef] [PubMed]
- Castro-Giner, F.; Gkountela, S.; Donato, C.; Alborelli, I.; Quagliata, L.; Ng, C.K.Y.; Piscuoglio, S.; Aceto, N. Cancer Diagnosis Using a Liquid Biopsy: Challenges and Expectations. Diagnostics 2018, 8, 31. [Google Scholar] [CrossRef]
- Lone, S.N.; Nisar, S.; Masoodi, T.; Singh, M.; Rizwan, A.; Hashem, S.; El-Rifai, W.; Bedognetti, D.; Batra, S.K.; Haris, M.; et al. Liquid biopsy: A step closer to transform diagnosis, prognosis and future of cancer treatments. Mol. Cancer 2022, 21, 79. [Google Scholar] [CrossRef] [PubMed]
- Aravanis, A.M.; Lee, M.; Klausner, R.D. Next-Generation Sequencing of Circulating Tumor DNA for Early Cancer Detection. Cell 2017, 168, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Nikanjam, M.; Kato, S.; Kurzrock, R. Liquid biopsy: Current technology and clinical applications. J. Hematol. Oncol. 2022, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Xu, T.; Zhu, H.; Yu, J. Clinical Applications of Cerebrospinal Fluid Circulating Tumor DNA as a Liquid Biopsy for Central Nervous System Tumors. Onco Targets Ther. 2020, 13, 719–731. [Google Scholar] [CrossRef]
- Han, H.S.; Lee, K.W. Liquid Biopsy: An Emerging Diagnostic, Prognostic, and Predictive Tool in Gastric Cancer. J. Gastric Cancer 2024, 24, 4–28. [Google Scholar] [CrossRef]
- Khurshid, Z.; Zafar, M.S.; Khan, R.S.; Najeeb, S.; Slowey, P.D.; Rehman, I.U. Role of Salivary Biomarkers in Oral Cancer Detection. Adv. Clin. Chem. 2018, 86, 23–70. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, D.; Li, S.; Ren, J.; Huang, W.; Liu, D.; Wang, W. Bronchoalveolar lavage fluid assessment facilitates precision medicine for lung cancer. Cancer Biol. Med. 2023, 21, 230–251. [Google Scholar] [CrossRef] [PubMed]
- Oshi, M.; Murthy, V.; Takahashi, H.; Huyser, M.; Okano, M.; Tokumaru, Y.; Rashid, O.M.; Matsuyama, R.; Endo, I.; Takabe, K. Urine as a Source of Liquid Biopsy for Cancer. Cancers 2021, 13, 2652. [Google Scholar] [CrossRef] [PubMed]
- Heitzer, E.; Perakis, S.; Geigl, J.B.; Speicher, M.R. The potential of liquid biopsies for the early detection of cancer. NPJ Precis. Oncol. 2017, 1, 36. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, M.; Ren, S.; Liu, Y.; Zhang, J.; Li, S.; Gao, W.; Gong, X.; Liu, J.; Wang, Y.; et al. Exploring genetic alterations in circulating tumor DNA from cerebrospinal fluid of pediatric medulloblastoma. Sci. Rep. 2021, 11, 5638. [Google Scholar] [CrossRef] [PubMed]
- Chicard, M.; Colmet-Daage, L.; Clement, N.; Danzon, A.; Bohec, M.; Bernard, V.; Baulande, S.; Bellini, A.; Deveau, P.; Pierron, G.; et al. Whole-Exome Sequencing of Cell-Free DNA Reveals Temporo-spatial Heterogeneity and Identifies Treatment-Resistant Clones in Neuroblastoma. Clin. Cancer Res. 2018, 24, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Connal, S.; Cameron, J.M.; Sala, A.; Brennan, P.M.; Palmer, D.S.; Palmer, J.D.; Perlow, H.; Baker, M.J. Liquid biopsies: The future of cancer early detection. J. Transl. Med. 2023, 21, 118. [Google Scholar] [CrossRef]
- Vasseur, D.; Sassi, H.; Bayle, A.; Tagliamento, M.; Besse, B.; Marzac, C.; Arbab, A.; Auger, N.; Cotteret, S.; Aldea, M.; et al. Next-Generation Sequencing on Circulating Tumor DNA in Advanced Solid Cancer: Swiss Army Knife for the Molecular Tumor Board? A Review of the Literature Focused on FDA Approved Test. Cells 2022, 11, 1901. [Google Scholar] [CrossRef]
- Andersson, D.; Fagman, H.; Dalin, M.G.; Stahlberg, A. Circulating cell-free tumor DNA analysis in pediatric cancers. Mol. Aspects Med. 2020, 72, 100819. [Google Scholar] [CrossRef] [PubMed]
- Miguez, A.C.K.; Barros, B.D.F.; de Souza, J.E.S.; da Costa, C.M.L.; Cunha, I.W.; Barbosa, P.; Apezzato, M.L.P.; de Souza, S.J.; Carraro, D.M. Assessment of somatic mutations in urine and plasma of Wilms tumor patients. Cancer Med. 2020, 9, 5948–5959. [Google Scholar] [CrossRef]
- Eibl, R.H.; Schneemann, M. Liquid Biopsy and Primary Brain Tumors. Cancers 2021, 13, 5429. [Google Scholar] [CrossRef]
- Huang, T.Y.; Piunti, A.; Lulla, R.R.; Qi, J.; Horbinski, C.M.; Tomita, T.; James, C.D.; Shilatifard, A.; Saratsis, A.M. Detection of Histone H3 mutations in cerebrospinal fluid-derived tumor DNA from children with diffuse midline glioma. Acta Neuropathol. Commun. 2017, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.M.; Shah, R.H.; Pentsova, E.I.; Pourmaleki, M.; Briggs, S.; Distefano, N.; Zheng, Y.; Skakodub, A.; Mehta, S.A.; Campos, C.; et al. Tracking tumour evolution in glioma through liquid biopsies of cerebrospinal fluid. Nature 2019, 565, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Escudero, L.; Llort, A.; Arias, A.; Diaz-Navarro, A.; Martinez-Ricarte, F.; Rubio-Perez, C.; Mayor, R.; Caratu, G.; Martinez-Saez, E.; Vazquez-Mendez, E.; et al. Circulating tumour DNA from the cerebrospinal fluid allows the characterisation and monitoring of medulloblastoma. Nat. Commun. 2020, 11, 5376. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ji, H.; Yang, W.; Zhang, M.; Guo, Y.; Li, B.; Wang, J.; Chen, R.; Chen, Y.; Wang, X. Liquid biopsy using ascitic fluid and pleural effusion supernatants for genomic profiling in gastrointestinal and lung cancers. BMC Cancer 2022, 22, 1020. [Google Scholar] [CrossRef] [PubMed]
- Lousada-Fernandez, F.; Rapado-Gonzalez, O.; Lopez-Cedrun, J.L.; Lopez-Lopez, R.; Muinelo-Romay, L.; Suarez-Cunqueiro, M.M. Liquid Biopsy in Oral Cancer. Int. J. Mol. Sci. 2018, 19, 1704. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, H.; Spaepen, M.; Mc Bride, W.H.; Nuyts, S. The clinical relevance of microsatellite alterations in head and neck squamous cell carcinoma: A critical review. Eur. J. Hum. Genet. 2007, 15, 734–741. [Google Scholar] [CrossRef]
- Chiangjong, W.; Bhakdi, S.C.; Woramongkolchai, N.; Vanichapol, T.; Pongsakul, N.; Hongeng, S.; Chutipongtanate, S. Cell-Main Spectra Profile Screening Technique in Simulation of Circulating Tumour Cells Using MALDI-TOF Mass Spectrometry. Cancers 2021, 13, 3775. [Google Scholar] [CrossRef]
- Hu, X.; Zang, X.; Lv, Y. Detection of circulating tumor cells: Advances and critical concerns. Oncol. Lett. 2021, 21, 422. [Google Scholar] [CrossRef]
- Yang, R.; Zheng, S.; Dong, R. Circulating tumor cells in neuroblastoma: Current status and future perspectives. Cancer Med. 2023, 12, 7–19. [Google Scholar] [CrossRef]
- Sanders, D.G.; Wiley, F.M.; Moss, T.J. Serial immunocytologic analysis of blood for tumor cells in two patients with neuroblastoma. Cancer 1991, 67, 1423–1427. [Google Scholar] [CrossRef]
- Magbanua, M.J.; Park, J.W. Isolation of circulating tumor cells by immunomagnetic enrichment and fluorescence-activated cell sorting (IE/FACS) for molecular profiling. Methods 2013, 64, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Burchill, S.A.; Bradbury, F.M.; Smith, B.; Lewis, I.J.; Selby, P. Neuroblastoma cell detection by reverse transcriptase-polymerase chain reaction (RT-PCR) for tyrosine hydroxylase mRNA. Int. J. Cancer 1994, 57, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Zhang, B.; Zheng, Y.; Zheng, C.; Liu, B.; Zheng, S.; Dong, K.; Dong, R. Circulating tumor cells detection in neuroblastoma patients by EpCAM-independent enrichment and immunostaining-fluorescence in situ hybridization. EBioMedicine 2018, 35, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Haber, D.A.; Velculescu, V.E. Blood-based analyses of cancer: Circulating tumor cells and circulating tumor DNA. Cancer Discov. 2014, 4, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Su, L.; Qian, C. Circulating tumor DNA: A promising biomarker in the liquid biopsy of cancer. Oncotarget 2016, 7, 48832–48841. [Google Scholar] [CrossRef]
- Doculara, L.; Trahair, T.N.; Bayat, N.; Lock, R.B. Circulating Tumor DNA in Pediatric Cancer. Front. Mol. Biosci. 2022, 9, 885597. [Google Scholar] [CrossRef] [PubMed]
- Marczynski, G.T.; Laus, A.C.; Dos Reis, M.B.; Reis, R.M.; Vazquez, V.L. Circulating tumor DNA (ctDNA) detection is associated with shorter progression-free survival in advanced melanoma patients. Sci. Rep. 2020, 10, 18682. [Google Scholar] [CrossRef]
- Kahana-Edwin, S.; Cain, L.E.; McCowage, G.; Darmanian, A.; Wright, D.; Mullins, A.; Saletta, F.; Karpelowsky, J. Neuroblastoma Molecular Risk-Stratification of DNA Copy Number and ALK Genotyping via Cell-Free Circulating Tumor DNA Profiling. Cancers 2021, 13, 3365. [Google Scholar] [CrossRef]
- Krumbholz, M.; Hellberg, J.; Steif, B.; Bauerle, T.; Gillmann, C.; Fritscher, T.; Agaimy, A.; Frey, B.; Juengert, J.; Wardelmann, E.; et al. Genomic EWSR1 Fusion Sequence as Highly Sensitive and Dynamic Plasma Tumor Marker in Ewing Sarcoma. Clin. Cancer Res. 2016, 22, 4356–4365. [Google Scholar] [CrossRef]
- Kjaer, E.K.R.; Vase, C.B.; Rossing, M.; Ahlborn, L.B.; Hjalgrim, L.L. Detection of circulating tumor-derived material in peripheral blood of pediatric sarcoma patients: A systematic review. Transl. Oncol. 2023, 34, 101690. [Google Scholar] [CrossRef]
- Jimenez, I.; Chicard, M.; Colmet-Daage, L.; Clement, N.; Danzon, A.; Lapouble, E.; Pierron, G.; Bohec, M.; Baulande, S.; Berrebi, D.; et al. Circulating tumor DNA analysis enables molecular characterization of pediatric renal tumors at diagnosis. Int. J. Cancer 2019, 144, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Tombolan, L.; Rossi, E.; Binatti, A.; Zin, A.; Manicone, M.; Facchinetti, A.; Lucchetta, S.; Affinita, M.C.; Bonvini, P.; Bortoluzzi, S.; et al. Clinical significance of circulating tumor cells and cell-free DNA in pediatric rhabdomyosarcoma. Mol. Oncol. 2022, 16, 2071–2085. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.L.; Xu, L.; Kooi, I.; Murphree, A.L.; Prabakar, R.K.; Reid, M.; Stachelek, K.; Le, B.H.A.; Welter, L.; Reiser, B.J.; et al. Genomic cfDNA Analysis of Aqueous Humor in Retinoblastoma Predicts Eye Salvage: The Surrogate Tumor Biopsy for Retinoblastoma. Mol. Cancer Res. 2018, 16, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Leung, F.; Kulasingam, V.; Diamandis, E.P.; Hoon, D.S.; Kinzler, K.; Pantel, K.; Alix-Panabieres, C. Circulating Tumor DNA as a Cancer Biomarker: Fact or Fiction? Clin. Chem. 2016, 62, 1054–1060. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Cortes, J.; Santarpia, L.; Vivancos, A.; Tabernero, J.; Reis-Filho, J.S.; Seoane, J. Circulating tumour cells and cell-free DNA as tools for managing breast cancer. Nat. Rev. Clin. Oncol. 2013, 10, 377–389. [Google Scholar] [CrossRef]
- Panachan, J.; Rojsirikulchai, N.; Pongsakul, N.; Khowawisetsut, L.; Pongphitcha, P.; Siriboonpiputtana, T.; Chareonsirisuthigul, T.; Phornsarayuth, P.; Klinkulab, N.; Jinawath, N.; et al. Extracellular Vesicle-Based Method for Detecting MYCN Amplification Status of Pediatric Neuroblastoma. Cancers 2022, 14, 2627. [Google Scholar] [CrossRef]
- Chaemsaithong, P.; Luewan, S.; Taweevisit, M.; Chiangjong, W.; Pongchaikul, P.; Thorner, P.S.; Tongsong, T.; Chutipongtanate, S. Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease. Int. J. Mol. Sci. 2023, 24, 5658. [Google Scholar] [CrossRef]
- Chutipongtanate, S.; Pattanapanyasat, K.; Meller, J.; Serrano Lopez, J. Editorial: Driving extracellular vesicles toward applications in precision medicine. Front. Med. (Lausanne) 2022, 9, 1049697. [Google Scholar] [CrossRef]
- Yu, W.; Hurley, J.; Roberts, D.; Chakrabortty, S.K.; Enderle, D.; Noerholm, M.; Breakefield, X.O.; Skog, J.K. Exosome-based liquid biopsies in cancer: Opportunities and challenges. Ann. Oncol. 2021, 32, 466–477. [Google Scholar] [CrossRef]
- Pitt, J.M.; Kroemer, G.; Zitvogel, L. Extracellular vesicles: Masters of intercellular communication and potential clinical interventions. J. Clin. Investig. 2016, 126, 1139–1143. [Google Scholar] [CrossRef]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a new frontier of cancer liquid biopsy. Mol. Cancer 2022, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Dawson, S.J.; Tsui, D.W.; Murtaza, M.; Biggs, H.; Rueda, O.M.; Chin, S.F.; Dunning, M.J.; Gale, D.; Forshew, T.; Mahler-Araujo, B.; et al. Analysis of circulating tumor DNA to monitor metastatic breast cancer. N. Engl. J. Med. 2013, 368, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Cabus, L.; Lagarde, J.; Curado, J.; Lizano, E.; Perez-Boza, J. Current challenges and best practices for cell-free long RNA biomarker discovery. Biomark. Res. 2022, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.C.M.; Heider, K.; Gale, D.; Murphy, S.; Fisher, E.; Mouliere, F.; Ruiz-Valdepenas, A.; Santonja, A.; Morris, J.; Chandrananda, D.; et al. ctDNA monitoring using patient-specific sequencing and integration of variant reads. Sci. Transl. Med. 2020, 12, aaz8084. [Google Scholar] [CrossRef] [PubMed]
- Malenica, M.; Vukomanovic, M.; Kurtjak, M.; Masciotti, V.; Dal Zilio, S.; Greco, S.; Lazzarino, M.; Krusic, V.; Percic, M.; Jelovica Badovinac, I.; et al. Perspectives of Microscopy Methods for Morphology Characterisation of Extracellular Vesicles from Human Biofluids. Biomedicines 2021, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrugger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, aau6977. [Google Scholar] [CrossRef]
- Yue, B.; Yang, H.; Wang, J.; Ru, W.; Wu, J.; Huang, Y.; Lan, X.; Lei, C.; Chen, H. Exosome biogenesis, secretion and function of exosomal miRNAs in skeletal muscle myogenesis. Cell Prolif. 2020, 53, e12857. [Google Scholar] [CrossRef]
- Schwager, S.C.; Reinhart-King, C.A. Mechanobiology of microvesicle release, uptake, and microvesicle-mediated activation. Curr. Top. Membr. 2020, 86, 255–278. [Google Scholar] [CrossRef]
- Li, B.; Antonyak, M.A.; Zhang, J.; Cerione, R.A. RhoA triggers a specific signaling pathway that generates transforming microvesicles in cancer cells. Oncogene 2012, 31, 4740–4749. [Google Scholar] [CrossRef]
- Clos-Sansalvador, M.; Monguio-Tortajada, M.; Roura, S.; Franquesa, M.; Borras, F.E. Commonly used methods for extracellular vesicles’ enrichment: Implications in downstream analyses and use. Eur. J. Cell Biol. 2022, 101, 151227. [Google Scholar] [CrossRef] [PubMed]
- Oeyen, E.; Van Mol, K.; Baggerman, G.; Willems, H.; Boonen, K.; Rolfo, C.; Pauwels, P.; Jacobs, A.; Schildermans, K.; Cho, W.C.; et al. Ultrafiltration and size exclusion chromatography combined with asymmetrical-flow field-flow fractionation for the isolation and characterisation of extracellular vesicles from urine. J. Extracell. Vesicles 2018, 7, 1490143. [Google Scholar] [CrossRef]
- Stam, J.; Bartel, S.; Bischoff, R.; Wolters, J.C. Isolation of extracellular vesicles with combined enrichment methods. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2021, 1169, 122604. [Google Scholar] [CrossRef]
- Turner, N.P.; Abeysinghe, P.; Kwan Cheung, K.A.; Vaswani, K.; Logan, J.; Sadowski, P.; Mitchell, M.D. A Comparison of Blood Plasma Small Extracellular Vesicle Enrichment Strategies for Proteomic Analysis. Proteomes 2022, 10, 19. [Google Scholar] [CrossRef]
- Chutipongtanate, S.; Kongsomros, S.; Pongsakul, N.; Panachan, J.; Khowawisetsut, L.; Pattanapanyasat, K.; Hongeng, S.; Thitithanyanont, A. Anti-SARS-CoV-2 effect of extracellular vesicles released from mesenchymal stem cells. J. Extracell. Vesicles 2022, 11, e12201. [Google Scholar] [CrossRef] [PubMed]
- Willms, E.; Cabanas, C.; Mager, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.E. Extracellular Vesicles and Metastasis. Cold Spring Harb. Perspect. Med. 2020, 10, a037275. [Google Scholar] [CrossRef]
- Haug, B.H.; Hald, O.H.; Utnes, P.; Roth, S.A.; Lokke, C.; Flaegstad, T.; Einvik, C. Exosome-like Extracellular Vesicles from MYCN-amplified Neuroblastoma Cells Contain Oncogenic miRNAs. Anticancer Res. 2015, 35, 2521–2530. [Google Scholar]
- Ma, J.; Xu, M.; Yin, M.; Hong, J.; Chen, H.; Gao, Y.; Xie, C.; Shen, N.; Gu, S.; Mo, X. Exosomal hsa-miR199a-3p Promotes Proliferation and Migration in Neuroblastoma. Front. Oncol. 2019, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Colletti, M.; Petretto, A.; Galardi, A.; Di Paolo, V.; Tomao, L.; Lavarello, C.; Inglese, E.; Bruschi, M.; Lopez, A.A.; Pascucci, L.; et al. Proteomic Analysis of Neuroblastoma-Derived Exosomes: New Insights into a Metastatic Signature. Proteomics 2017, 17, 1600430. [Google Scholar] [CrossRef] [PubMed]
- Kaid, C.; Assoni, A.; Marcola, M.; Semedo-Kuriki, P.; Bortolin, R.H.; Carvalho, V.M.; Okamoto, O.K. Proteome and miRNome profiling of microvesicles derived from medulloblastoma cell lines with stem-like properties reveals biomarkers of poor prognosis. Brain Res. 2020, 1730, 146646. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Wu, X.Y.; Liu, X.D.; Zheng, D.F.; Li, H.S.; Yang, B.; Zhang, J.; Chang, Q. Aggressive Medulloblastoma-Derived Exosomal miRNAs Promote In Vitro Invasion and Migration of Tumor Cells Via Ras/MAPK Pathway. J. Neuropathol. Exp. Neurol. 2020, 79, 734–745. [Google Scholar] [CrossRef]
- Jackson, H.K.; Mitoko, C.; Linke, F.; Macarthur, D.; Kerr, I.D.; Coyle, B. Extracellular Vesicles Potentiate Medulloblastoma Metastasis in an EMMPRIN and MMP-2 Dependent Manner. Cancers 2023, 15, 2601. [Google Scholar] [CrossRef]
- Chen, W.W.; Balaj, L.; Liau, L.M.; Samuels, M.L.; Kotsopoulos, S.K.; Maguire, C.A.; Loguidice, L.; Soto, H.; Garrett, M.; Zhu, L.D.; et al. BEAMing and Droplet Digital PCR Analysis of Mutant IDH1 mRNA in Glioma Patient Serum and Cerebrospinal Fluid Extracellular Vesicles. Mol. Ther. Nucleic Acids 2013, 2, e109. [Google Scholar] [CrossRef]
- Yue, X.; Lan, F.; Xia, T. Hypoxic Glioma Cell-Secreted Exosomal miR-301a Activates Wnt/beta-catenin Signaling and Promotes Radiation Resistance by Targeting TCEAL7. Mol. Ther. 2019, 27, 1939–1949. [Google Scholar] [CrossRef]
- Zeng, A.; Wei, Z.; Yan, W.; Yin, J.; Huang, X.; Zhou, X.; Li, R.; Shen, F.; Wu, W.; Wang, X.; et al. Exosomal transfer of miR-151a enhances chemosensitivity to temozolomide in drug-resistant glioblastoma. Cancer Lett. 2018, 436, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zeng, A.; Zhang, Z.; Shi, Z.; Yan, W.; You, Y. Exosomal transfer of miR-1238 contributes to temozolomide-resistance in glioblastoma. EBioMedicine 2019, 42, 238–251. [Google Scholar] [CrossRef]
- Jiao, C.; Jiao, X.; Zhu, A.; Ge, J.; Xu, X. Exosomal miR-34s panel as potential novel diagnostic and prognostic biomarker in patients with hepatoblastoma. J. Pediatr. Surg. 2017, 52, 618–624. [Google Scholar] [CrossRef]
- Liu, W.; Chen, S.; Liu, B. Diagnostic and prognostic values of serum exosomal microRNA-21 in children with hepatoblastoma: A Chinese population-based study. Pediatr. Surg. Int. 2016, 32, 1059–1065. [Google Scholar] [CrossRef]
- Hu, Y.; Zai, H.; Jiang, W.; Yao, Y.; Ou, Z.; Zhu, Q. miR-126 in Extracellular Vesicles Derived from Hepatoblastoma Cells Promotes the Tumorigenesis of Hepatoblastoma through Inducing the Differentiation of BMSCs into Cancer Stem Cells. J. Immunol. Res. 2021, 2021, 6744715. [Google Scholar] [CrossRef]
- Miller, I.V.; Raposo, G.; Welsch, U.; Prazeres da Costa, O.; Thiel, U.; Lebar, M.; Maurer, M.; Bender, H.U.; von Luettichau, I.; Richter, G.H.; et al. First identification of Ewing’s sarcoma-derived extracellular vesicles and exploration of their biological and potential diagnostic implications. Biol. Cell 2013, 105, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Tsugita, M.; Yamada, N.; Noguchi, S.; Yamada, K.; Moritake, H.; Shimizu, K.; Akao, Y.; Ohno, T. Ewing sarcoma cells secrete EWS/Fli-1 fusion mRNA via microvesicles. PLoS ONE 2013, 8, e77416. [Google Scholar] [CrossRef]
- Ventura, S.; Aryee, D.N.; Felicetti, F.; De Feo, A.; Mancarella, C.; Manara, M.C.; Picci, P.; Colombo, M.P.; Kovar, H.; Care, A.; et al. CD99 regulates neural differentiation of Ewing sarcoma cells through miR-34a-Notch-mediated control of NF-kappaB signaling. Oncogene 2016, 35, 3944–3954. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Aiba, H.; Yoshida, T.; Yamamoto, N.; Hayashi, K.; Takeuchi, A.; Miwa, S.; Igarashi, K.; Nguyen, T.D.; Ishii, K.A.; et al. Osteosarcoma-Derived Small Extracellular Vesicles Enhance Tumor Metastasis and Suppress Osteoclastogenesis by miR-146a-5p. Front. Oncol. 2021, 11, 667109. [Google Scholar] [CrossRef] [PubMed]
- Jerez, S.; Araya, H.; Hevia, D.; Irarrazaval, C.E.; Thaler, R.; van Wijnen, A.J.; Galindo, M. Extracellular vesicles from osteosarcoma cell lines contain miRNAs associated with cell adhesion and apoptosis. Gene 2019, 710, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Uotani, K.; Yoshida, A.; Morita, T.; Nezu, Y.; Kobayashi, E.; Yoshida, A.; Uehara, T.; Omori, T.; Sugiu, K.; et al. Clinical significance of circulating miR-25-3p as a novel diagnostic and prognostic biomarker in osteosarcoma. Oncotarget 2017, 8, 33375–33392. [Google Scholar] [CrossRef]
- Zhong, L.; Liao, D.; Li, J.; Liu, W.; Wang, J.; Zeng, C.; Wang, X.; Cao, Z.; Zhang, R.; Li, M.; et al. Rab22a-NeoF1 fusion protein promotes osteosarcoma lung metastasis through its secretion into exosomes. Signal Transduct. Target. Ther. 2021, 6, 59. [Google Scholar] [CrossRef]
- Endo-Munoz, L.; Cai, N.; Cumming, A.; Macklin, R.; Merida de Long, L.; Topkas, E.; Mukhopadhyay, P.; Hill, M.; Saunders, N.A. Progression of Osteosarcoma from a Non-Metastatic to a Metastatic Phenotype Is Causally Associated with Activation of an Autocrine and Paracrine uPA Axis. PLoS ONE 2015, 10, e0133592. [Google Scholar] [CrossRef]
- Ghayad, S.E.; Rammal, G.; Ghamloush, F.; Basma, H.; Nasr, R.; Diab-Assaf, M.; Chelala, C.; Saab, R. Exosomes derived from embryonal and alveolar rhabdomyosarcoma carry differential miRNA cargo and promote invasion of recipient fibroblasts. Sci. Rep. 2016, 6, 37088. [Google Scholar] [CrossRef] [PubMed]
- Fahs, A.; Hussein, N.; Zalzali, H.; Ramadan, F.; Ghamloush, F.; Tamim, H.; El Homsi, M.; Badran, B.; Boulos, F.; Tawil, A.; et al. CD147 Promotes Tumorigenesis via Exosome-Mediated Signaling in Rhabdomyosarcoma. Cells 2022, 11, 2267. [Google Scholar] [CrossRef] [PubMed]
- Rammal, G.; Fahs, A.; Kobeissy, F.; Mechref, Y.; Zhao, J.; Zhu, R.; Diab-Assaf, M.; Saab, R.; Ghayad, S.E. Proteomic Profiling of Rhabdomyosarcoma-Derived Exosomes Yield Insights into Their Functional Role in Paracrine Signaling. J. Proteome Res. 2019, 18, 3567–3579. [Google Scholar] [CrossRef] [PubMed]
- Damanti, C.C.; Ferrone, L.; Gaffo, E.; Garbin, A.; Tosato, A.; Contarini, G.; Gallingani, I.; Angioni, R.; Molon, B.; Borile, G.; et al. Plasma small-extracellular vesicles enriched in miR-122-5p promote disease aggressiveness in pediatric anaplastic large-cell lymphoma. Cancer Commun. 2023, 43, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Garbin, A.; Contarini, G.; Damanti, C.C.; Tosato, A.; Bortoluzzi, S.; Gaffo, E.; Pizzi, M.; Carraro, E.; Lo Nigro, L.; Vinti, L.; et al. MiR-146a-5p enrichment in small-extracellular vesicles of relapsed pediatric ALCL patients promotes macrophages infiltration and differentiation. Biochem. Pharmacol. 2023, 215, 115747. [Google Scholar] [CrossRef] [PubMed]
- Damanti, C.C.; Gaffo, E.; Lovisa, F.; Garbin, A.; Di Battista, P.; Gallingani, I.; Tosato, A.; Pillon, M.; Carraro, E.; Mascarin, M.; et al. MiR-26a-5p as a Reference to Normalize MicroRNA qRT-PCR Levels in Plasma Exosomes of Pediatric Hematological Malignancies. Cells 2021, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Lovisa, F.; Garbin, A.; Crotti, S.; Di Battista, P.; Gallingani, I.; Damanti, C.C.; Tosato, A.; Carraro, E.; Pillon, M.; Mafakheri, E.; et al. Increased Tenascin C, Osteopontin and HSP90 Levels in Plasmatic Small Extracellular Vesicles of Pediatric ALK-Positive Anaplastic Large Cell Lymphoma: New Prognostic Biomarkers? Diagnostics 2021, 11, 253. [Google Scholar] [CrossRef]
- Repetto, O.; Lovisa, F.; Elia, C.; Enderle, D.; Romanato, F.; Buffardi, S.; Sala, A.; Pillon, M.; Steffan, A.; Burnelli, R.; et al. Proteomic Exploration of Plasma Exosomes and Other Small Extracellular Vesicles in Pediatric Hodgkin Lymphoma: A Potential Source of Biomarkers for Relapse Occurrence. Diagnostics 2021, 11, 917. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, J.I.; Dyberg, C.; Wickstrom, M. Neuroblastoma-A Neural Crest Derived Embryonal Malignancy. Front. Mol. Neurosci. 2019, 12, 9. [Google Scholar] [CrossRef]
- Maris, J.M. Recent advances in neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef]
- Honore, N.; Galot, R.; van Marcke, C.; Limaye, N.; Machiels, J.P. Liquid Biopsy to Detect Minimal Residual Disease: Methodology and Impact. Cancers 2021, 13, 5364. [Google Scholar] [CrossRef] [PubMed]
- Orr, B.A. Pathology, diagnostics, and classification of medulloblastoma. Brain Pathol. 2020, 30, 664–678. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Ronvaux, L.; Riva, M.; Coosemans, A.; Herzog, M.; Rommelaere, G.; Donis, N.; D’Hondt, L.; Douxfils, J. Liquid Biopsy in Glioblastoma. Cancers 2022, 14, 3394. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Romero, N.; Carrion-Navarro, J.; Areal-Hidalgo, P.; Ortiz de Mendivil, A.; Asensi-Puig, A.; Madurga, R.; Nunez-Torres, R.; Gonzalez-Neira, A.; Belda-Iniesta, C.; Gonzalez-Rumayor, V.; et al. BRAF V600E Detection in Liquid Biopsies from Pediatric Central Nervous System Tumors. Cancers 2019, 12, 66. [Google Scholar] [CrossRef]
- Wong, H.A.; Fatimy, R.E.; Onodera, C.; Wei, Z.; Yi, M.; Mohan, A.; Gowrisankaran, S.; Karmali, P.; Marcusson, E.; Wakimoto, H.; et al. The Cancer Genome Atlas Analysis Predicts MicroRNA for Targeting Cancer Growth and Vascularization in Glioblastoma. Mol. Ther. 2015, 23, 1234–1247. [Google Scholar] [CrossRef]
- Ranganathan, S.; Lopez-Terrada, D.; Alaggio, R. Hepatoblastoma and Pediatric Hepatocellular Carcinoma: An Update. Pediatr. Dev. Pathol. 2020, 23, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.; Munding, J.; Gruner, M.; Liffers, S.T.; Verdoodt, B.; Hauk, J.; Steinstraesser, L.; Tannapfel, A.; Hermeking, H. Frequent concomitant inactivation of miR-34a and miR-34b/c by CpG methylation in colorectal, pancreatic, mammary, ovarian, urothelial, and renal cell carcinomas and soft tissue sarcomas. Virchows Arch. 2011, 458, 313–322. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, J.; Wu, B.; Wang, X.; Jiang, Y.; Zhu, D. Role of extracellular vesicles in osteosarcoma. Int. J. Med. Sci. 2022, 19, 1216–1226. [Google Scholar] [CrossRef]
- Biazzo, A.; De Paolis, M. Multidisciplinary approach to osteosarcoma. Acta Orthop. Belg. 2016, 82, 690–698. [Google Scholar]
- Chou, A.J.; Gorlick, R. Chemotherapy resistance in osteosarcoma: Current challenges and future directions. Expert Rev. Anticancer Ther. 2006, 6, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Ucci, A.; Cappariello, A.; Ponzetti, M.; Tennant, F.; Loftus, A.E.P.; Shefferd, K.; Maurizi, A.; Delle Monache, S.; Teti, A.; Rucci, N. Anti-osteoblastogenic, pro-inflammatory and pro-angiogenic effect of extracellular vesicles isolated from the human osteosarcoma cell line MNNG/HOS. Bone 2021, 153, 116130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Samuel, G.; Crow, J.; Godwin, A.K.; Zeng, Y. Molecular assessment of circulating exosomes toward liquid biopsy diagnosis of Ewing sarcoma family of tumors. Transl. Res. 2018, 201, 136–153. [Google Scholar] [CrossRef]
- Skapek, S.X.; Ferrari, A.; Gupta, A.A.; Lupo, P.J.; Butler, E.; Shipley, J.; Barr, F.G.; Hawkins, D.S. Rhabdomyosarcoma. Nat. Rev. Dis. Primers 2019, 5, 1. [Google Scholar] [CrossRef]
- Satou, A.; Bennani, N.N.; Feldman, A.L. Update on the classification of T-cell lymphomas, Hodgkin lymphomas, and histiocytic/dendritic cell neoplasms. Expert Rev. Hematol. 2019, 12, 833–843. [Google Scholar] [CrossRef]
- Sandlund, J.T. Non-Hodgkin Lymphoma in Children. Curr. Hematol. Malig. Rep. 2015, 10, 237–243. [Google Scholar] [CrossRef]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wills, C.A.; Chen, L.; Zhang, J.; Zhao, Y.; Zhou, M.; Sundstrom, J.M.; Schell, T.; Spiegelman, V.S.; Young, M.M.; et al. Small extracellular vesicles induce resistance to anti-GD2 immunotherapy unveiling tipifarnib as an adjunct to neuroblastoma immunotherapy. J. Immunother. Cancer 2022, 10, e004399. [Google Scholar] [CrossRef]
- Chen, W.; Hao, X.; Yang, B.; Zhang, Y.; Sun, L.; Hua, Y.; Yang, L.; Yu, J.; Zhao, J.; Hou, L.; et al. MYCN-amplified neuroblastoma cell-derived exosomal miR-17-5p promotes proliferation and migration of non-MYCN amplified cells. Mol. Med. Rep. 2021, 23, 245. [Google Scholar] [CrossRef]
- Challagundla, K.B.; Wise, P.M.; Neviani, P.; Chava, H.; Murtadha, M.; Xu, T.; Kennedy, R.; Ivan, C.; Zhang, X.; Vannini, I.; et al. Exosome-mediated transfer of microRNAs within the tumor microenvironment and neuroblastoma resistance to chemotherapy. J. Natl. Cancer Inst. 2015, 107, djv135. [Google Scholar] [CrossRef]
- Gassmann, H.; Schneider, K.; Evdokimova, V.; Ruzanov, P.; Schober, S.J.; Xue, B.; von Heyking, K.; Thiede, M.; Richter, G.H.S.; Pfaffl, M.W.; et al. Ewing Sarcoma-Derived Extracellular Vesicles Impair Dendritic Cell Maturation and Function. Cells 2021, 10, 2081. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.A.; Koh, E.J.; Kim, R.N.; Byun, J.W.; Phi, J.H.; Yang, J.; Wang, K.C.; Park, A.K.; Hwang, D.W.; Lee, J.Y.; et al. Extracellular vesicle-associated miR-135b and -135a regulate stemness in Group 4 medulloblastoma cells by targeting angiomotin-like 2. Cancer Cell Int. 2020, 20, 558. [Google Scholar] [CrossRef] [PubMed]
- Epple, L.M.; Griffiths, S.G.; Dechkovskaia, A.M.; Dusto, N.L.; White, J.; Ouellette, R.J.; Anchordoquy, T.J.; Bemis, L.T.; Graner, M.W. Medulloblastoma exosome proteomics yield functional roles for extracellular vesicles. PLoS ONE 2012, 7, e42064. [Google Scholar] [CrossRef] [PubMed]
- Purvis, I.J.; Velpula, K.K.; Guda, M.R.; Nguyen, D.; Tsung, A.J.; Asuthkar, S. B7-H3 in Medulloblastoma-Derived Exosomes; A Novel Tumorigenic Role. Int. J. Mol. Sci. 2020, 21, 7050. [Google Scholar] [CrossRef] [PubMed]
- Bisaro, B.; Mandili, G.; Poli, A.; Piolatto, A.; Papa, V.; Novelli, F.; Cenacchi, G.; Forni, M.; Zanini, C. Proteomic analysis of extracellular vesicles from medullospheres reveals a role for iron in the cancer progression of medulloblastoma. Mol. Cell Ther. 2015, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Dhamdhere, M.R.; Gowda, C.P.; Singh, V.; Liu, Z.; Carruthers, N.; Grant, C.N.; Sharma, A.; Dovat, S.; Sundstrom, J.M.; Wang, H.G.; et al. IGF2BP1 regulates the cargo of extracellular vesicles and promotes neuroblastoma metastasis. Oncogene 2023, 42, 1558–1571. [Google Scholar] [CrossRef]
- Mancarella, C.; Giusti, V.; Caldoni, G.; Laginestra, M.A.; Parra, A.; Toracchio, L.; Giordano, G.; Roncuzzi, L.; Piazzi, M.; Blalock, W.; et al. Extracellular vesicle-associated IGF2BP3 tunes Ewing sarcoma cell migration and affects PI3K/Akt pathway in neighboring cells. Cancer Gene Ther. 2023, 30, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Macklin, R.; Wang, H.; Loo, D.; Martin, S.; Cumming, A.; Cai, N.; Lane, R.; Ponce, N.S.; Topkas, E.; Inder, K.; et al. Extracellular vesicles secreted by highly metastatic clonal variants of osteosarcoma preferentially localize to the lungs and induce metastatic behaviour in poorly metastatic clones. Oncotarget 2016, 7, 43570–43587. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.L.; Turlapati, R.; Liu, T.; Schulte, J.H.; Huttelmaier, S. IGF2BP1 harbors prognostic significance by gene gain and diverse expression in neuroblastoma. J. Clin. Oncol. 2015, 33, 1285–1293. [Google Scholar] [CrossRef]
- Xue, P.; Huang, S.; Han, X.; Zhang, C.; Yang, L.; Xiao, W.; Fu, J.; Li, H.; Zhou, Y. Exosomal miR-101-3p and miR-423-5p inhibit medulloblastoma tumorigenesis through targeting FOXP4 and EZH2. Cell Death Differ. 2022, 29, 82–95. [Google Scholar] [CrossRef]
- Huang, S.; Xue, P.; Han, X.; Zhang, C.; Yang, L.; Liu, L.; Wang, X.; Li, H.; Fu, J.; Zhou, Y. Exosomal miR-130b-3p targets SIK1 to inhibit medulloblastoma tumorigenesis. Cell Death Dis. 2020, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Romero, N.; Esteban-Rubio, S.; Rackov, G.; Carrion-Navarro, J.; Belda-Iniesta, C.; Ayuso-Sacido, A. Extracellular vesicles compartment in liquid biopsies: Clinical application. Mol. Aspects Med. 2018, 60, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Irmer, B.; Chandrabalan, S.; Maas, L.; Bleckmann, A.; Menck, K. Extracellular Vesicles in Liquid Biopsies as Biomarkers for Solid Tumors. Cancers 2023, 15, 1307. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Previously identified EV biomarkers for developing pediatric cancer liquid biopsies. An illustration depicting biomolecule-containing EVs from transcriptomics and proteomics investigations in medulloblastoma (mainly arising in the cerebellum), glioma (commonly arising in the cerebral cortex), lymphoma (arising in lymph nodes of the neck, chest (mediastinum), axilla, abdomen, and groin), hepatoblastoma (in the liver), neuroblastoma (mainly arising from neural crest-derived cells), Ewing’s sarcoma (commonly arising in bones of the pelvis, legs, arms, and chest wall), osteosarcoma (mainly found in the femur, tibia, and humerus), and rhabdomyosarcoma (mainly found in skeletal muscles in the arms, legs, head, neck, and abdomen). The figure was created with BioRender.com.
Figure 1.
Previously identified EV biomarkers for developing pediatric cancer liquid biopsies. An illustration depicting biomolecule-containing EVs from transcriptomics and proteomics investigations in medulloblastoma (mainly arising in the cerebellum), glioma (commonly arising in the cerebral cortex), lymphoma (arising in lymph nodes of the neck, chest (mediastinum), axilla, abdomen, and groin), hepatoblastoma (in the liver), neuroblastoma (mainly arising from neural crest-derived cells), Ewing’s sarcoma (commonly arising in bones of the pelvis, legs, arms, and chest wall), osteosarcoma (mainly found in the femur, tibia, and humerus), and rhabdomyosarcoma (mainly found in skeletal muscles in the arms, legs, head, neck, and abdomen). The figure was created with BioRender.com.

Figure 2.
Potential roles of EVs in pediatric cancer tumorigenesis and metastasis. An illustration depicting the involvement of pathologic EVs that promote proliferation and metastasis, modulate the tumor microenvironment, and induce resistance to chemotherapy in various types of pediatric cancers. This figure was created with BioRender.com.
Figure 2.
Potential roles of EVs in pediatric cancer tumorigenesis and metastasis. An illustration depicting the involvement of pathologic EVs that promote proliferation and metastasis, modulate the tumor microenvironment, and induce resistance to chemotherapy in various types of pediatric cancers. This figure was created with BioRender.com.

Table 2.
Transcript and protein detections from EV studies of cell lines and patient-derived specimens that have been proposed as potential molecules for EV-based liquid biopsy in pediatric cancers.
Table 2.
Transcript and protein detections from EV studies of cell lines and patient-derived specimens that have been proposed as potential molecules for EV-based liquid biopsy in pediatric cancers.
| Cancer Type | Vesicle Type | Transcript | Protein | Main Finding | References |
|---|---|---|---|---|---|
| Neuroblastoma | LEVs derived from MYCN-amplified neuroblastoma cells and patient-derived bone marrow plasma | X | MYCN mRNA was detectable in LEVs, but not sEVs, of MYCN-amplified neuroblastoma cell lines and patient-derived bone marrow plasma, where the presence and absence of MYCN-LEVs were associated with MYCN amplification status and treatment–relapse disease states. | [46] | |
| sEVs derived from MYCN-amplified neuroblastoma cell lines | X | miR-92a was the highest expressed in EV derived from the neuroblastoma cell line. | [70] | ||
| sEVs derived from plasma of healthy and neuroblastoma patients | X | RT-qPCR validation discovered that sEVs (exosome) containing miR-199a-3p were significantly higher expressed in neuroblastoma patients compared with the healthy donors. | [71] | ||
| sEVs isolated from neuroblastoma cells derived from abdominal primary tumors and bone marrow metastasis | X | Six proteins uniquely presented in metastatic neuroblastoma sEVs, e.g., signal peptidase complex catalytic subunit SEC11, cell division cycle-associated protein 3, nuclear pore complex protein Nup107, calcium, and integrin-binding protein 1. EV proteins of primary tumors are involved in neuronal development and function, while proteins exclusively present in EVs derived from neuroblastoma–bone marrow metastatic models are associated with cell survival, proliferation, and progression. | [72] | ||
| Medulloblastoma | LEVs derived from highly aggressive stem-like medulloblastoma cells overexpressing the pluripotent factor OCT4A | X | X | The interactome analysis of distinct proteins and miRNA suggested that ERK, PI3K/AKT/mTOR, EGF/EFGR, and stem cell self-renewal are the main oncogenic signaling pathways altered in these aggressive medulloblastoma cells. LEVs carried four proteins (UBE2M, HNRNPCL2, HNRNPCL3, and HNRNPCL4) and five miRNAs (miR-4449, miR-500b, miR-3648, miR-1291, and miR-3607). | [73] |
| sEVs derived from group 3 medulloblastoma cell lines | X | sEVs (Exosome) derived from the group 3 medulloblastoma cell line with increased levels of miR-181a-5p, miR-125b-5p, and let-7b-5p could promote in vitro invasion and migratory abilities of a less invasive medulloblastoma cell line through the activation of ERK in the Ras/MAPK pathway. | [74] | ||
| sEVs derived from metastatic medulloblastoma cell lines compared with non-metastatic medulloblastoma cell lines | X | The sEVs derived from the metastatic medulloblastoma cell line had significantly increased in MMP-2 localized on their external surface. This study found a high level of MMP-2 activity in CSF from three of four patients associated with tumor progression. | [75] | ||
| Glioma | sEVs derived from CSF and serum of glioma patients | X | A combination of biofluid EV-derived RNA and BEAMing RT-PCR could detect and quantify mutant and wild-type IDH1 RNA transcripts in CSF of patients with gliomas. | [76] | |
| sEVs derived from serum or conditioned media of glioma patients | X | Hypoxic GBM cells secrete sEVs containing miR-301a, which can promote radiation resistance in normoxia-cultured cells. Hypoxic sEVs containing miR-301a directly targeted GBM tumor suppressor TCEAL7 genes and actively suppressed their expression in normoxic glioma cells. | [77] | ||
| sEVs derived from temozolomide (TMZ)-resistant glioblastoma multiforme cells | X | The lower expression of miR-151 in sEVs was related to an increased resistance to temozolomide (TMZ), in which the restoration of miR-151a expression sensitized TMZ-resistant glioblastoma multiforme cells. | [78] | ||
| sEVs derived from TMZ-resistant glioblastoma cells | X | The high expression of sEVs containing miR-1238 led to the acquired resistance against temozolomide in glioblastoma-sensitive cells. | [79] | ||
| Hepatoblastoma | sEVs derived from the serum of hepatoblastoma patients | X | The level of sEV-derived miR-34a/b/c was significantly lower in the serum of patients with hepatoblastoma compared to healthy control groups. | [80] | |
| sEVs derived from plasma of pediatric hepatoblastoma patients | X | The elevated level of miR-21 containing sEVs in hepatoblastoma patients might be another biomarker for hepatoblastoma. | [81] | ||
| sEVs derived from the hepatoblastoma cell line | X | sEV-derived miR-126 was upregulated in hepatoblastoma cells, which suggested that this microRNA promoted the tumorigenesis of liver cancer. | [82] | ||
| Ewing’s sarcoma | sEVs derived from Ewing’s sarcoma cell line | X | RT-qPCR detected ES-specific transcripts such as EWSR1-FLI1 from Ewing’s sarcoma-derived EVs. | [83] | |
| LEVs derived from Ewing’s sarcoma cell of xenografted mice | X | EWS/FLI1 fusion mRNA (resulting from the t(11;22) (q24;q12) translocation) could be identified in MVs derived from Ewing sarcoma cells and was also detectable in MVs from plasma samples of ES cell-xenografted animals. | [84] | ||
| sEVs isolated from silenced CD99 expression of patients derived Ewing’s sarcoma cell line | X | Ewing sarcoma cells with silenced CD99 expression released EVs containing high levels of miR-34a. | [85] | ||
| Osteosarcoma | sEVs derived from malignant human osteosarcoma cell lines | X | Small extracellular vesicles derived from malignant human osteosarcomas contain miR-146a-5p. | [86] | |
| sEVs derived from six different human osteosarcoma or osteoblast cell lines | X | Next-generation miRNA sequencing revealed miRNAs in cell lines with different degrees of metastatic potential and found that mi-21-5p, miR-143-3p, miR-148a-3p, and 181a-5p are highly expressed in sEV-derived metastatic SAOS2 cells. | [87] | ||
| sEVs derived from the osteosarcoma cell line | X | Circulating miR-17-5p and miR-25-3p could be identified in osteosarcoma cells, which are used as a novel diagnostic and prognostic biomarker and also reflect tumor burden in the osteosarcoma mouse model. | [88] | ||
| sEVs derived from the Rab22a-NeoF1 fusion protein cell line | X | The osteosarcoma Rab22a-NeoF1 fusion protein was secreted via EV by binding to the KFERQ-like motif of HSP90, which was taken up by macrophages and other cancer cells. EV derived from osteosarcoma contained programmed death-ligand 1 (PD-L1) and N-cadherin. | [89] | ||
| sEVs derived from the metastatic osteosarcoma cell line | X | The elevation of uPA and its receptor (uPAR) was cargo via sEVs secreted from metastatic osteosarcoma cells. | [90] | ||
| Rhabdomyosarcoma | sEVs derived from rhabdomyosarcoma cell lines | X | Ten miRNAs were common among the two rhabdomyosarcoma cell lines (JR1 and RD), while only two miRNAs (miR-1246 and miR-1268) were present in EVs of all cell lines. | [91] | |
| sEV derived from the rhabdomyosarcoma cell lines | X | CD147 was exclusively expressed in metastatic tumors of human rhabdomyosarcoma tissue, which was involved in modulating the microenvironment through rhabdomyosarcoma-secreted sEVs. | [92] | ||
| sEV derived from alveolar and embryonal rhabdomyosarcoma cell lines | X | A proteomic study revealed 122 common proteins in alveolar rhabdomyosarcoma-derived EVs and 161 common proteins in embryonal rhabdomyosarcoma-derived EVs. The biological process analysis suggested that 81 proteins were common to both subtypes, which involved cell signaling, cell movement, and cancer. | [93] | ||
| Pediatric lymphoma | sEVs derived from plasma of ALCL patients | X | miR-122-5p was elevated in plasma sEVs derived from ALCL patients, which are critical in promoting tumor cell dissemination and aggressiveness. | [94] | |
| sEVs derived from plasma of 20 pediatric anaplastic lymphoma kinase-positive ALCL patients | X | Small RNA-sequencing analysis in plasma sEVs from 20 pediatric ALCA suggested that miR-146a-5p and miR-378a-3p showed a negative prognostic impact in both univariate and multivariate analysis. | [95] | ||
| sEVs derived from Burkitt lymphoma, Hodgkin lymphoma, and mature B-cell acute lymphoblastic leukemia | X | Burkitt lymphoma, Hodgkin lymphoma, and mature B-cell acute lymphoblastic leukemia most stably expressed miR-26a-5p and miR-486-5p in sEVs. | [96] | ||
| sEVs derived from plasma pediatric ALCL patients and healthy donors | X | The Reactome database and KEGG networks highlighted a dramatic increase in proteins of the PI3K/AKT pathway in ALCL-sEVs, which included heat shock protein 90-kDa isoform alpha-1, osteopontin, and tenascin plasma EV derived from pediatric ALCL patients. | [97] | ||
| sEVs derived from plasma of non-relapsed and relapsed nodular sclerosis Hodgkin lymphoma | X | LC-MS/MS identified these 11 unique protein spots, including five more abundant in non-relapsed HL (e.g., isoform 2 preproprotein of complement C4-A, complement C4-B, fibrinogen γ chain, inter-α-trypsin inhibitor heavy chain H2, and immunoglobulin heavy chain constant region mu) and six more abundant in relapsed HL (e.g., apolipoprotein A-I, apolipoprotein A-IV, clusterin, haptoglobin, α-1-acid glycoprotein 1, and transthyretin). | [98] |
Table 3.
The relationship between EV-based liquid biopsy, their molecular cargos, and functional involvement in childhood cancer tumorigenesis and metastasis.
Table 3.
The relationship between EV-based liquid biopsy, their molecular cargos, and functional involvement in childhood cancer tumorigenesis and metastasis.
| Disease Stage | Disease | Significant Biology Process | Vesicle Type | EV Molecular Cargo | Main Finding | References |
|---|---|---|---|---|---|---|
| Tumorigenesis | Neuroblastoma | Regulation of infiltrated immune cells after chemotherapy treatment | sEVs | Not specified | Small EVs suppressed splenic NK cell maturation in vivo and dinutuximab-induced NK cell-mediated antibody-dependent cellular cytotoxicity in vitro upon dinutuximab treatment. | [118] |
| Facilitating the migration and proliferation of non-MYCN amplified cells | sEVs released by MYCN-amplified neuroblastoma cells | miR-17-5p | miR-17-5p is crucial in facilitating the migration and proliferation of non-MYCN amplified cells. | [119] | ||
| Contribute to resistance to chemotherapy and promote cancer progression | sEVs derived from neuroblastoma cell line | miR-21, miR-155 | The data presented in this study demonstrate the distinct function of EV-derived miR-21 and miR-155 in intercellular communication between neuroblastoma cells and human monocytes, contributing to developing resistance to chemotherapy and cancer progression. | [120] | ||
| Ewing’s sarcoma | Regulation of pro-inflammatory response | sEVs derived from Ewing’s sarcoma cell lines | Not specified | Exposure to Ewing’s sarcoma EVs inhibited the process of cellular development towards moDCs, as indicated by the decreased expression levels of co-stimulatory molecules (e.g., CD80, CD86, and HLA-DR). Ewing’s sarcoma EVs exhibited the ability to suppress the proliferation of CD4+ and CD8+ T cells, as well as the release of IFNγ, while simultaneously increasing the secretion of IL-10 and IL-6. | [121] | |
| Rhabdosarcoma | Promotes tumor cell aggressiveness and modulates the microenvironment | sEV derived from rhabdomyosarcoma cell lines | CD147 | Treatment of normal fibroblasts with rhabdomyosarcoma-derived EVs increased proliferation, migration, and invasion, whereas CD147-downregulated rhabdomyosarcoma cells block these effects. | [92] | |
| Medulloblastoma | Preserve stem cell characteristics or communicate with neighboring cells to promote the progression of group 4 of medulloblastoma | sEVs derived from bulk tumor cells and brain tumor sheroid-forming cells | miR-135b, miR-135a | The suppression of miR-135b and miR-135a leads to a decrease in the stemness of medulloblastoma brain tumor spheroid-forming cells, in which AMOTL2 was targeted by miR-135b and miR-135a. | [122] | |
| Promote in vitro invasion and migratory capacities of tumor cells via activating the ERK pathway in the Ras/MAPK signaling cascade | sEVs derived from Group 3 medulloblastoma cell lines | miR-181a-5p, miR-125b-5p, let-7b-5p | The upregulation of these miRNAs led to more significant in vitro invasion and migratory capacities of tumor cells via activating the ERK pathway in the Ras/MAPK signaling cascade. | [74] | ||
| Stimulate the proliferation of tumor cells, facilitate migration, and regulate T cell responses in vitro | sEVs derived from medulloblastoma cell lines | Not specified | The examination of the functional properties of EVs can stimulate the proliferation of tumor cells, facilitate migration, and regulate T cell responses, which might play a crucial role in the progression of medulloblastoma. | [123] | ||
| Play a role in the progression of medulloblastoma | sEVs derived from control and overexpressed B7-H3 cells | B7-H3 (immunosuppressive immune check point) | This study revealed a novel role in EV production and packaging for B7-H3 that may contribute to medulloblastoma progression. | [124] | ||
| Involvement in the advancement and invasion of medulloblastoma. | sEVs derived from medulloblastoma cell line | Iron carrier proteins | This study establishes the relationship between iron metabolism and the advancement and invasion of medulloblastoma. The reduction in iron induces cell cycle arrest in the G1/S phases, resulting in the suppression of cell proliferation and the initiation of apoptosis. | [125] | ||
| Cancer EVs promote metastasis | Neuroblastoma | Pro-metastatic | sEVs derived from neuroblastoma cell line | IGF2BP1 | IGF2BP1 affects the levels of SEMA3A and SHMT2 in EVs, and this regulation plays a role in developing a pro-metastatic milieu in metastatic organs. | [126] |
| Ewing’s sarcoma | Promote tumor cell migration | sEVs derived from Ewing’s sarcoma cell line | IGF2BP3 | This data indicated that IGF2BP3 containing EVs may have a role in regulating phenotypic heterogeneity and promoting cell migration of Ewing’s sarcoma. | [127] | |
| Osteosarcoma | Promote tumor cell migration and invasive phenotype | sEVs derived from high metastatic potential cell line | Several proteins relating to signaling pathways mediated by G-protein coupled receptors | The uptake of sEVs derived from the high metastatic osteosarcoma cell line by the same cell line with low metastatic ability leads to the induction of migratory and invasive phenotypes. Proteins relating to signaling pathways mediated by G-protein coupled receptors may play a crucial role in driving metastasis of osteosarcoma. | [128] | |
| Medulloblastoma | Promote medulloblastoma metastasis through extracellular matrix signaling | sEVs derived from metastatic medulloblastoma cell line | EMMPRIN, MMP-2 | This study provides evidence for the significance of EMMPRIN and MMP-2-associated sEVs in facilitating a conducive environment that promotes medulloblastoma metastasis. | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Singhto, N.; Pongphitcha, P.; Jinawath, N.; Hongeng, S.; Chutipongtanate, S. Extracellular Vesicles for Childhood Cancer Liquid Biopsy. Cancers 2024, 16, 1681. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16091681
AMA Style
Singhto N, Pongphitcha P, Jinawath N, Hongeng S, Chutipongtanate S. Extracellular Vesicles for Childhood Cancer Liquid Biopsy. Cancers. 2024; 16(9):1681. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16091681
Chicago/Turabian StyleSinghto, Nilubon, Pongpak Pongphitcha, Natini Jinawath, Suradej Hongeng, and Somchai Chutipongtanate. 2024. "Extracellular Vesicles for Childhood Cancer Liquid Biopsy" Cancers 16, no. 9: 1681. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16091681
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.