
Benefits of Puerarin on Metabolic Syndrome and Its Associated Cardiovascular Diseases in Rats Fed a High-Fat/High-Sucrose Diet

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Hemogram Analysis
MLR = ([absolute monocyte count (AMC)])/ALC;
SII = [ANC × absolute platelet count (APC)]/ALC
2.3. Serum Biochemical Determination
2.4. Atherogenic Index (AI)
2.5. Echocardiographic Analysis
2.6. Histological Examination
2.7. Immunofluorescence Analysis
2.8. RNA Isolation and Reverse Transcription-Quantitative PCR (qRT-PCR)
2.9. Statistical Analysis
3. Results
3.1. PUE Improved Metabolic Parameters in HFHS Diet Rats

3.2. PUE Decreased the Cardiovascular Damage Indicators in HFHS Diet Rats
3.3. PUE Mitigated Cardiac Dysfunction and Structural Remodeling in HFHS Diet Rats
3.4. PUE Ameliorated Arterial Stiffness in HFHS Diet Rats
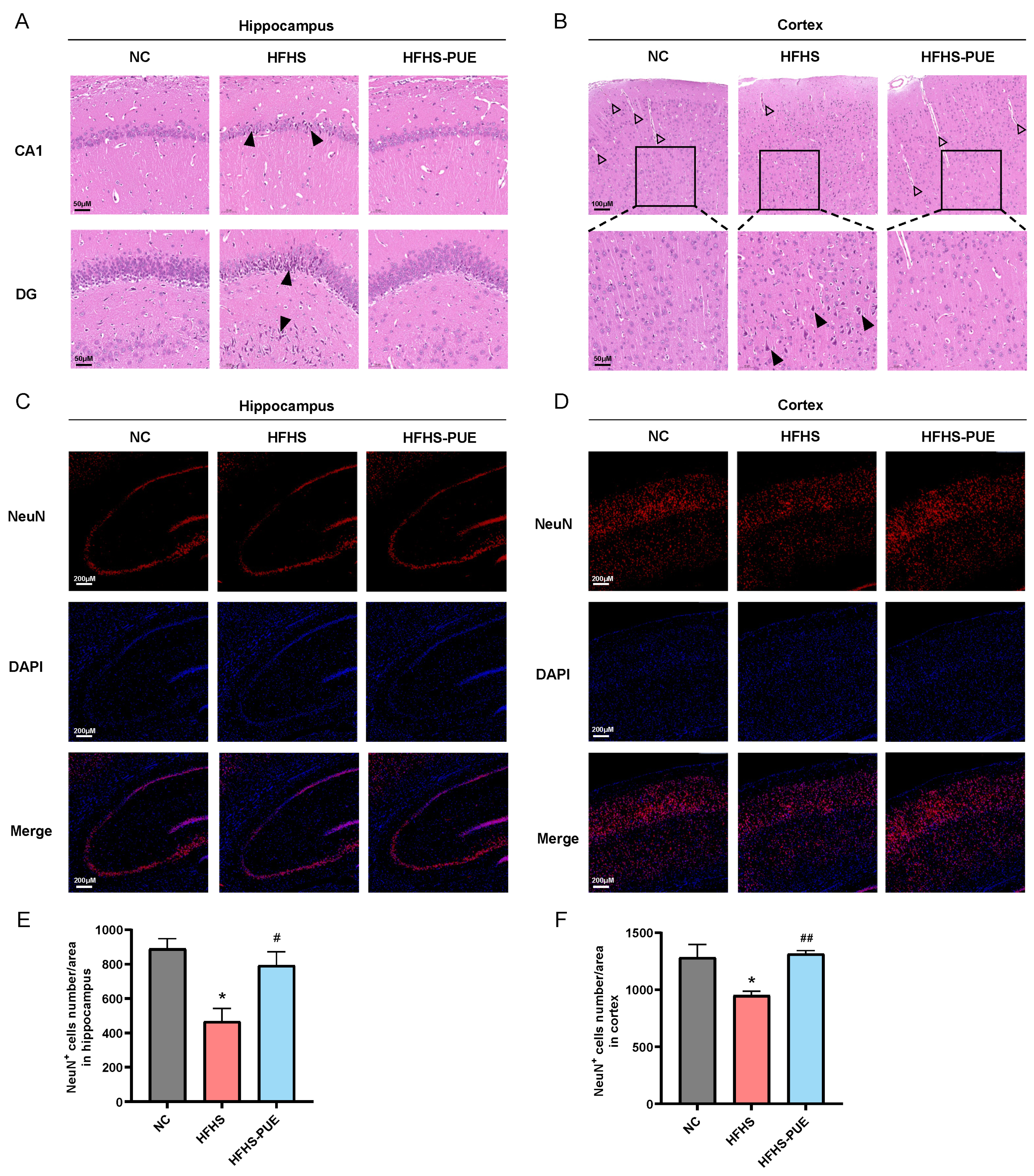
3.5. PUE Alleviated Neuronal Damage in HFHS Diet Rats
3.6. PUE Reduced the Systemic Inflammatory Indices in HFHS Diet Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Patho-physiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef]
- Gupta, A.; Gupta, V. Metabolic syndrome: What are the risks for humans? Biosci. Trends 2010, 4, 204–212. [Google Scholar]
- Purwowiyoto, S.L.; Prawara, A.S. Metabolic syndrome and heart failure: Mechanism and management. Med. Pharm. Rep. 2021, 94, 15–21. [Google Scholar] [CrossRef]
- Arcopinto, M.; Schiavo, A.; Salzano, A.; Bossone, E.; D’assante, R.; Marsico, F.; Demelo-Rodriguez, P.; Baliga, R.R.; Cittadini, A.; Marra, A.M. Metabolic Syndrome in Heart Failure: Friend or Foe? Heart Fail. Clin. 2019, 15, 349–358. [Google Scholar] [CrossRef]
- Ingelsson, E.; Arnlöv, J.; Lind, L.; Sundström, J. Metabolic syndrome and risk for heart failure in middle-aged men. Heart 2006, 92, 1409–1413. [Google Scholar] [CrossRef]
- Stehouwer, C.D.; Henry, R.M.; Ferreira, I. Arterial stiffness in diabetes and the metabolic syndrome: A pathway to cardiovascular disease. Diabetologia 2008, 51, 527–539. [Google Scholar] [CrossRef]
- Laurent, S.; Boutouyrie, P.; Asmar, R.; Gautier, I.; Laloux, B.; Guize, L.; Ducimetiere, P.; Benetos, A. Aortic stiffness is an independent predictor of all-cause and cardiovascular mortality in hypertensive patients. Hypertension 2001, 37, 1236–1241. [Google Scholar] [CrossRef]
- Cruickshank, K.; Riste, L.; Anderson, S.G.; Wright, J.S.; Dunn, G.; Gosling, R.G. Aortic pulse-wave velocity and its relationship to mortality in diabetes and glucose intolerance: An integrated index of vascular function? Circulation 2002, 106, 2085–2090. [Google Scholar] [CrossRef]
- Willum-Hansen, T.; Staessen, J.A.; Torp-Pedersen, C.; Rasmussen, S.; Thijs, L.; Ibsen, H.; Jeppesen, J. Prognostic value of aortic pulse wave velocity as index of arterial stiffness in the general population. Circulation 2006, 113, 664–670. [Google Scholar] [CrossRef]
- Mattace-Raso, F.U.; Van Der Cammen, T.J.; Hofman, A.; Van Popele, N.M.; Bos, M.L.; Schalekamp, M.A.; Asmar, R.; Reneman, R.S.; Hoeks, A.P.; Breteler, M.M.; et al. Arterial stiffness and risk of coronary heart disease and stroke: The Rotterdam Study. Circulation 2006, 113, 657–663. [Google Scholar] [CrossRef]
- Wang, D.; Bu, T.; Li, Y.; He, Y.; Yang, F.; Zou, L. Pharmacological Activity, Pharmacokinetics, and Clinical Research Progress of Puerarin. Antioxidants 2022, 11, 2121. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, L.; Wang, M. Effects of puerarin on chronic inflammation: Focus on the heart, brain, and arteries. Aging Med. 2021, 4, 317–324. [Google Scholar] [CrossRef]
- Jing, X.; Zhou, J.; Zhang, N.; Zhao, L.; Wang, S.; Zhang, L.; Zhou, F. A Review of the Effects of Puerarin on Glucose and Lipid Metabolism in Metabolic Syndrome: Mechanisms and Opportunities. Foods 2022, 11, 3941. [Google Scholar] [CrossRef]
- Liu, X.; Huang, R.; Wan, J. Puerarin: A potential natural neuroprotective agent for neurological disorders. Biomed. Pharmacother. 2023, 162, 114581. [Google Scholar] [CrossRef]
- Zhu, T.; Wang, L.; Wang, L.P.; Wan, Q. Therapeutic targets of neuroprotection and neurorestoration in ischemic stroke: Applications for natural compounds from medicinal herbs. Biomed. Pharmacother. 2022, 148, 112719. [Google Scholar] [CrossRef]
- Xu, H.; Yu, S.; Lin, C.; Dong, D.; Xiao, J.; Ye, Y.; Wang, M. Roles of flavonoids in ischemic heart disease: Cardioprotective effects and mechanisms against myocardial ischemia and reperfusion injury. Phytomedicine 2024, 126, 155409. [Google Scholar] [CrossRef]
- Qin, W.; Guo, J.; Gou, W.; Wu, S.; Guo, N.; Zhao, Y.; Hou, W. Molecular mechanisms of isoflavone puerarin against cardiovascular diseases: What we know and where we go. Chin. Herb. Med. 2022, 14, 234–243. [Google Scholar] [CrossRef]
- Zhao, Y.; Shao, W.; Zhu, Q.; Zhang, R.; Sun, T.; Wang, B.; Hu, X. Association between systemic immune-inflammation index and metabolic syndrome and its components: Results from the National Health and Nutrition Examination Survey 2011–2016. J. Transl. Med. 2023, 21, 691. [Google Scholar] [CrossRef]
- Hassan, S.; El-Twab, S.A.; Hetta, M.; Mahmoud, B. Improvement of lipid profile and antioxidant of hypercholesterolemic albino rats by polysaccharides extracted from the green alga Ulva lactuca Linnaeus. Saudi J. Biol. Sci. 2011, 18, 333–340. [Google Scholar] [CrossRef]
- Nagueh, S.F.; Smiseth, O.A.; Appleton, C.P.; Byrd, B.F., III; Dokainish, H.; Edvardsen, T.; Flachskampf, F.A.; Gillebert, T.C.; Klein, A.L.; Lancellotti, P.; et al. Recommendations for the Evaluation of Left Ventricular Diastolic Function by Echocardiography: An Update from the American Society of Echocardiography and the European Association of Cardiovascular Imaging. J. Am. Soc. Echocardiogr. 2016, 29, 277–314. [Google Scholar] [CrossRef]
- Cecelja, M.; Chowienczyk, P. Dissociation of aortic pulse wave velocity with risk factors for cardiovascular disease other than hypertension: A systematic review. Hypertension 2009, 54, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Kwok, M.K.; Leung, G.M.; Xu, L.; Tse, H.F.; Lam, T.H.; Schooling, C.M. Effect of puerarin supplementation on cardiovascular disease risk factors: A randomized, double-blind, placebo-controlled, 2-way crossover trial. Biomed. Pharmacother. 2022, 153, 113472. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Zhang, H.; Peng, C. Effects of Puerarin on the Prevention and Treatment of Cardiovascular Diseases. Front. Pharmacol. 2021, 12, 771793. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xia, L.; Song, J.; Hu, H.; Zang, N.; Yang, J.; Zou, Y.; Wang, L.; Zheng, X.; He, Q.; et al. Puerarin ameliorates metabolic dysfunction-associated fatty liver disease by inhibiting ferroptosis and inflammation. Lipids Health Dis. 2023, 22, 202. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, F.J.; Shang, L.C.; Zhang, Y.H.; Zhou, Y.; Shi, X.L. Puerarin protects against high-fat high-sucrose diet-induced non-alcoholic fatty liver disease by modulating PARP-1/PI3K/AKT signaling pathway and facilitating mitochondrial homeostasis. Phytother. Res. 2019, 33, 2347–2359. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.W.; Yang, H.K.; Jun, M.S.; Lee, B.C. Puerarin Attenuates Obesity-Induced Inflammation and Dyslipidemia by Regulating Macrophages and TNF-Alpha in Obese Mice. Biomedicines 2022, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Barua, L.; Faruque, M.; Banik, P.C.; Ali, L. Atherogenic index of plasma and its association with cardiovascular disease risk factors among postmenopausal rural women of Bangladesh. Indian. Heart J. 2019, 71, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.P.; Chan, S.W.; Chan, A.S.; Chen, S.L.; Ma, X.J.; Xu, H.X. Puerarin decreases serum total cholesterol and enhances thoracic aorta endothelial nitric oxide synthase expression in diet-induced hypercholesterolemic rats. Life Sci. 2006, 79, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Sabarinathan, M.; Ds, D.R.; Ananthi, N.; Krishnan, M. Atherogenic index of plasma, lipid accumulation and visceral adiposity in metabolic syndrome patients. Bioinformation 2022, 18, 1109–1113. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, L.; Zhang, D.; Chen, Y.; Jin, F. Puerarin alleviates coronary heart disease via suppressing inflammation in a rat model. Gene 2021, 771, 145354. [Google Scholar] [CrossRef]
- Chen, R.; Xue, J.; Xie, M. Puerarin prevents isoprenaline-induced myocardial fibrosis in mice by reduction of myocardial TGF-β1 expression. J. Nutr. Biochem. 2012, 23, 1080–1085. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.Z.; Huang, Z.; Ma, S.C.; Zen, Z.; Luo, B.; Lin, X.; Xu, X. [Study on the effect and mechanism of puerarin on the size of infarction in patients with acute myocardial infarction]. Zhongguo Zhong Xi Yi Jie He Za Zhi 2004, 24, 790–792. [Google Scholar] [PubMed]
- Xiao, L.Z.; Gao, L.J.; Ma, S.C. [Comparative study on effects of puerarin and granulocyte colony-stimulating factor in treating acute myocardial infarction]. Zhongguo Zhong Xi Yi Jie He Za Zhi 2005, 25, 210–213. [Google Scholar] [PubMed]
- Gargiulo, P.; Marsico, F.; Renga, F.; Dell’aversana, S.; Esposito, I.; Marciano, C.; Dellegrottaglie, S.; Perrone-Filardi, P.; Paolillo, S. The metabolic syndrome in heart failure: Insights to specific mechanisms. Heart Fail. Rev. 2020, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Capone, F.; Sotomayor-Flores, C.; Bode, D.; Wang, R.; Rodolico, D.; Strocchi, S.; Schiattarella, G.G. Cardiac metabolism in HFpEF: From fuel to signalling. Cardiovasc. Res. 2023, 118, 3556–3575. [Google Scholar] [CrossRef] [PubMed]
- Redfield, M.M.; Borlaug, B.A. Heart Failure with Preserved Ejection Fraction: A Review. JAMA 2023, 329, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Borlaug, B.A. Evaluation and management of heart failure with preserved ejection fraction. Nat. Rev. Cardiol. 2020, 17, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Froehlich, J.; Galis, Z.S.; Lakatta, E.G. Increased expression of matrix metalloproteinase-2 in the thickened intima of aged rats. Hypertension 1999, 33, 116–123. [Google Scholar] [CrossRef]
- Wilkinson, I.B.; Prasad, K.; Hall, I.R.; Thomas, A.; Maccallum, H.; Webb, D.J.; Frenneaux, M.P.; Cockcroft, J.R. Increased central pulse pressure and augmentation index in subjects with hypercholesterolemia. J. Am. Coll. Cardiol. 2002, 39, 1005–1011. [Google Scholar] [CrossRef]
- Mitchell, G.F.; Guo, C.Y.; Benjamin, E.J.; Larson, M.G.; Keyes, M.J.; Vita, J.A.; Vasan, R.S.; Levy, D. Cross-sectional correlates of increased aortic stiffness in the community: The Framingham Heart Study. Circulation 2007, 115, 2628–2636. [Google Scholar] [CrossRef]
- Uchida, J.; Machida, Y.; Iwai, T.; Kuwabara, N.; Kabei, K.; Naganuma, T.; Kumada, N.; Nakatani, T. Glucose intolerance is associated with increased intimal-medial thickness of the carotid artery and increased pulse-wave velocity in renal transplant recipients. Transplant. Proc. 2013, 45, 1535–1539. [Google Scholar] [CrossRef] [PubMed]
- Chirinos, J.A.; Segers, P.; Hughes, T.; Townsend, R. Large-Artery Stiffness in Health and Disease: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2019, 74, 1237–1263. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Dong, S.; Wu, Y.; He, Y.; Lu, M.; Shi, D.; Feng, N.; Yin, S.; Jiang, Y.; Zhang, A.; et al. Ameliorated biomechanical properties of carotid arteries by puerarin in spontaneously hypertensive rats. BMC Complement. Med. Ther. 2021, 21, 173. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Zhong, X.; Li, Z.; Cai, S.; Yang, P.; Ou, C.; Chen, M. Puerarin inhibits vascular calcification of uremic rats. Eur. J. Pharmacol. 2019, 855, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Xiang, D.X.; Yuan, H.Y.; Xiao, Y.; Yuan, L.Q.; Li, H.B. Puerarin attenuates calcification of vascular smooth muscle cells. Am. J. Chin. Med. 2014, 42, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Luo, Y.; Liu, T.; Zhong, X.; Yan, J.; Huang, Q.; Tao, J.; He, Q.; Guo, M.; Hu, Y. The Effect of Puerarin on Carotid Intima-media Thickness in Patients with Active Rheumatoid Arthritis: ARandomized Controlled Trial. Clin. Ther. 2018, 40, 1752–1764.e1. [Google Scholar] [CrossRef] [PubMed]
- Koivistoinen, T.; Aatola, H.; Hutri-Kähönen, N.; Juonala, M.; Viikari, J.S.; Laitinen, T.; Taittonen, L.; Lehtimäki, T.; Kööbi, T.; Raitakari, O.T.; et al. Systemic hemodynamics in young adults with the metabolic syndrome: The Cardiovascular Risk in Young Finns Study. Ann. Med. 2010, 42, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, A.; Najjar, S.S.; Orru, M.; Usala, G.; Piras, M.G.; Ferrucci, L.; Cao, A.; Schlessinger, D.; Uda, M.; Lakatta, E.G. The central arterial burden of the metabolic syndrome is similar in men and women: The SardiNIA Study. Eur. Heart J. 2010, 31, 602–613. [Google Scholar] [CrossRef]
- Lilitkarntakul, P.; Dhaun, N.; Melville, V.; Kerr, D.; Webb, D.J.; Goddard, J. Risk factors for metabolic syndrome independently predict arterial stiffness and endothelial dysfunction in patients with chronic kidney disease and minimal comorbidity. Diabetes Care 2012, 35, 1774–1780. [Google Scholar] [CrossRef]
- Więckowska-Gacek, A.; Mietelska-Porowska, A.; Wydrych, M.; Wojda, U. Western diet as a trigger of Alzheimer’s disease: From metabolic syndrome and systemic inflammation to neuroinflammation and neurodegeneration. Ageing Res. Rev. 2021, 70, 101397. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.H.; Choi, K.H.; Jang, Y.J.; Bae, S.S.; Choi, B.T.; Shin, H.K. Hypercholesterolemia accelerates amyloid β-induced cognitive deficits. Int. J. Mol. Med. 2013, 31, 577–582. [Google Scholar] [CrossRef]
- Heverin, M.; Maioli, S.; Pham, T.; Mateos, L.; Camporesi, E.; Ali, Z.; Winblad, B.; Cedazo-Minguez, A.; Björkhem, I. 27-hydroxycholesterol mediates negative effects of dietary cholesterol on cognition in mice. Behav. Brain Res. 2015, 278, 356–359. [Google Scholar] [CrossRef]
- Rutkowsky, J.M.; Lee, L.L.; Puchowicz, M.; Golub, M.S.; Befroy, D.E.; Wilson, D.W.; Anderson, S.; Cline, G.; Bini, J.; Borkowski, K.; et al. Reduced cognitive function, increased blood-brain-barrier transport and inflammatory responses, and altered brain metabolites in LDLr -/-and C57BL/6 mice fed a western diet. PLoS ONE 2018, 13, e0191909. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, L.; Zhong, F.; Zhou, K.; Lu, C.; Cheng, X.; Wang, S. Cognitive impairment in patients with heart failure: Molecular mechanism and therapy. Heart Fail. Rev. 2023, 28, 807–820. [Google Scholar] [CrossRef]
- Atti, A.R.; Valente, S.; Iodice, A.; Caramella, I.; Ferrari, B.; Albert, U.; Mandelli, L.; De Ronchi, D. Metabolic Syndrome, Mild Cognitive Impairment, and Dementia: A Meta-Analysis of Longitudinal Studies. Am. J. Geriatr. Psychiatry 2019, 27, 625–637. [Google Scholar] [CrossRef]
- Cerasuolo, M.; Di Meo, I.; Auriemma, M.C.; Trojsi, F.; Maiorino, M.I.; Cirillo, M.; Esposito, F.; Polito, R.; Colangelo, A.M.; Paolisso, G.; et al. Iron and Ferroptosis More than a Suspect: Beyond the Most Common Mechanisms of Neurodegeneration for New Therapeutic Approaches to Cognitive Decline and Dementia. Int. J. Mol. Sci. 2023, 24, 9637. [Google Scholar] [CrossRef]
- Andreone, B.J.; Larhammar, M.; Lewcock, J.W. Cell Death and Neurodegeneration. Cold Spring Harb. Perspect. Biol. 2020, 12, a036434. [Google Scholar] [CrossRef]
- Silveira Rossi, J.L.; Barbalho, S.M.; Reverete De Araujo, R.; Bechara, M.D.; Sloan, K.P.; Sloan, L.A. Metabolic syndrome and cardiovascular diseases: Going beyond traditional risk factors. Diabetes Metab. Res. Rev. 2022, 38, e3502. [Google Scholar] [CrossRef]
- Vozarova, B.; Weyer, C.; Lindsay, R.S.; Pratley, R.E.; Bogardus, C.; Tataranni, P.A. High white blood cell count is associated with a worsening of insulin sensitivity and predicts the development of type 2 diabetes. Diabetes 2002, 51, 455–461. [Google Scholar] [CrossRef]
- Akbas, E.M.; Demirtas, L.; Ozcicek, A.; Timuroglu, A.; Bakirci, E.M.; Hamur, H.; Ozcicek, F.; Turkmen, K. Association of epicardial adipose tissue, neutrophil-to-lymphocyte ratio and platelet-to-lymphocyte ratio with diabetic nephropathy. Int. J. Clin. Exp. Med. 2014, 7, 1794–1801. [Google Scholar]
- Gijsberts, C.M.; Ellenbroek, G.; Ten Berg, M.J.; Huisman, A.; Van Solinge, W.W.; Lam, C.S.; Asselbergs, F.W.; Den Ruijter, H.M.; Pasterkamp, G.; Hoefer, I.E.; et al. Effect of Monocyte-to-Lymphocyte Ratio on Heart Failure Characteristics and Hospitalizations in a Coronary Angiography Cohort. Am. J. Cardiol. 2017, 120, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ma, Y.; Geng, X.B.; Tan, Z.; Wang, J.H.; Cui, C.; Wang, H.L.; Shang, X.M. Platelet-to-lymphocyte ratio relates to poor prognosis in elderly patients with acute myocardial infarction. Aging Clin. Exp. Res. 2021, 33, 619–624. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Y.; Yang, Y.; Jiang, S.; Liu, C.; Han, Y.; Jiang, J.; Wang, Y. Benefits of Puerarin on Metabolic Syndrome and Its Associated Cardiovascular Diseases in Rats Fed a High-Fat/High-Sucrose Diet. Nutrients 2024, 16, 1273. https://0-doi-org.brum.beds.ac.uk/10.3390/nu16091273
Mu Y, Yang Y, Jiang S, Liu C, Han Y, Jiang J, Wang Y. Benefits of Puerarin on Metabolic Syndrome and Its Associated Cardiovascular Diseases in Rats Fed a High-Fat/High-Sucrose Diet. Nutrients. 2024; 16(9):1273. https://0-doi-org.brum.beds.ac.uk/10.3390/nu16091273
Chicago/Turabian StyleMu, Yu, Yalin Yang, Shuang Jiang, Chilu Liu, Yanxing Han, Jiandong Jiang, and Yuhong Wang. 2024. "Benefits of Puerarin on Metabolic Syndrome and Its Associated Cardiovascular Diseases in Rats Fed a High-Fat/High-Sucrose Diet" Nutrients 16, no. 9: 1273. https://0-doi-org.brum.beds.ac.uk/10.3390/nu16091273