
Can Arbuscular Mycorrhizal Fungi Enhance Crop Productivity and Quality in Hydroponics? A Meta-Analysis


1
Department of Horticulture and Crop Science, School of Agriculture, The University of Jordan, Amman 11942, Jordan
2
Department of Plant Protection, School of Agriculture, The University of Jordan, Amman 11942, Jordan
*
Author to whom correspondence should be addressed.
Sustainability 2024, 16(9), 3662; https://0-doi-org.brum.beds.ac.uk/10.3390/su16093662
Submission received: 22 February 2024
/
Revised: 14 April 2024
/
Accepted: 20 April 2024
/
Published: 26 April 2024
(This article belongs to the Special Issue Sustainable Agriculture for Crop Cultivation)
Abstract
:Finding environmentally friendly solutions for crop growth and productivity has been gaining more attention recently. This meta-analysis aims to understand the combined application of arbuscular mycorrhizal fungi (AMF) and hydroponic systems compared to AMF in conventional (soil) systems. The analysis of up-to-date studies revealed that the root colonization, calculated as the proportion of colonized root segments relative to the total root length, by AMF in conventional (soil-based) culture exceeded hydroponic (or soilless) culture systems by 16.8%. The mean root colonization by AMF was determined to be 52.3% in hydroponic systems and 61.1% in conventional systems. Within hydroponic systems, the root colonization ranged from 2% to 20% after 10 days of inoculation, and notably, it exceeded 50% after 30 to 65 days, depending on the growing substrate and species. Under hydroponics, AMF application had a higher (compared to none-inoculated) positive effect on crop biomass and yield than fruit and leaf quality (antioxidants, phenols, and sugars) as well as leaf nutrients. However, AMF do not always have the potential to improve crop growth, quality and productivity in hydroponics. Among the studies analyzed in this review, approximately 34% (no effect: 29%; negative: 5%) reported no discernible positive effect on biomass or yield, 37% (no effect: 16%; negative: 21%) on fruit or leaf quality, and 60% (no effect: 47%; negative: 13%) on nutrient levels within plant tissues. To improve the performance of AMF in hydroponic systems, the meta-analysis recommended maintaining phosphorus levels in the nutrient solution within the range of 0.15 to 15.5 mg L−1 as elevated levels (40–75 mg L−1) were found to significantly reduce AMF colonization. Additionally, it was observed that certain hydroponic techniques, such as the presence of air bubbles generated by air pumps in floating hydroponic systems (Deep Flow technique) and continuous circulation of the nutrient solution (Ebb and Flow systems), may create dynamic conditions that could potentially hinder the introduction of AMF spores into hydroponic systems and potentially compromise the integrity of the spores and hyphae.
1. Introduction
Climate change, the growing human population, soil nutrient depletion, water scarcity, the excessive utilization of fossil fuels, biodiversity loss, the emergence of superweeds, and the resistance of insects and phytopathogens collectively constitute a group of factors that hinder optimal production of crops and threat food security [1]. Soilless culture represents an advanced horticultural method that facilitates the provision of precisely calibrated nutrient levels to plants within controlled settings, commonly within greenhouse environments, as elucidated by Tzortzakis et al. [2]. When soilless cultivation is mentioned, it is mainly referred to the techniques of hydroponics, which come from two Greek words ‘hydro’ meaning water and ‘ponos’ meaning labor [3,4]. Hydroponics is, therefore, the technique of growing plants in a soilless condition with inert media (coir, volcanic tuff, perlite, etc.) or with their roots immersed in a nutrient solution without the use of any substrate [2,3]. The global market of this culture system is valued at $9.5 billion in 2020 and is anticipated to reach $17.9 billion by the year 2026 [5]. Among the water culture techniques, float hydroponics is adopted for the cultivation of leafy-green crops, such as lettuce, while substrate-based (solid) culture is commercially used for fruiting vegetables like tomatoes and peppers [6]. The primary advantage of hydroponics is the separation of plant growth from the constraints typically associated with soil, including soil-borne diseases, non-arable soil, soil salinity, poor physical properties, low temperature, and limited nutrient availability [2]. The plant microclimate, including pH, nutrients, and moisture content can be easily managed in hydroponic systems, thereby having a positive influence on growth and overall productivity [7].
Soil contains abundant beneficial microorganisms that play crucial roles in supporting plant nutrition, generating phytohormones, managing phytopathogens, and enhancing soil structure [8]. In contrast, hydroponic culture systems typically lack these advantageous microorganisms unless deliberately introduced into the system [8]. Arbuscular mycorrhizal fungi (AMF) have long been recognized in agriculture for their dependable and efficient symbiotic attributes towards a wide range of important crops [9]. AMF are a group of beneficial fungi that form symbiotic associations with the roots of most land plants. They belong to the Glomeromycota phylum and are characterized by their ability to form arbuscules and vesicles within plant root cells. These specialized structures facilitate the exchange of nutrients between the fungus and the plant host, with the fungus providing essential nutrients such as phosphorus and nitrogen to the plant in exchange for carbon compounds. Common species of AMF include Rhizophagus irregularis (formerly Glomus intraradices), Funneliformis mosseae (formerly Glomus mosseae), and Claroideoglomus etunicatum (formerly Glomus etunicatum) [10]. AMF can be applied as inoculants to seeds, roots, or growing media to establish symbiotic associations with plants and promote their overall health and productivity. AMF is being strongly promoted as a solution to the challenges caused by current unsustainable agricultural practices [9]. These fungi possess the ability to enhance crop growth and yield, mitigate the negative impacts of abiotic stressors such as drought, heat, and salinity, as well as biotic stressors like pests and diseases [10]. Advances in modern techniques have enabled us to investigate multiple AMF domains, such as physiology, function, and community associations of mycorrhizal fungi [11]. Worldwide, a diverse array of businesses, spanning from small to large enterprises and start-ups, actively participate in the production and distribution of AMF formulations or consortia products that encompass advantageous soil bacteria [12]. Over the past few decades, a substantial number of patents, exceeding 40 in total, have been generated, specifically pertaining to AMF. These patents encompass a broad spectrum of aspects related to AMF, including their advantageous attributes, techniques for inoculum preparation, formulation methodologies, and diverse application approaches [13,14,15]. In 2019, the global market for AMF biofertilizer reached $268.8 million, with a projected increase of 131.25% expected by 2025, bringing it to an estimated $621.6 million [16]. Consequently, AMF stands as a prominent alternative technology, holding great promise forwards sustainable agriculture production amid the global food crisis and the unpredictable challenges posed by shifting climate conditions [1].
The use of AMF approach in hydroponic-grown crops offers a promising approach to achieve both environmental sustainability and improved crop quality [17]. The combined use of bacteria and mycorrhiza bio-fertilizers in a soilless system successfully reduced mineral fertilizers by 20% [18]. Furthermore, when 80% of mineral fertilizers are used in combination with a mixture of mycorrhiza and bacteria bio-fertilizers, the concentration of mineral nutrients (N, P, K, Ca, Mg, Fe, Mn, Zn, Cu) in the leaves of pepper (Capsicum annuum L.) plants were higher than that of the control (100% mineral fertilizer without bio-fertilizers) [18]. However, the use of AMF in hydroponic culture is controversial. The process of inoculating AMF is labor intensive [14]; for example, plants need to be cultured in sand and then transferred to the common nutrient solution. Moreover, in hydroponic culture, the system directly supplies all the necessary nutrients rendering the need for AMF to assist in nutrient uptake is quite luxurious [19]. In this context, all nutrients are provided in ionic or readily accessible forms, eliminating the necessity for root expansion in search of water and nutrients in hydroponics, as these resources are abundantly available [19]. Nevertheless, several studies indicated that the application of AMF in soilless or hydroponic culture has the potential to promote growth and productivity [10,20,21]. Hydroponic methods such as nutrient film technique (NFT) provide AMF with adequate air and nutrient solution without disturbing hyphal development [22]. Soilless (substrate, sawdust) grown tomato (Solanum lycopersicum L.) inoculated with Funneliformis monosporum (Glomus monosporum), F. mosseae (G. mosseae) had higher plant length, fruit yield and fruit number compared to the control [10]. Additionally, Fusarium oxysporum infection was significantly lower in the root inoculated with AMF compared to untreated control plants. Similarly, tomato seedlings inoculated with Entrophospora etunicata (G. etunicatum) and Rhizophagus intraradices (G. intraradices) grown in soilless culture (vermiculite) for six weeks exhibited increased shoot fresh weight, shoot-to-root ratio, root biomass, and relative growth rate with a root colonization of 23.3% [23]. While the beneficial relationship between plants and AMF has been extensively studied for several terrestrial plant species, its exploration in aquatic environments such as hydroponic culture and emergent aquatic species remains relatively limited.
The objective of this review is to critically analyze and expand the knowledge of the utilization of AMF in hydroponic systems compared to soil-based culture with the goal of elucidating the primary factors influencing the AMF-hydroponic association to promote crop growth and productivity. One novelty of this review is the assessment of substantial, up-to-date available research results exploring the effect of AMF inoculation under hydroponic system culture. This offers an overview of the effectiveness associated with the combined utilization of AMF in conjunction with hydroponic systems for crop production, contrasting it with either hydroponic or AMF-supported soil cultivation. Such insights aim to stimulate further research efforts and the development of commercial applications in the future. Moreover, we discussed why the crops did not consistently exhibit a positive response to AMF inoculation in hydroponics and the possible solutions to enhance their activity under these soilless cultivation systems.
2. AMF and Hydroponics Culture
The growing interest in the applications and associated benefits of AMF worldwide is evident through the abundant scientific literature, comprising research and review articles. Data mining from scientific database using ‘mycorrhizal’ as the search term showed that the total published research was 20,544 publications in the Springer Nature database, 3378 in Taylor and Francis, 1066 in MDPI and 17,621 in ScienceDirect® over the last two decades (Figure 1). Arbuscular mycorrhizae which are found in more than 80% of terrestrial plants constitute the most prevalent form of symbiosis on Earth [24]. These beneficial fungi, belonging to the Glomeromycota sub-phylum within the Fungi kingdom, form a symbiotic relationship known as arbuscular mycorrhiza (AM) with the roots of terrestrial plants, including several economic crops [25]. Within this relationship, AMF enhances nutrient uptake by plant roots, particularly phosphate, while receiving carbohydrates and lipids from the host plant in return [24]. AMF are obligate biotrophs and extremely depend on their host plant to get organic carbon, while plants benefit from essential soil-derived nutrients (such as phosphorus (P), nitrogen (N), and zinc (Zn) provided by the fungi) and improved water relations [26,27,28]. Obligate biotrophic fungi like AMF have developed complex mechanisms for accessing plant roots and benefiting from their resources [28]. The transport of nutrients by AMF occurs through tree-shaped structures called arbuscules [29]. The root system of plants has fewer competing microbes compared to the surrounding root rhizosphere. Additionally, it benefits from a consistent and plentiful supply of resources over time and space. When this space is filled, obligate biotrophs become highly reliant on the living host to the extent that they lose their ability to grow on non-living material [28]. During the symbiotic process, AMF is typically regulated by effector proteins, which are messenger proteins released by AMF to maximize compatibility with the host plant and reduce the probability of attack by disease-resistance proteins [29]. Additionally, AMF releases biochemical compounds such as glycoprotein (ex. Glomalin), which can enhance soil physical and chemical properties, especially soil fertility [30].
Mycorrhizal associations enhance plants’ tolerance against biotic stressors like diseases and insect pests, as well as abiotic stressors such as drought and salinity [30]. AMF is known to regulate the translocation of heavy metals (e.g., Cadmium) in the root system and therefore, AMF is an essential consideration when cultivating plants in polluted soils [31]. AMF provides an eco-friendly approach to a sustainable environment [18]. The symbiotic relationship between AMF and plants plays a significant role in ecosystem management and agricultural practices [32]. The synergistic interaction between AMF and rhizobia can stimulate enhanced root colonization, thereby further promoting plant growth and N acquisition [33]. In white clover (Trifolium repens), the inoculation of both AMF (Paraglomus occultum) and rhizobium (Rhizobium trifolii) showed positive effects on leaf and root biomass production, and plant height, and taproot length within 15–20 days of inoculation [33].
Soilless cultivation encompasses various plant growth systems that replace traditional soil with porous substrates or nutrient solutions, such as the NFT [2]. Employing AMF in hydroponic cultivation represents a promising and environmentally friendly approach to increasing crop yields, improving quality, increase antioxidant compounds all while reducing reliance on chemical fertilizers [8]. Despite the significant volume of research studies focused on hydroponic culture or AMF (e.g., Springer–Nature, hydroponics/soilless 13,307; AMF/mycorrhiza 26,500), the total number of publications specifically dedicated to the use of AMF in hydroponics (soilless) culture remains limited. Online searches using ‘AMF/mycorrhiza × hydroponic/soilless’ as search term within the period between 2002–2023 (2002–2006 + 2007–2011 + 2012–2016 + 2017–2023) yielded 1240 publications in Springer Nature database, 200 in Taylor and Francis, 61 in PubMed® and 848 in ScienceDirect® (Figure 1). The incorporation of AMF into hydroponic systems presents challenges stemming from cost and complexity, as growers may face additional expenses and logistical difficulties in sourcing AMF inoculants, optimizing inoculation methods, and managing the symbiotic relationship between fungi and plants in a soilless environment. Furthermore, limited research and awareness about the potential benefits of AMF in hydroponics contribute to their underutilization, with growers often prioritizing factors such as nutrient management and disease control over the integration of AMF. The relatively low popularity of AMF in hydroponics (Figure 1), as evidenced by the scarcity of recent publications, is influenced by various factors. Traditional hydroponic systems, designed to deliver optimized nutrient solutions directly to plants, diminish the perceived necessity for root symbionts like AMF to scavenge for nutrients. Moreover, the historical focus of AMF research on soil-based agricultural systems diverts attention from their potential advantages in hydroponics. Challenges related to the application of AMF inoculants, coupled with skepticism about their efficacy and cost-effectiveness, may further discourage exploration of their use in hydroponic setups. Despite these barriers, growing interest in sustainable agriculture and soil health may catalyze renewed exploration and adoption of AMF in hydroponic systems in the future [34].
3. Does the Plant Consistently Exhibit a Positive Response to AMF Inoculation in Hydroponic Systems?
The metadata analysis results unveiled noteworthy differences in AMF root colonization over time between conventional agriculture (soil) and hydroponic cultivation systems (Figure 2). On average, the root colonization percentage by AMF was 16.8% higher in soil-based systems compared to hydroponic systems. Specifically, the mean root colonization (%) by AMF was approximately 52.3% in hydroponic systems and 61.1% in soil-based systems. In a study by Robinson Boyer et al. [35], it was observed that the colonization of strawberry roots by AMF (R. irregularis) was reduced when grown in soilless substrates. One possibility is that the absence of soil in soilless substrates alters the microenvironment and conditions required for AMF colonization. Furthermore, the physical characteristics of AMF colonization on strawberry roots exhibited disparities between soilless and soil-based cultivation. In hydroponically grown plants (using coir as a substrate), the colonization structure appeared to be more compact and smaller in size [35]. In another study by Hung and Sylvia [36], the germination rate of G. etunicatum spores in aeroponic cultures was found to be significantly lower than that observed in soil, with rates of 46% and 64%, respectively, after a 2-week incubation period at 28 °C. However, it’s worth noting that this difference did not affect the inoculum’s infectivity potential. The water regime was identified as a significant factor influencing root colonization. For instance, in the case of Panicum hemitomon and Leersia hexandra, root colonization by AMF was diminished, and the plants exhibited reduced size in waterlogged soil conditions [37].
The use of AMF when growing plants in a hydroponic system is controversial. There is conflicting evidence of the feasibility of such use (Figure 3). Based on the comprehensive examination of studies within this review, it has been demonstrated that approximately 34% of these studies reported either a negative outcome or no significant effect of AMF on biomass or yield; around 37% of the studies found no substantial impact or negative effects on the quality of fruits or leaves; and in the case of nutrient levels within plant tissues, approximately 60% of the studies revealed no or negative effect when AMF were employed within hydroponic systems (Figure 3). Recent investigations focused on the application of AMF in organic soilless vegetable production unveiled that Glomus spp. did not demonstrate a significant association with improved physiological parameters (e.g., photosynthesis, stomatal conductance, transpiration) or yield for eggplant (Solanum melongena L.) [103]. In another study by Hawkins and George [53], root O2 and CO2 fluxes were measured in hydroponically grown AMF and non-AMF Triticum aestivum. Seedlings were cultivated with 4 mM N and 10 mM P. The total root colonization reached 48%, with 18% attributable to G. mosseae. While the root O2 uptake rate remained consistent between the two groups, CO2 release was higher in AMF-associated wheat compared to non-mycorrhizal wheat. However, there were no discernible differences in biomass and nutrient concentration (specifically N and P) between the mycorrhizal and non-mycorrhizal wheat [53]. In the context of strawberry, the inoculation with bacteria and AMF (B. velezensis and G. intraradices) resulted in an increased chlorophyll content index but decreased certain fruit quality components, including total soluble solids, titratable acidity, and pH [46]. Furthermore, when lettuce seedlings were inoculated with AMF (G. mosseae and G. intraradices) and grown in soilless substrates such as rockwool or coconut coir, the leaf concentrations of Cu, Mn, and Zn were lower compared to non-inoculated specimens. Additionally, no significant differences were observed in terms of yield and macronutrient content between AMF-inoculated and non-inoculated lettuce [55]. In the case of cucumber (C. sativus) grown in a modified Johnson nutrient solution and inoculated with AMF (G. mosseae), certain findings were noted such as lower plant growth and shoot phenol content, higher shoot SPAD values, photosynthesis rates, transpiration rates, mesophyll conductance, root exudate phenol content, and antioxidant activity [104].
Several explanations have been suggested to clarify why plant response to AMF inoculation is not consistently positive under hydroponic systems [14,29,64]. In hydroponics, the systems provide the required mineral nutrients and water to the roots and therefore there is no need to stimulate the AMF symbiosis [29]. The addition of macro- and micronutrient fertilizers in hydroponics can potentially lead to an increase in the soilless nutrient solution electrical conductivity. The electrical conductivity of full-strength commercial nutrient solution’s ranges from 1.2 to 2.5 dS m−1, and in closed-hydroponic systems, salinity levels can potentially increase. Irrigation with saline water containing 4500 ppm of NaCl, MgCl2, and CaCl2 (in a 3:1:1 ratio by weight) has been shown to reduce AMF colonization [82]. Under salinity conditions, AMF reduced leaf Na+, Ca2+ and sucrose concentration while increasing the ratios of K+, K+/Na+, Ca2+/Na+ and Mg2+/Na+. Additionally, AMF can enhance glucose, fructose, and proline concentrations in trifoliate orange (Poncirus trifoliata) seedlings [66]. The spread of pathogens such as root rot have become a continuous threat to crop production in commercial soilless systems [7]. This can increase the risk of attack by root pathogens because the inoculum is distributed by recirculating nutrient solution [116]. In comparison, AMF-inoculated strawberry plants grown in soil exhibited higher tolerance to soil-borne pathogens and water stress than those grown in soilless (coir) substrate [35].
4. Potential Use of AMF in Hydroponic Culture
In hydroponic systems where nutrient levels are already optimized, mycorrhizal fungi play crucial roles beyond nutrient extraction. They facilitate enhanced water uptake, improve nutrient absorption efficiency, and promote stress tolerance in plants, thereby contributing to overall plant health and productivity [8,19]. Additionally, mycorrhizal symbiosis fosters robust root growth and structure, supports beneficial microbial communities, and ultimately enhances plant growth and yield [23,30]. While nutrient extraction may not be their primary function, mycorrhizal fungi offer multifaceted benefits that make them valuable additions to hydroponic systems, aiding in the creation of resilient, healthy crops.
Various hydroponic techniques have been proposed for integrating AMF, including floating, nutrient film technique (NFT), aeroponics, and Ebb and flow systems. In the floating system, plant roots are suspended in a nutrient solution-filled bed or reservoir, with an air pump ensuring adequate oxygen supply to prevent root oxygen deprivation. NFT systems involve suspending plants above a continuous film of nutrient solution. In aeroponics, plants are suspended in the air, with AMF-inoculated roots exposed to a mist of nutrient solution [14]. These enclosed frameworks enhance the aeration surrounding the root system and could play a significant role in improving the performance of AMF. For example, Bahia grass (Paspalum notatum) and sweet potato (Ipomoea batatas) pre-inoculated with G. deserticola, G. etunicatum, and G. intraradices showed significant root colonization ranging from 28–45%, after 12 weeks in aeroponic cultures [36]. The spores count ranged from 4.0 spores per cm of colonized root for G. etunicatum to 51 spores per cm for G. intraradices [36]. In another study involving Hibiscus sabdariffa, inoculation with AMF species such as Glomus sp., Gigaspora sp. and Scutellospora sp. under soilless culture resulted in the proliferation of fungal spores in the growing medium, ranging from 55 to 61 spores/10 g substrate. Root colonization levels ranged from 59% to 64%, and root volume doubled compared to the control [38].
The combined utilization of AMF and hydroponic culture has revealed a potential positive effect on the growth and productivity of several crops, as summarized in Table 1. In a floating hydroponic culture system, AMF contributed to the increased formation of lateral branches in the basil plants (Ocimum basilicum L.) without causing the thickening of their stems. In addition, AMF enhanced nutrient uptake including N, P, K, Ca, Mg, Fe, Mn, Zn, Cu [8]. When considering the total harvest data, AMF increased basil yield by 13.94% compared to 50% control (the response of the bio-fertilizers where mineral fertilizers are reduced by 50%) [8]. Under soilless culture using coir substrate, AMF inoculation led to an increase in the size and quantity of class 1 strawberry fruit, especially under low N input condition [35]. In a perlite-based hydroponic system, the mycorrhizal inoculation of pepper plants hydroponically grown showed an increase in plant dry weight by 17% to 36% compared to un-inoculated plants with two AMF species (G. caledonium and G. clarum) used [39]. In a close hydroponic system, corn seedlings colonized by AMF exhibited a significantly higher P uptake rate (from Hoagland solution reservoir at 9 and 21 h) compared to the non-colonized plantlets [117]. Under soilless culture utilizing peat moss and perlite mix (1:1 v/v), AMF-inoculated pepper plants displayed higher photosynthesis, stomatal conductance, leaf chlorophyll, leaf nutrient (N, P, K), leaf area, and fruit production compared to the non-AMF plants [118]. Saffron (Crocus sativus) cultivation commonly occurs in soil, with spice yield ranging from 0.15 to 1.5 g m−2, depending on cultural practices [105]. In soilless systems, saffron can benefit from essential nutrients without competition with pathogens or abiotic stresses associated with nutrient-soil interaction. These systems provide an optimal environment for saffron growth, allowing for precise control over nutrient availability and minimizing the risk of soil-borne diseases. Additionally, soilless cultivation methods reduce the likelihood of nutrient imbalances or deficiencies, promoting healthier plant development and potentially higher yields. Although saffron corms inoculated with AMF (R. intraradices) and produced in hydroponic system exhibited spice yields like those in soil, they showed higher levels of polyphenols, anthocyanins, vitamin C, and elevated antioxidant activity [105].
In terms of its environmental impact, the use of AMF can lead to a reduction in the necessity for chemical fertilizers and pesticides by improving nutrient uptake and enhancing plant resistance to diseases [8,117,118,119]. For example, cultivating watermelon seedlings inoculated with AMF in a hydroponic system, using 80% full-strength nutrient solution, resulted in a 49.5% increase in total yield (12.4 vs. 8.3 kg m−2) compared to the control [106]. Additionally, the P content in AMF-inoculated melons increased by 80% in the 80% full-strength nutrient solution compared to the full-strength control [106]. In the case of hydroponically grown baby spinach (Spinacia oleracea L.), where AMF was used in combination with 50% mineral fertilizers, leaf yields were 20% lower than those achieved with 100% mineral fertilizers [17]. However, AMF significantly improved the quality of the internal leaves, including enhancing total phenolic content, vitamin C levels, and total soluble solids [17]. Inoculating tomatoes with AMF (F. mosseae) grown in a nutritional solution within a coconut coir medium resulted in increased root biomass, total root length, surface area, and volume compared to the control [120]. Moreover, the expression levels of N transporters such as LeAMT1.1, LeAMT1.2, and LeNRT2.3 were significantly up regulated by AMF inoculation [120]. Mycorrhizal colonization in soilless grown cut flower snapdragons (Antirrhinum majus L.) significantly increased the vase life of flowers by reducing ethylene production [40]. Although AMF did not lead to an increase in the number of flowers per spray or the concentration of flower P, its broader influence on host ethylene production indicates that this microbial association might offer a viable alternative to chemical ethylene inhibitors like silver thiosulfate and silver nitrate, which are known for their high toxicity [40,121,122].
5. The Percentage of AMF Root Colonization over Time within Hydroponic Systems
Figure 4 illustrates the progressive colonization of roots by AMF over time across various substrates, including nutrient solutions, soil, sand, vermiculite, and peatmoss. Despite the extensive research related to AMF (as shown in Figure 1), relatively few studies have specifically focused on the dynamics of root colonization over time (Figure 4). These studies have revealed that after an initial 10 days following inoculation, the percentage of root colonization by AMF typically falls within the range of 2% to 20%, depending on the specific growing substrate. Notably, in soilless substrates, root colonization by AMF consistently exceeded 50% after 30 to 65 days, with variations influenced by both the growing substrate and the plant species under investigation. In the early stages of transplanting Linum usitatissimum in the hydroponic floating system (inoculation was done before transplanting), the degree of AMF colonization in the nutrient solution was slower compared to the sand culture. However, over time, the colonization percentage gradually increased, eventually reaching levels like those observed in the sand culture, at approximately 90%, after 8 weeks [41]. This could be attributed to the rapid growth of the root system in nutrient solution.
For Sudan grass (Sorghum sudanese), roots inoculated with AMF (G. intraradices) and grown using a hydroponic technique, specifically an ultrasonic nebulizer aeroponic system, the root colonization percentage reached a substantial 80% [123]. Moreover, the total number of viable AMF propagules per gram of inoculum increased to 175,000 after 12 weeks of pre-inoculation [123]. In the case of Chile pepper (Capsicum annuum L. cv. San Luis) plantlets grown in a soilless system composed of peatmoss and perlite (in a 1:1 v/v ratio), AMF colonization proceeded gradually, reaching approximately 50% within 45 days. This colonization significantly contributed to enhancements in photosynthesis, stomatal conductance, and water relations adjustments. These improvements facilitated rapid recovery during acclimatization and subsequently resulted in greater growth during the post-acclimatization period [118]. Furthermore, in the context of white clover (T. repens), following inoculation with AMF (P. occultum), the roots exhibited an increase in hydrogen peroxide (H2O2), which serves as a signaling molecule for plant defense against microbial infestations. This increase occurred between day 5 and day 10 after AMF inoculation [33].

6. How to Enhance AMF Activity in Hydroponic Systems?
6.1. Phosphorus Level in Nutrient Solution
The composition of the nutrient solution in hydroponic systems plays a significant role in influencing root colonization and sporulation of AMF [127]. Plants could control the AMF colonization extent based on their nutritional requirements, possibly as a mechanism to optimize energy resources and minimize the carbon input cost generated with AMF symbiosis [29]. The symbiosis relationship, specifically the delivery of phosphate, is one of the main determinants of this relationship [29]. When phosphate availability in root rhizosphere is high, the colonization and the symbiotic phosphate uptake pathway are inhibited. Conversely, when phosphate levels are low, the process shifts from the direct rhizodermal-uptake pathway to the symbiotic-uptake pathway [29]. For instance, when P. notatum was inoculated with AMF (G. intraradices, Acaulospora longula, Scutellospora heterogama, G. margarita) and grown in soilless culture (sandy media) of different Hoagland nutrient solutions, the sporulation of AMF varied significantly. The addition of NH4NO3 reduced both colonization and sporulation of AMF (G. intraradices) compared to nutrient solutions without P. Hoagland nutrient solutions without P increased sporulation in the growing media and root colonization of all AMF compared to those produced by a tap water or full-strength nutrient solutions containing the recommended P levels [127]. In this review, the meta-analysis revealed that in hydroponic systems, the recommended P level in nutrient solutions ranged from 0.15 to 15 mg L−1 and higher levels (40–75 mg L−1) could have a negative effect on AM fungus colonization and activity (Figure 5).
When there is an excess of phosphorus (P) in the root rhizosphere, the formation of intact arbuscules may decrease, and instead, vesicles may develop [128]. Since phosphorus availability in the growing substrates is restricted, plants have evolved various mechanisms to boost their absorptive surfaces, such as root elongation and branching [130]. In a study conducted by Vejsadová et al. [131], the influence of P-solubilizing bacterial isolates and a collection of bacterial strains on the development of native G. fasciculatum colonization in corn roots was investigated. Under hydroponic conditions, the P-solubilizing isolate F27 significantly increased the number of arbuscules in roots and P concentration in the shoots (by 30–35%), compared to plants with AMF alone [131].
Arbuscular mycorrhizal (AM) symbiosis offers a promising strategy for enhancing plant P acquisition. However, when the substrate contains high P levels, it can reduce fungal colonization and impede mycorrhizal P uptake [128]. In a study conducted by Valentine and Kleinert [129], the impact of increased P availability on the respiration of AMF (G. mosseae) in tomato roots grown hydroponically for 72 h was investigated. Initially, seedlings were cultivated in a low P environment (2 μM, approximately 0.06 mg L−1) for three weeks. Subsequently, the same inoculated transplants were exposed to elevated P doses (2 mM P, around 62 mg L−1) for 72 h. The additional P led to a reduction in AMF activity, root CO2 and O2 fluxes, as well as the concentration of organic acids produced from root-zone CO2 assimilation [129]. This short-term increase in low P conditions (72 h) negatively impacted the activity of the fungal symbiont, AMF root respiration, and the metabolism of root-zone CO2-derived organic acids [129]. A study involving Lythrum salicaria grown in soilless sand culture at various P concentrations (0, 0.04, 0.40, 4.0, and 17 mg L−1 nutrient solution) for 49 days, with or without AMF inoculation, revealed that AMF colonization occurred in plants at the 0.04, 0.40, and 4.0 mg-P L−1 concentrations, but there was no colonization in plants grown at the 17 mg-P L−1 nutrient solution [65]. Another experiment with wheat (T. aestivum) inoculated with G. mosseae and grown in nutrient solution with different P levels showed that the modified Long Ashton solution (10 µM P, 0.3 mg L−1) resulted in higher root dry weight and viable cultures of mycorrhizal colonization compared to the modified Knop plus Hoagland medium (900 µM P, approximately 28 mg L−1) [93]. Interestingly, mycorrhization by Glomus occurred in bean roots colonized by rhizobia when the P concentration in the hydro-aeroponic solution was ranged from 75 µM (~2.3 mg L−1) to 250 μM (~7.7 mg L−1) [64]. For the 75 μM P treatment, the roots exhibited colonization percentages of 14% for hyphae, 7% for arbuscules, and 2.66% for vesicles. In the case of the 250 μM P treatment, the colonization percentages were 32% for hyphae, 15% for arbuscules, and 8% for vesicles [64].
6.2. Static vs. Dynamic Hydroponic System
When the fungus spores are located at a distance from the root, plant-secreted flavonoids encourage hyphal elongation and stimulate pre-symbiotic fungal growth [132]. During this stage (presymbiosis), both plant and fungus communicate via oligosaccharides and butenolides [132]. After that, AM fungus becomes close to the root and a high degree of hyphal branching is mediated by plant strigolactones. Perception of cutin monomers by germinated spores triggers hyphopodium development on the root surface. After that, the tips of growing hyphae attach to the root epidermis hyphopodia, and the hyphae penetrate the root epidermis and colonize the cortex [26,133]. In addition, germinated fungal spores of AM fungi secrete sulfated and non-sulfated lipochitooligosaccharide that trigger lateral root formation [133]. The cultivation of AMF independently of the host plant is not possible [53]. There is a unique process that occur between AMF and the plant that might require a steady (static) state around the root rhizosphere. Permanent nutrient solution circulation in NFT, aerobic hydroponics as well as the bursting of air bubbles (air pump) in floating systems create a dynamic state that could reduce the possibility of injecting AMF spores to the hydroponic system and might damage the spores and/or hyphae [22,53]. In floating systems where the nutrient solution remains static, the bursting of air bubbles from air pumps may damage the delicate extraradical hyphae or restrict their formation [14].
Nurbaity et al. [59] assessed the use of Deep Flow (where plant roots are completely submerged in the nutrient solution) and Ebb and Flow (where roots are periodically flooded with nutrient solution and then drained) to optimize the production of mycorrhizal fungi; with AMF-inoculated Sorghum bicolor was used as host plants. They found that Ebb and Flow resulted in higher plant biomass, root mycorrhizal colonization (79%), and spores’ number (962 spore/10 g medium) when compared to Deep Flow technique. However, aeration pumps may turn on periodically to reduce the negative impact on AM fungal formation and development [41,53]. A discontinuous aeration approach involving short periods of aeration (2 h periods, four times per day) has been successfully provided adequate aeration with minimal disturbance of the fungus [53].
6.3. Effects of Aeration, Substrate Type, pH, and Inoculation Frequency on Mycorrhizal Colonization
The oxygen concentration (aeration) in hydroponic systems plays a critical role in root and shoot growth as well as nutrient uptake. For instance, shoot and root biomass of AMF-inoculated Sorghum bicolor, and Guizotia abyssinica increased with O2 concentration up to 16% [91]. However, when inoculated plants were exposed to a root oxygen concentration of 21%, as well as the non-aerated conditions, they exhibited lower shoot and root biomass compared to the 16% oxygen concentration [91]. However, that study used a soil-peat-sand mixture, so, this study was not conducted under hydroponic conditions. An assessment of hydroponic culture aiming to develop a tripartite symbiosis involving AMF species (G. intraradices, Gigaspora rosea, and Acaulospora mellea), beans, and rhizobia showed interesting findings [64]. When transplants were transferred from the initial sand culture to nutrient solution, common bean roots were intensely colonized by AMF. Gigaspora colonized well under sand culture conditions, but not in nutrient solution (liquid), while no root-colonization was found with Acaulospora under either culture condition [64].
Monitoring CO2 release in the growing substrate is a useful technique for assessing microorganism activity [134]. An analysis of structure and activity, including CO2 released by substrates, potential enzymatic activity, and lignocellulosic composition, of spontaneous fungal communities in various organic substrates used for soilless culture (wood fibers, coir fibers, and peats) showed that enzymatic activity performance was poorly correlated with microbial respiration and lignocellulosic composition. The structure of fungal communities was unique to each substrate, with the highest specificity for wood fibers and the lowest for peat. Additionally, activity was highly dependent on substrate origin and production process. Overall, the spontaneous development of microorganisms within a soilless growing media could have specific effects on organic matter development and disease reduction [134]. Hydroponic culture pH can significantly influence root colonization of AMF. Sorghum plants (S. bicolor L.) were grown in a soilless system with different pH levels (4.0, 5.0, 6.0, and 7.0) using four VAM isolates. Root colonization varied with pH, increasing as pH levels rose, particularly with certain isolates. Roots colonized by specific isolates showed higher dry matter yield but lower shoot concentrations of certain nutrients. Overall, shoot concentrations of magnesium, calcium, manganese, zinc, and copper remained consistent across VAM isolates [57]. In Thompson’s [135] study, the impact of nutrient concentration and pH on the performance of AMF in wheat and corn was investigated. He found that the presence of the NH4-N form had a negative effect on the colonization process as well as the sporocarp and ectocarpic spores in both crops. For optimal vesicle and arbuscule development by G. mosseae in corn, it was determined that the nutrient solution should have a pH level exceeding 7.4 and a phosphorus content below 0.1% (10 mg L−1). Furthermore, the ideal nutrient solution composition for the interaction between corn and G. fasciculatum was identified as having a pH range between 5.6 and 6.2, with root phosphorus levels below 0.08% (equivalent to 8 mg L−1), and root N concentrations around 1.5% (equivalent to 150 mg L−1). In the case of wheat-G. mosseae interactions in a hydroponic setting, it was found that the most favorable conditions involved a nutrient solution with a pH range of 7.2 to 7.7, root phosphorus levels below 0.055% (equivalent to 5.5 mg L−1), and root N concentrations exceeding 1.07% (equivalent to 170 mg L−1) [135]. To increase the colonization rate in hydroponic systems, one possible approach is to re-inoculate the plants multiple times or add AMF spores to a nutrient solution tank [33]. Double inoculation of AMF and rhizobia favored root AMF colonization and nodules compared to single inoculation [33].
7. Conclusions
AMF have been extensively evaluated for their beneficial effects on crop growth and productivity in conventional (soil-based) agriculture systems. However, their potential in hydroponic systems has received less attention but is still promising. The integration of AMF with various hydroponic techniques offers a range of advantages for crop growth and productivity. Interestingly, the combination of AMF and hydroponics presents a promising approach for modern and eco-friendly crop cultivation. Insights from hydroponic experiments highlight variations in initial colonization rates, with gradual increases observed over time. These differences in colonization patterns between soilless and soil-based cultivation suggest a complex interaction of factors influencing AMF interactions. The level of phosphorus in the nutrient solution in hydroponic systems plays a crucial role in shaping the colonization and sporulation of AMF. Plants exhibit a sophisticated control mechanism over AMF colonization, adjusting it according to their nutritional requirements, particularly phosphate availability.
The complex process of AMF colonization involves a series of finely tuned interactions between the plant and fungus. Plant-secreted flavonoids encourage early fungal growth and communication, while strigolactones mediate hyphal branching and root attachment. Oxygen concentration, pH levels, and nutrient solutions play pivotal roles in shaping the dynamics of AMF colonization in hydroponic systems. The pH levels significantly influence root colonization, and the interaction between pH and specific AMF isolates can impact nutrient uptake. Monitoring microbial activity through CO2 release in substrates reveals substrate-specific effects on organic matter development. Additionally, strategies like double inoculation can enhance root colonization and nodulation in hydroponic systems, offering promising avenues for optimizing AMF interactions in controlled environments.
Overall, although there is evidence supporting the potential advantages of integrating AMF into hydroponic systems, additional research is necessary to optimize their application and evaluate their effectiveness across various scenarios. These scenarios could be when there are nutrient-deficient conditions, drought environments, stressful conditions (salinity, heavy metals), disease suppression, and certain crop requirements. Certainly, exploring future studies and practical applications of combining AMF with hydroponic systems opens up exciting possibilities in agricultural research and practice. Future studies could focus on optimizing the integration of AMF with hydroponic systems to maximize plant growth, nutrient uptake, and overall system efficiency. Further research is also needed to better understand the mechanisms underlying the interactions between AMF and hydroponic systems. It is also essential to assess the economic viability of incorporating AMF into hydroponic production systems. The key consideration lies in determining whether inoculating with AMF enhances the economic yield of a crop compared to solely increasing element concentrations in the nutrient solution.
Author Contributions
Conceptualization, Y.A.O. and M.M.T.; methodology, Y.A.O. and K.M.A.; data curation, Y.A.O.; writing—original draft preparation, Y.A.O., K.M.A. and M.M.T.; writing—review and editing, Y.A.O., K.M.A. and M.M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data available on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Raymond, F.; Pierre, E.; Souleymanou, A.; Therese, N.O.; Fabrice, F.B.; Dieudonne, N. Arbuscular mycorrhizal fungi symbiosis and food security. In Unravelling Plant-Microbe Synergy; Chandra, D., Bhatt, P., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 227–244. [Google Scholar]
- Tzortzakis, N.; Massa, D.; Vandecasteele, B. The tripartite of soilless systems, growing media, and plants through an Intensive Crop Production Scheme. Agronomy 2022, 12, 1896. [Google Scholar] [CrossRef]
- Fussy, A.; Papenbrock, J. An Overview of Soil and Soilless Cultivation Techniques—Chances, Challenges and the Neglected Question of Sustainability. Plants 2022, 1, 1153. [Google Scholar] [CrossRef]
- Shrestha, A.; Dunn, B. Hydroponics. Oklahoma Cooperative Extension Fact Sheets, HLA-6442. 2017. Available online: https://extension.okstate.edu/fact-sheets/print-publications/hla/hydroponics-hla-6442.pdf (accessed on 9 April 2024).
- MMRP. Hydroponics Market by Type (Aggregate Systems, Liquid Systems), Crop Type (Vegetables, Fruits, Flowers), Equipment (HVAC, LED Grow Lights, Irrigation Systems, Material Handling Equipment, Control Systems), Input Type, and Region—Global Forecast to 2026. Available online: https://www.marketsandmarkets.com/Market-Reports/hydroponic-market-94055021.html (accessed on 9 April 2024).
- Tüzel, Y.; Gül, A.; Tüzel, I.; Öztekin, G. Different soilless culture systems and their management. J. Agric. Environ. Sci. 2019, 73, 7–12. [Google Scholar] [CrossRef]
- Azizoglu, U.; Yilmaz, N.; Simsek, O.; Ibal, J.; Tagele, S.; Shin, J. The fate of plant growth-promoting rhizobacteria in soilless agriculture: Future perspectives. 3 Biotech 2021, 11, 382. [Google Scholar] [CrossRef]
- Dasgan, H.Y.; Aldiyab, A.; Elgudayem, F.; Ikiz, B.; Gruda, N.S. Effect of biofertilizers on leaf yield, nitrate amount, mineral content and antioxidants of basil (Ocimum basilicum L.) in a floating culture. Sci. Rep. 2022, 12, 20917. [Google Scholar] [CrossRef]
- Othman, Y.; Tahat, M.; Alananbeh, K.; Al-Ajlouni, M. Arbuscular mycorrhizal fungi inoculation improves flower yield and postharvest quality component of gerbera grown under different salinity levels. Agriculture 2022, 12, 978. [Google Scholar] [CrossRef]
- Utkhede, R. Increased growth and yield of hydroponically grown greenhouse tomato plants inoculated with arbuscular mycorrhizal fungi and Fusarium oxysporum f. sp. radicis-lycopersici. Biocontrol 2006, 51, 393–400. [Google Scholar] [CrossRef]
- Martinez-Garcia, L.; Garcia, K.; Hammer, E.; Vayssières, A. Mycorrhiza for all: An under-earth revolution. New Phytol. 2013, 198, 652–655. [Google Scholar] [CrossRef]
- Sudheer, S.; Johny, L.; Srivastava, S.; Adholeya, A. The trade-in-trade: Multifunctionalities, current market and challenges for arbuscular mycorrhizal fungal inoculants. Symbiosis 2023, 89, 259–272. [Google Scholar] [CrossRef]
- Cano, C.; Bago, A. Aseptic Mycorrhization Inoculant and In Vitro and Ex Vitro Application Methods. WO/2007/014974, 8 February 2007. Available online: https://patentimages.storage.googleapis.com/5e/63/28/a40078860f75e6/WO2007014974A1.pdf (accessed on 9 April 2024).
- Ijdo, M.; Cranenbrouck, S.; Declerck, S. Methods for large-scale production of AM fungi: Past, present, and future. Mycorrhiza 2011, 21, 1–16. [Google Scholar] [CrossRef]
- Sylvia, D.; Jarstfer, A. Sheared Roots as a VA-Mycorrhizal Inoculum and Methods for Enhancing Growth. U.S. Patent No. 5096481, 17 March 1992. Available online: https://patentimages.storage.googleapis.com/e2/f3/94/a39f2da36be624/US5096481.pdf (accessed on 9 April 2024).
- MIO. Mycorrhiza-Based Biofertilizer Market Growth Trends and Forecast (2020–2025). Mordor Intelligence Organization. 2020. Available online: https://www.mordorintelligence.com/industry-reports/mycorrhiza-based-biofertilizer-market (accessed on 9 April 2024).
- Dasgan, H.Y.; Kacmaz, S.; Arpaci, B.B.; Ikiz, B.; Gruda, N.S. Biofertilizers improve the leaf quality of hydroponically grown baby spinach (Spinacia oleracea L.). Agronomy 2023, 13, 575. [Google Scholar] [CrossRef]
- Dasgan, H.Y.; Yilmaz, M.; Dere, S.; Ikiz, B.; Gruda, N.S. Bio-fertilizers reduced the need for mineral fertilizers in soilless-grown Capia pepper. Horticulturae 2023, 9, 188. [Google Scholar] [CrossRef]
- Amaranthus, M.; Eagan, J. Do mycorrhizae have a role in hydroponics? Urban. Gard. Mag. 2009, 8, 72–79. [Google Scholar]
- Cela, F.; Avio, L.; Giordani, T.; Vangelisti, A.; Cavallini, A.; Turrini, A.; Sbrana, C.; Pardossi, A.; Incrocci, L. Arbuscular mycorrhizal fungi increase nutritional quality of soilless grown lettuce while overcoming low phosphorus supply. Foods 2022, 11, 3612. [Google Scholar] [CrossRef]
- Saouy, M.; Pengchai, P.; Choonluchanon, S. Development of arbuscular mycorrhizal spore production in hydroponic culture on leaf lettuce (Lactuca sativar var crispa L.). Chiang Mai Univ. J. Nat. Sci. 2011, 10, 147–158. [Google Scholar]
- Lee, Y.; George, E. Development of a nutrient film technique culture system for arbuscular mycorrhizal plants. HortScience 2005, 40, 378–380. [Google Scholar] [CrossRef]
- Oseni, T.O.; Shongwe, N.; Masarirambi, M. Effect of arbuscular mycorrhiza (AM) inoculation on the performance of tomato nursery seedlings in vermiculite. Int. J. Agric. Biol. 2010, 12, 789–792. [Google Scholar]
- Akiyama, K. Chemical identification and functional analysis of apocarotenoids involved in the development of arbuscular mycorrhizal symbiosis. Biosci. Biotechnol. Biochem. 2007, 71, 1405–1414. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2008; pp. 1–9. [Google Scholar]
- Das, D.; Torabi, S.; Chapman, P.; Gutjahr, C.A. Flexible, low-cost hydroponic co-cultivation system for studying arbuscular mycorrhiza symbiosis. Front. Plant Sci. 2020, 11, 63. [Google Scholar] [CrossRef]
- Smith, S.; Smith, F. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Spanu, P.; Panstruga, R. Editorial, Biotrophic Plant-Microbe Interactions. Front. Plant Sci. 2017, 8, 192. [Google Scholar] [CrossRef]
- Gutjahr, C.; Parniske, M. Cell and developmental biology of arbuscular mycorrhiza symbiosis. Annu. Rev. Cell Dev. Biol. 2013, 29, 593–617. [Google Scholar] [CrossRef]
- Wu, Q.; Srivastava, A.; Zou, Y.; Malhotra, S. Mycorrhizas in citrus: Beyond soil fertility and plant nutrition. Ind. J. Agric. Sci. 2017, 87, 427–443. [Google Scholar] [CrossRef]
- Rask, K.; Johansen, J.; Kjøller, R.; Ekelund, F. Differences in arbuscular mycorrhizal colonization influence cadmium uptake in plants. Environ. Exp. Bot. 2019, 162, 223–229. [Google Scholar] [CrossRef]
- Al-Karaki, G.; Othman, Y.; Al-Ajmi, A. Effects of mycorrhizal fungi inoculation on landscape turf establishment under Arabian Gulf region conditions. Arab. Gulf J. Sci. Res. 2007, 25, 147–152. [Google Scholar]
- Liu, X.; Xie, M.; Hashem, A.; Abd-Allah, E.; Wu, Q. Arbuscular mycorrhizal fungi and rhizobia synergistically promote root colonization, plant growth, and nitrogen acquisition. Plant Growth Regul. 2023, 100, 691–701. [Google Scholar] [CrossRef]
- Tahat, M.; Alananbeh, K.; Othman, Y.; Leskovar, D. Soil health and sustainable agriculture. Sustainability 2020, 12, 4859. [Google Scholar] [CrossRef]
- Robinson, B.L.; Feng, W.; Gulbis, N.; Hajdu, K.; Harrison, R.; Jeffries, P.; Xu, X. The use of arbuscular mycorrhizal fungi to improve strawberry production in coir substrate. Front. Plant Sci. 2016, 7, 1237. [Google Scholar] [CrossRef]
- Hung, L.; Sylvia, D. Production of vesicular–arbuscular mycorrhizal fungus inoculum in aeroponic culture. Appl. Environ. Microbiol. 1988, 54, 353–357. [Google Scholar] [CrossRef]
- Miller, S.; Sharitz, R. Manipulation of flooding and arbuscular mycorrhiza formation influences growth and nutrition of two semiaquatic grass species. Funct. Ecol. 2000, 14, 738–748. [Google Scholar] [CrossRef]
- Sembok, W.; Kassim, N.; Hamzah, Y.; Rahman, Z. Effects of mycorrhizal inoculation on growth and quality f Roselle (Hibiscus sabdariffa L. ) grown in soilless culture system. Malays. Appl. Bio. 2015, 44, 57–62. [Google Scholar]
- Íkiz, Ö.; Abak, K.; Daşgan, H.Y.; Ortaş, I. Effects of mycorrhizal inoculation in soilless culture on pepper plant growth. Acta Hortic. 2009, 807, 533–540. [Google Scholar] [CrossRef]
- Besmer, Y.; Koide, R. Effect of mycorrhizal colonization and phosphorus on ethylene production by snapdragon (Antirrhinum majus L.) flowers. Mycorrhiza 1999, 9, 161–166. [Google Scholar] [CrossRef]
- Dugassa, D.; Grunewaldt-Stöcker, G.; Schönbeck, F. Growth of Glomus intraradices and its effect on linseed (Linum usitatissimum L.) in hydroponic culture. Mycorrhiza 1995, 5, 279–282. [Google Scholar]
- Jarstfer, A.; Farmer-Koppenol, S.; Sylvia, D. Tissue magnesium and calcium affect arbuscular mycorrhiza development and fungal reproduction. Mycorrhiza 1988, 7, 237–342. [Google Scholar] [CrossRef]
- Bitterlich, M.; Franken, P.; Graefe, J. Atmospheric drought and low light impede mycorrhizal effects on leaf photosynthesis-a glasshouse study on tomato under naturally fluctuating environmental conditions. Mycorrhiza 2019, 29, 13–28. [Google Scholar] [CrossRef]
- Hayek, S.; Grosch, R.; Gianinazzi-Pearson, V.; Franken, P. Bio-protection and alternative fertilization of petunia using mycorrhiza in a soilless production system. Agron. Sustain. Dev. 2012, 32, 765–771. [Google Scholar] [CrossRef]
- Cekic, C.; Yilmaz, E. Effect of arbuscular mycorrhiza and different doses of phosphor on vegetative and generative components of strawberries applied with different phosphor doses in soilless culture. Afr. J. Agric. Res. 2011, 6, 4736–4739. [Google Scholar]
- Palencia, P.; Martínez, F.; Pestana, M.; Oliveira, J.; Correia, P. Effect of Bacillus velezensis and Glomus intraradices on fruit quality and growth parameters in strawberry soilless growing system. Hortic. J. 2015, 84, 122–130. [Google Scholar] [CrossRef]
- Dasgan, H.; Kusvuran, S.; Ortas, I. Responses of soilless grown tomato plants to arbuscular mycorrhizal fungal (Glomus fasciculatum) colonization in re-cycling and open systems. Afr. J. Biotechnol. 2008, 7, 3606–3613. [Google Scholar]
- Bhowmik, S.; Yadav, G.; Datta, M. Rapid mass multiplication of Glomus mosseae inoculum as influenced by some biotic and abiotic factors. Bangladesh, J. Bot. 2015, 44, 209–214. [Google Scholar] [CrossRef]
- Cecatto, A.; Ruiz, F.; Calvete, E.; Martínez, J.; Palencia, P. Mycorrhizal inoculation affects the phytochemical content in strawberry fruits. Acta Sci. 2016, 38, 227–237. [Google Scholar] [CrossRef]
- Elmes, R.P.; Mosse, B. Vesicular-arbuscular endomycorrhizal inoculum production. II. Experiments with maize (Zea mays) and other hosts in nutrient flow culture. Can. J. Bot. 1984, 62, 1531–1536. [Google Scholar] [CrossRef]
- Eltrop, L.; Marschner, H. Growth and mineral nutrition of nonmycorrhizal and mycorrhizal Norway spruce (Picea abies) seedlings grown in semi-hydroponic sand culture, II. Carbon partitioning in plants supplied with ammonium or nitrate. New Phytol. 1996, 133, 479–486. [Google Scholar] [CrossRef]
- Gryndler, M.; Hršelová, H.; Sudová, R.; Gryndlerová, H.; Řezáčová, V.; Merhautová, V. Hyphal growth and mycorrhiza formation by the arbuscular mycorrhizal fungus Glomus claroideum BEG 23 is stimulated by humic substances. Mycorrhiza 2005, 15, 483–488. [Google Scholar] [CrossRef]
- Hawkins, H.; George, E. Hydroponic culture of the mycorrhizal fungus Glomus mosseae with Linum usitatissimum L., Sorghum bicolor L. and Triticum aestivum L. Plant Soil 1997, 196, 143–149. [Google Scholar] [CrossRef]
- Hawkins, H.; Cramer, M.; George, E. Root respiratory quotient and nitrate uptake in hydroponically grown non-mycorrhizal and mycorrhizal wheat. Mycorrhiza 1999, 9, 57–60. [Google Scholar] [CrossRef]
- Kowalska, I.; Konieczny, A.; Gąstoł, M. Effect of mycorrhiza and the phosphorus content in a nutrient solution on the yield and nutritional status of lettuce grown on various substrates. J. Elem. 2015, 20, 631–642. [Google Scholar] [CrossRef]
- Maboko, M.; Bertling, I.; Plooy, C. Arbuscular mycorrhiza has limited effects on yield and quality of tomatoes grown under soilless cultivation. Acta Agric. Scand. B Soil Plant Sci. 2013, 63, 261–270. [Google Scholar] [CrossRef]
- Medeiros, C.; Clark, R.B.; Ellis, J.R. Growth and nutrient uptake of sorghum cultivated with vesicular-arbuscular mycorrhiza isolates at varying pH. Mycorrhiza 1994, 4, 185–191. [Google Scholar] [CrossRef]
- Mosse, B.; Thompson, J. Vesicular-arbuscular endomycorrhizal inoculum production. I. Exploratory experiments with beans (Phaseolus vu1gnris) in nutrient flow culture. Can. J. Bot. 1983, 62, 1523–1530. [Google Scholar] [CrossRef]
- Nurbaity, A.; Istifadah, N.; Haryantini, B.; Ilhami, M.; Habibullah, M.; Arifin, M. Optimization of hydroponic technology for production of mycorrhiza biofertilizer. IOP Conf. Ser. Environ. Earth Sci. 2019, 347, 012017. [Google Scholar] [CrossRef]
- Ojala, J.; Jarrell, M. Hydroponic sand culture systems for mycorrhizal research. Plant Soil 1980, 57, 297–303. [Google Scholar] [CrossRef]
- Oztekin, G.; Tuzel, Y.; Tuzel, H. Does mycorrhiza improve salinity tolerance in grafted plants? Sci. Hortic. 2013, 149, 55–60. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kim, K.; Walitang, D.; Chanratana, M.; Kang, Y.; Chung, B.; Sa, T. Trap culture technique for propagation of arbuscular mycorrhizal fungi using different host plants. Korean J. Soil Sci. Fert. 2016, 49, 608–613. [Google Scholar] [CrossRef]
- Selvakumar, G.; Shagol, C.; Kang, Y.; Chung, B.N.; Han, S.G.; Sa, T. Arbuscular mycorrhizal fungi spore propagation using single spore as starter inoculum and a plant host. J. Appl. Microbiol. 2018, 124, 1556–1565. [Google Scholar] [CrossRef]
- Tajini, F.; Suriyakup, P.; Vailhe, H.; Jansa, J.; Drevon, J. Assess suitability of hydroaeroponic culture to establish tripartite symbiosis between different AMF species, beans, and rhizobia. BMC Plant Biol. 2009, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- White, J.; Charvat, I. The mycorrhizal status of an emergent aquatic, Lythrum salicaria L., at different levels of phosphorus availability. Mycorrhiza 1999, 9, 191–197. [Google Scholar] [CrossRef]
- Zou, Y.; Wu, Q. Sodium chloride stress induced changes in leaf osmotic adjustment of trifoliate orange (Poncirus trifoliata) seedlings inoculated with mycorrhizal fungi. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 64–69. [Google Scholar] [CrossRef]
- Al-Karaki, G. Benefit-cost and water-use efficiency of arbuscular mycorrhizal durum wheat grown under drought stress. Mycorrhiza 1998, 8, 41–45. [Google Scholar] [CrossRef]
- Al-Karaki, G. Growth of mycorrhizal tomato and mineral acquisition under salt stress. Mycorrhiza 2000, 10, 51–54. [Google Scholar] [CrossRef]
- Al-Karaki, G. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar] [CrossRef]
- Arruda, B.; Herrera, W.; Rojas-García, J.; Turner, C.; Pavinato, P. Cover crop species and mycorrhizal colonization on soil phosphorus dynamics. Rhizosphere 2021, 19, 100396. [Google Scholar] [CrossRef]
- Beltrano, J.; Ronco, M. Improved tolerance of wheat plants (Triticum aestivum L.) to drought stress and rewatering by the arbuscular mycorrhizal fungus Glomus claroideum: Effect on growth and cell membrane stability. Braz. J. Plant Physiol. 2008, 20, 29–37. [Google Scholar] [CrossRef]
- Betancur-Agudelo, M.; Meyer, E.; Lovato, P. Arbuscular mycorrhizal fungus richness in the soil and root colonization in vineyards of different ages. Rhizosphere 2021, 17, 100307. [Google Scholar] [CrossRef]
- Chandrasekeran, A.; Mahalingam, P. Diversity of arbuscular mycorrhizae fungi from orchard ecosystem. J. Plant Pathol. Microbiol. 2014, 5, 2. [Google Scholar] [CrossRef]
- Chenchouni, H.; Mekahlia, M.; Beddiar, A. Effect of inoculation with native and commercial arbuscular mycorrhizal fungi on growth and mycorrhizal colonization of olive (Olea europaea L.). Sci. Hortic. 2020, 261, 108969. [Google Scholar] [CrossRef]
- Chiomento, J.; De Nardi, F.; Filippi, D.; Trentin, T.; Dornelles, A.; Fornari, M.; Nienow, A.; Calvete, E. Morpho-horticultural performance of strawberry cultivated on substrate with arbuscular mycorrhizal fungi and biochar. , Sci. Hortic. 2021, 282, 110053. [Google Scholar] [CrossRef]
- Ding, Y.; Fan, Q.; He, J.; Wu, H.; Zou, Y.; Wu, Q.; Kuča, K. Effects of mycorrhizas on physiological performance and root TIPs expression in trifoliate orange under salt stress. Arch. Agron. Soil Sci. 2020, 66, 182–192. [Google Scholar] [CrossRef]
- Hart, M.; Ehret, D.L.; Krumbein, A.; Leung, C.; Murch, S.; Turi, C.; Franken, P. Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 2015, 25, 359–376. [Google Scholar] [CrossRef]
- Hashem, A.; Akhter, A.; Alqarawi, A.; Singh, G.; Almutairi, K.; Abd_Allah, E. Mycorrhizal fungi induced activation of tomato defense system mitigates Fusarium wilt stress. Saudi J. Biol. Sci. 2021, 28, 5442–5450. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, M.; Liu, H.; Zhao, Q.; Lin, X. Intercropping with sweet corn (Zea mays L. var. Rugosa Bonaf.) expands P acquisition channels of chili pepper (Capsicum annuum L.) via arbuscular mycorrhizal hyphal networks. J. Soils Sediments 2019, 19, 1632–1639. [Google Scholar] [CrossRef]
- Huang, Y.; Srivastava, A.; Zou, Y.; Ni, Q.; Han, Y.; Wu, Q. Mycorrhizal-induced calmodulin mediated changes in antioxidant enzymes and growth response of drought-stressed trifoliate orange. Front. Microbiol. 2014, 5, 682. [Google Scholar] [CrossRef] [PubMed]
- Keller-Pearson, M.; Liu, Y.; Peterson, A.; Pederson, K.; Willems, L.; Ané, J.; Silva, E. Inoculation with arbuscular mycorrhizal fungi has a more significant positive impact on the growth of open-pollinated heirloom varieties of carrots than on hybrid cultivars under organic management conditions. Agric. Ecosyst. Environ. 2020, 289, 106712. [Google Scholar] [CrossRef]
- Khalil, H.; Eissa, A.; El-Shazly, S.; Aboul-Nasr, A. Improved growth of salinity-stressed citrus after inoculation with mycorrhizal fungi. Sci. Hortic. 2011, 130, 624–632. [Google Scholar] [CrossRef]
- Leventis, G.; Tsiknia, M.; Feka, M.; Ladikou, E.V.; Papadakis, I.E.; Chatzipavlidis, I.; Papadopoulou, K.; Ehaliotis, C. Arbuscular mycorrhizal fungi enhance growth of tomato under normal and drought conditions, via different water regulation mechanisms. Rhizosphere 2021, 19, 100394. [Google Scholar] [CrossRef]
- Mason, P.; Ibrahim, K.; Ingleby, K.; Munro, R.; Wilson, J. Mycorrhizal development and growth of inoculated Eucalyptus globulus (Labill.) seedlings in wet and dry conditions in the glasshouse. For. Ecol. Manag. 2000, 128, 269–277. [Google Scholar] [CrossRef]
- Mathur, N.; Singh, J.; Bohra, S.; Bohra, A.; Vyas, A. Arbuscular mycorrhizal fungi alleviate salt stress of Trichosanthes dioica Roxb. Acta Agric. Scand. B Soil Plant Sci. 2010, 60, 510–516. [Google Scholar]
- Naseer, M.; Zhu, Y.; Li, F.; Yang, Y.; Wang, S.; Xiong, Y. Nano-enabled improvements of growth and colonization rate in wheat inoculated with arbuscular mycorrhizal fungi. Environ. Pollut. 2022, 295, 118724. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Pérez-Tornero, O.; Morte, A. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014, 171, 76–85. [Google Scholar] [CrossRef]
- Ngo, H.; Watts-Williams, S.; Cavagnaro, T. Mycorrhizal growth and phosphorus responses of tomato differ with source but not application rate of phosphorus fertilizers. Appl. Soil Ecol. 2021, 166, 104089. [Google Scholar] [CrossRef]
- Ngwene, B.; Gabriel, E.; George, E. Influence of different mineral nitrogen sources (NO3−-N vs. NH4+-N) on arbuscular mycorrhiza development and N transfer in a Glomus intraradices–cowpea symbiosis. Mycorrhiza 2013, 23, 107–117. [Google Scholar] [CrossRef]
- Ruscitti, M.; Arango, M.; Beltrano, J. Improvement of copper stress tolerance in pepper plants (Capsicum annuum L.) by inoculation with arbuscular mycorrhizal fungi. Theor. Exp. Plant Physiol. 2017, 29, 37–49. [Google Scholar] [CrossRef]
- Saif, S. The influence of soil aeration on the efficiency of vesicular-arbuscular mycorrhizae II. Effect of soil oxygen on growth and mineral uptake in Eupatorium odoratum L., Sorghum bicolor (L.) Moench and Guizotia abyssinica (L.f.) Cass. inoculated with vesicular-arbuscular mycorrhizal fungi. New Phytol. 1983, 95, 405–417. [Google Scholar]
- Schubert, R.; Werner, S.; Cirka, H.; Rödel, P.; Tandron, M.Y.; Mock, H.-P.; Hutter, I.; Kunze, G.; Hause, B. Effects of arbuscular mycorrhization on fruit quality in industrialized tomato production. Int. J. Mol. Sci. 2020, 21, 7029. [Google Scholar] [CrossRef]
- Sensoy, S.; Demir, S.; Turkmen, O.; Erdinc, C.; Savur, O. Responses of some different pepper (Capsicum annuum L.) genotypes to inoculation with two different arbuscular mycorrhizal fungi. Sci. Hortic. 2007, 113, 92–95. [Google Scholar] [CrossRef]
- Singh, A.; Thakur, A.; Sharma, S.; Gill, P.; Kalia, A. Bio-inoculants enhance growth, nutrient uptake, and buddability of citrus plants under protected nursery condition. Commun. Soil Sci. Plant Anal. 2018, 49, 2571–2586. [Google Scholar] [CrossRef]
- Vani, M.S.; Hindumathi, A.; Reddy, B.N. Beneficial effect of arbuscular mycorrhizal fungus, Glomus fasciculatum on plant growth and nutrient uptake in tomato. Indian. Phytopathol. 2018, 71, 115–122. [Google Scholar] [CrossRef]
- Vázquez, E.; Benito, M.; Espejo, R.; Teutscherova, N. No-tillage and liming increase the root mycorrhizal colonization, plant biomass and N content of a mixed oat and vetch crop. Soil Tillage Res. 2020, 200, 104623. [Google Scholar] [CrossRef]
- Wang, H.; An, T.; Huang, D.; Liu, R.; Xu, B.; Zhang, S.; Deng, X.; Siddique, K.; Chen, Y. Arbuscular mycorrhizal symbioses alleviating salt stress in maize is associated with a decline in root-to-leaf gradient of Na+/K+ ratio. BMC Plant Biol. 2021, 21, 457. [Google Scholar] [CrossRef]
- Wilson, G.; Hartnett, D. Interspecific variation in plant responses to mycorrhizal colonization in tallgrass prairie. Am. J. Bot. 1998, 85, 1732–1738. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; He, X.H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant. 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Liu, W.; Ye, X.F.; Zai, H.F.; Zhao, L.J. Alleviation of salt stress in citrus seedlings inoculated with mycorrhiza: Changes in leaf antioxidant defense systems. Plant Soil Environ. 2010, 56, 470–475. [Google Scholar] [CrossRef]
- Wu, Q.; Zou, Y.; He, X. Mycorrhizal symbiosis enhances tolerance to NaCl stress through selective absorption but not selective transport of K+ over Na+ in trifoliate orange. Scient. Horticul. 2013, 160, 366–374. [Google Scholar] [CrossRef]
- Ziane, H.; Hamza, N.; Meddad-Hamza, A. Arbuscular mycorrhizal fungi and fertilization rates optimize tomato (Solanum lycopersicum L.) growth and yield in a Mediterranean agroecosystem. J. Saudi Soc. Agricul. Sci. 2021, 20, 454–458. [Google Scholar] [CrossRef]
- Shaik, A.; Singh, S. Influence of arbuscular mycorrhizal fungi on physiology and yield of eggplant in organic soilless production system. HortScience 2022, 57, 759–768. [Google Scholar] [CrossRef]
- Haghighi, M.; Mohmmadnia, S.; Pessarakli, M. Effects of mycorrhiza colonization on growth, root exudates, antioxidant activity and photosynthesis trait of cucumber grown in Johnson modified nutrient solution. J. Plant Nutr. 2016, 39, 2079–2091. [Google Scholar] [CrossRef]
- Caser, M.; Demasi, S.; Victorino, Í.M.M.; Donno, D.; Faccio, A.; Lumini, E.; Bianciotto, V.; Scariot, V. Arbuscular mycorrhizal fungi modulate the crop performance and metabolic profile of saffron in soilless cultivation. Agronomy 2019, 9, 232. [Google Scholar] [CrossRef]
- Dere, S.; Coban, A.; Akhoundnejad, Y.; Ozsoy, S.; Dasgan, H. Use of mycorrhiza to reduce mineral fertilizers in soilless melon (Cucumis melo L.) cultivation. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 1331–1336. [Google Scholar] [CrossRef]
- Roussis, I.; Beslemes, D.; Kosma, C.; Triantafyllidis, V.; Zotos, A.; Tigka, E.; Mavroeidis, A.; Karydogianni, S.; Kouneli, V.; Travlos, I.; et al. The Influence of arbuscular mycorrhizal fungus Rhizophagus irregularis on the growth and quality of processing tomato (Lycopersicon esculentum Mill.) seedlings. Sustainability 2022, 14, 9001. [Google Scholar] [CrossRef]
- Cardoso, D.S.C.P.; Martinez, H.E.P.; de Abreu, J.A.A.; Kasuya, M.C.M.; Sediyama, M.A.N. Growth-promoting fungi and potassium doses affects productivity and nutrition of cherry-type tomatoes. J. Plant Nutr. 2023, 46, 835–851. [Google Scholar] [CrossRef]
- Michałojć, Z.; Jarosz, Z.; Pitura, K.; Dzida, K. Effect of mycorrhizal colonization and nutrient solutions concentration on the yielding and chemical composition of tomato grown in rockwool and straw medium. Acta Sci. Pol. Hortorum Cultus 2015, 14, 15–27. [Google Scholar]
- Haghighi, M.; Mozafariyan, M.; Abdolahipour, B. Effect of cucumber mycorrhiza inoculation under low and high root temperature grown on hydroponic conditions. J. Crop Sci. Biotech. 2015, 18, 89–96. [Google Scholar] [CrossRef]
- Fakhro, A.; Andrade-Linares, D.; Bargen, S.; Bandte, M.; Büttner, C.; Grosch, R.; Schwarz, D.; Franken, P. Impact of Piriformospora indica on tomato growth and on interaction with fungal and viral pathogens. Mycorrhiza 2010, 20, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, K.; Nazarideljou, M. Morpho-physiological and quality attributes of gerbera (Gerbera jamesonii) cut flower under inoculated substrate with arbuscular mycorrhizal fungi in soilless system. J. Sci. Technol. 2017, 8, 27–39. [Google Scholar] [CrossRef]
- Dasgan, H.Y.; Cetinturk, T.; Altuntas, O. The effects of biofertilizers on soilless organically grown greenhouse tomato. Acta Hortic. 2017, 1164, 555–561. [Google Scholar] [CrossRef]
- Ullah, F.; Ullah, H.; Ishfaq, M.; Khan, R.; Gul, S.L.; Gulfraz, A.; Wang, C.; Zhifang, L. Genotypic variation of tomato to AMF inoculation in improving growth, nutrient uptake, yield, and photosynthetic activity. Symbiosis 2024, 92, 111–124. [Google Scholar] [CrossRef]
- Noor, H.; Ahmad, H.; Sayuti, Z. Effect of mycorrhiza, fertilizers and planting media on rock melon (Cucumis Melo Linn Cv. Glamour) growth using the canopytechture structure. Int. J. Appl. Agric. Sci. 2019, 5, 14–19. [Google Scholar] [CrossRef]
- Mishra, V.; Ellouze, W.; Howard, R. Utility of arbuscular mycorrhizal fungi for improved production and disease mitigation in organic and hydroponic greenhouse crops. J. Hortic. 2018, 5, 237. [Google Scholar] [CrossRef]
- Garcés-Ruiz, M.; Calonne-Salmon, M.; Plouznikoff, K.; Misson, C.; Navarrete-Mier, M.; Cranenbrouck, S.; Declerck, S. Dynamics of short-term phosphorus uptake by intact mycorrhizal and non-mycorrhizal maize plants grown in a circulatory semi-hydroponic cultivation system. Front. Plant Sci. 2017, 8, 285559. [Google Scholar] [CrossRef]
- Estrada-Luna, A.; Davies, F., Jr. Arbuscular mycorrhizal fungi influence water relations, gas exchange, abscisic acid and growth of micropropagated Chile Ancho pepper (Capsicum annuum) plantlets during acclimatization and post-acclimatization. J. Plant Physiol. 2003, 1, 1073–1083. [Google Scholar] [CrossRef]
- Wang, F.; Adams, C.; Yang, W.; Sun, Y.; Shi, Z. Benefits of arbuscular mycorrhizal fungi in reducing organic contaminant residues in crops: Implications for cleaner agricultural production. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1580–1612. [Google Scholar] [CrossRef]
- Feng, J.; Lv, W.; Xu, J.; Huang, Z.; Rui, W.; Lei, X.; Ju, X.; Li, Z. Overlapping Root Architecture and Gene Expression of Nitrogen Transporters for Nitrogen Acquisition of Tomato Plants Colonized with Isolates of Funneliformis mosseae in Hydroponic Production. Plants 2022, 11, 1176. [Google Scholar] [CrossRef]
- Othman, Y.; A’saf, T.; Al-Ajlouni, M.; Bany-Hani, M.; St-Hilaire, R. Holding solution pH and composition consistently improve vase life of rose, lily and gerbera. J. Phytol. 2023, 15, 57–62. [Google Scholar] [CrossRef]
- Ratte, H. Bioaccumulation and toxicity of silver compounds: A review. Environ. Toxicol. Chem. 1999, 18, 89–108. [Google Scholar] [CrossRef]
- Mohammad, A.; Khan, A.; Kuek, C. Improved aeroponic culture of inocula of arbuscular mycorrhizal fungi. Mycorrhiza 2000, 9, 337–339. [Google Scholar] [CrossRef]
- Qian, Y.; Zhao, G.; Zhou, J.; Zhao, H.; Mutter, T.; Huang, X. Combined bioremediation of bensulfuron-methyl contaminated soils with arbuscular mycorrhizal fungus and Hansschlegelia zhihuaiae S113. Front. Microbiol. 2022, 13, 843525. [Google Scholar] [CrossRef]
- Ballhorn, D.J.; Younginger, B.S.; Kautz, S. An aboveground pathogen inhibits belowground rhizobia and arbuscular mycorrhizal fungi in Phaseolus vulgaris. BMC Plant Biol. 2014, 14, 321. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Wu, Q.; Chang, W.; Fan, X.; Song, F. Proteomic analysis of the response of Funnelifor mismosseae/Medicago sativa to atrazine stress. BMC Plant Biol. 2018, 18, 289. [Google Scholar] [CrossRef]
- Douds, D.; Schenck, N. Increased sporulation of vesicular–arbuscular mycorrhizal fungi by manipulation of nutrient regimens. Appl. Environ. Microbiol. 1990, 56, 413–418. [Google Scholar] [CrossRef]
- Kobae, Y.; Ohmori, Y.; Saito, C.; Yano, K.; Ohtomo, R.; Fujiwara, T. Phosphate treatment strongly inhibits new arbuscule development but not the maintenance of arbuscule in mycorrhizal rice roots. Plant Physiol. 2016, 171, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Valentine, A.; Kleinert, A. Respiratory responses of arbuscular mycorrhizal roots to short-term alleviation of P deficiency. Mycorrhiza 2007, 17, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P.; Herrmann, L.; Lesueur, D.; Robin, A.; Trap, J.; Waithaisong, K.; Plassard, C. Impact of roots, microorganisms and microfauna on the fate of soil phosphorus in the rhizosphere. Annu. Plant Rev. 2015, 48, 377–408. [Google Scholar]
- Vejsadová, H.; Čšatská, V.; Hršelová, H.; Gryndler, M. Influence of bacteria on growth and phosphorus nutrition of mycorrhizal corn. J. Plant Nutr. 1993, 16, 1857–1866. [Google Scholar] [CrossRef]
- Choi, J.; Summers, W.; Paszkowski, U. Mechanisms underlying establishment of arbuscular mycorrhizal symbioses. Annu. Rev. Phytopathol. 2018, 56, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Nadal, M.; Paszkowski, U. Polyphony in the rhizosphere: Presymbiotic communication in arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2013, 16, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Montagne, V.; Charpentier, S.; Cannavo, P.; Capiaux, H.; Grosbellet, C.; Lebeau, T. Structure and activity of spontaneous fungal communities in organic substrates used for soilless crops. Sci. Hortic. 2015, 192, 148–157. [Google Scholar] [CrossRef]
- Thompson, J. Soilless culture of vesicular-arbuscular mycorrhizae of cereals: Effects of nutrient concentration and nitrogen source. Can. J. Bot. 1986, 64, 2282–2294. [Google Scholar] [CrossRef]
Figure 1.
Cumulative publications on mycorrhiza and hydroponics-related keywords in prominent scientific databases, 2002–2023. The search for each keyword in the databases was limited to include the keyword exclusively in the title.
Figure 1.
Cumulative publications on mycorrhiza and hydroponics-related keywords in prominent scientific databases, 2002–2023. The search for each keyword in the databases was limited to include the keyword exclusively in the title.

Figure 2.
Comparative root colonization (%) between hydroponic (soilless) and soil-based systems. Sources for AMF under hydroponic cultivation: [10,21,22,23,26,36,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65]. Sources for AMF under soil-based system: [9,31,32,36,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102]. Bars followed by different letters are significantly different at p < 0.05.
Figure 2.
Comparative root colonization (%) between hydroponic (soilless) and soil-based systems. Sources for AMF under hydroponic cultivation: [10,21,22,23,26,36,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65]. Sources for AMF under soil-based system: [9,31,32,36,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102]. Bars followed by different letters are significantly different at p < 0.05.

Figure 3.
Influence of AMF on crop yield and quality in hydroponic environments. The study was counted positive when the measured variable from AMF × hydroponic was higher than un-inoculated while the negative was when the response variable results from inoculated plants was less than control. Sources: [8,10,20,21,22,23,26,38,40,41,43,44,45,46,47,48,49,50,51,52,53,55,56,57,59,60,64,65,103,105,106,107,108,109,110,111,112,113,114,115].
Figure 3.
Influence of AMF on crop yield and quality in hydroponic environments. The study was counted positive when the measured variable from AMF × hydroponic was higher than un-inoculated while the negative was when the response variable results from inoculated plants was less than control. Sources: [8,10,20,21,22,23,26,38,40,41,43,44,45,46,47,48,49,50,51,52,53,55,56,57,59,60,64,65,103,105,106,107,108,109,110,111,112,113,114,115].

Figure 5.
Effect of different phosphorus concentration on AMF root colonization. Sources: [22,25,40,44,47,50,53,55,56,65,106,113,123,128,129]. Bars followed by different letters are significantly different at p < 0.05.


{kind=link}
{kind=link}
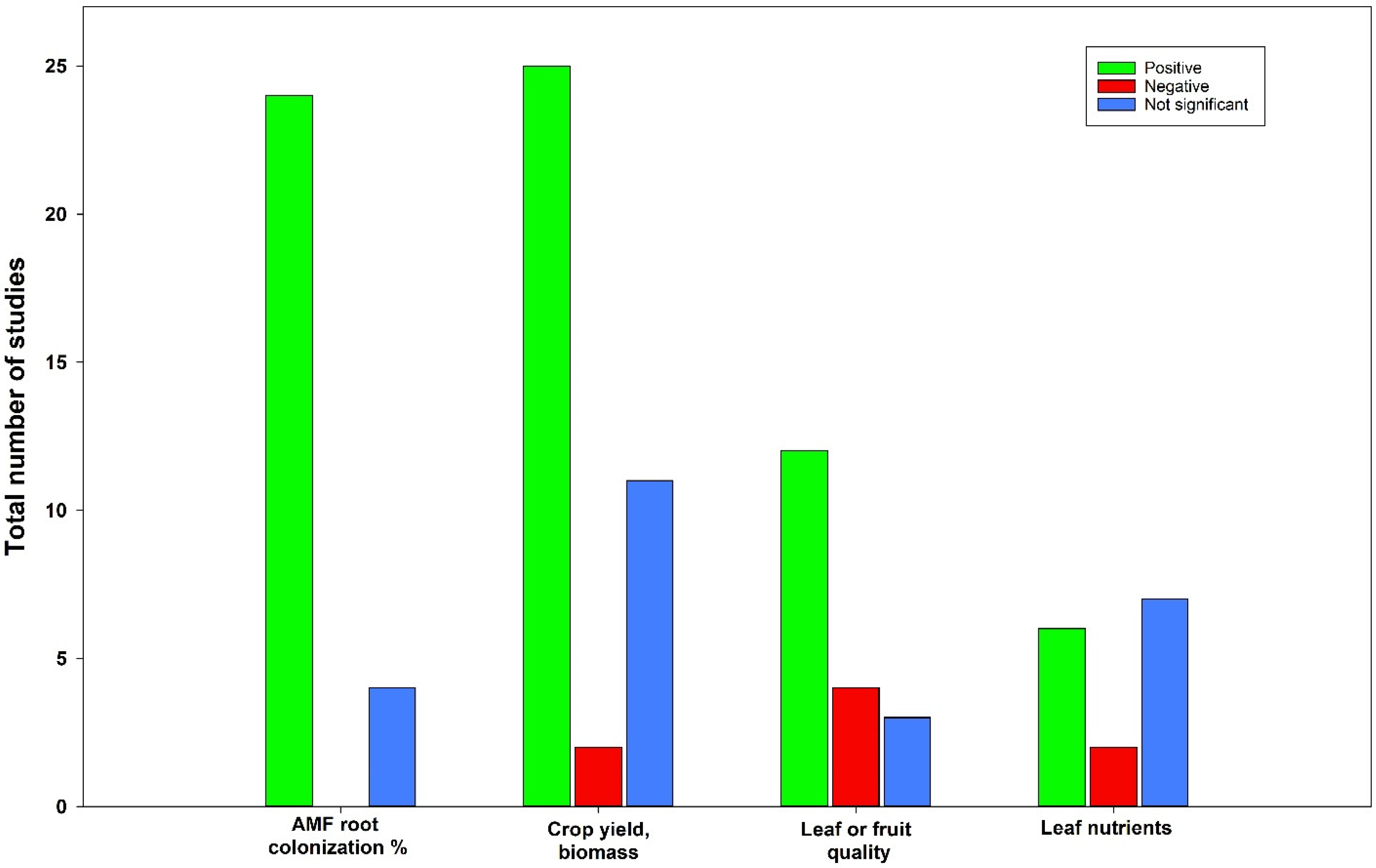{kind=link}
{kind=link}
{kind=link}
Table 1.
Crops reactions to AMF within hydroponic systems.
| Plant Host | AMF | Substrate/Hydroponic Type | Response | References |
|---|---|---|---|---|
| Lettuce (Lactuca sativa L.) | F. mosseae | Calcined clay/Ebb and flow. | AMF improved leaf gas exchanges and the nutritional quality (leaf content of chlorophylls, carotenoids, and phenols) of lettuce even at sub-optimal P. | [20] |
| Basil (O. basilicum L.) | Mixture of G. intraradices, G. aggregatum, G. mosseae, G. clarum, G. monosporus, G. deserticola, G. brasilianum, G. etunicatum, and Gigaspora margarita. | No substrate/Floating culture. | Compared to the control (none treated) bacteria, mycorrhiza, and micro-algae treatments increased basil yield by about 18.94%, 13.94%, and 5.72%, respectively. In addition, bio-fertilizers enhanced the intake of nutrients N, P, K, Ca, Mg, Fe, Mn, Zn and Cu. | [8] |
| Tomato (L. esculentum Mill.) | Rhizophagus irregularis | Mixture of organic peat and vermiculite (1:1 v/v)/Ebb and flow. | AMF transplants had higher shoot total dry weight, survival rate, leaf N and P content. | [107] |
| Tomato | Piriformospora indica fungi + mixture of mixture of AMF (Rhizophagus clarus, C. etunicatum and Gigaspora albida) | Coconut fiber/Ebb and flow. | Growth-promoting fungi and K doses improved cherry tomato yield and fruit quality. | [108] |
| Baby Spinach (S. oleracea L.) | G. intraradices, G. aggregatum, G. mosseae, G. clarum, G. monosporus, G. deserticola, G. brasilianum, G. etunicatum, G. margarita | No substrate/Float culture. | Biofertilizers improved the internal leaves’ quality (total phenolic, vitamin C, total soluble solids). | [17] |
| Onion (Allium cepa) | Glomus sp. (INVAM-FL329) | Sand/Ebb and flow. | Higher levels of magnesium sulfate (2.6 vs. 11.7 mM MgSO4) in nutrient solution significantly reduced tissue-Ca levels, root colonization and sporulation. | [42] |
| Tomato | G. monosporum and G. mosseae | Sawdust/Ebb and flow. | AMF significantly increased plant height, fruit yield and fruit number. | [10] |
| Saffron (C. sativus) | R. intraradices | Sand/Ebb and flow. | Corms had similar spice yield compared to soil, but these corms had higher polyphenols, anthocyanins, vitamin C, and elevated antioxidant activity. | [105] |
| Tomato | G. mosseae, G. intraradices, G. etunicatum and G. clarum | Rockwool/Ebb and flow. | Although AMF had no effect on the total and marketable yield, tomato fruit inoculated with AMF contained higher sugars compared to the plants growing without mycorrhiza. | [109] |
| Tomato | F. mosseae | Sand: vermiculite mixture (1:1 v/v)/Ebb and flow | Leaf photosynthetic capacities were higher in mycorrhizal plants when leaves contained more proteins and/or the plant-internal moisture stress was lower than in non-mycorrhizal plants. | [43] |
| Petunia (Petunia × hybrid) | G. mosseae, G. intraradices and Gigaspora rosea | Vermiculite/Ebb and flow. | Disease symptoms caused by the root pathogen Thielaviopsis basicola were significantly reduced in G. mosseae colonized plants. A negative effect of the pathogen on root colonization by G. intraradices was observed. | [44] |
| Lettuce | G. mosseae | No substrate/NFT, Perlite/Ebb and flow. | In both systems, AMF-inoculated lettuce had higher shoot dry weight than the non-inoculated. | [22] |
| Lettuce | G. verruculosum | No substrate/Float culture. | AMF-inoculated lettuce plants had higher mycorrhizal colonization (86%), spore population (no/g sand, 303) and shoot and root dry weight compared to the control | [21] |
| Tomato | G. etunicatum and G. intraradices | Vermiculite/Ebb and flow. | AMF-inoculated seedling had higher biomass and relative growth rate compared to the control. | [23] |
| Strawberry (Fragaria × Ananassa) | G. clarum and G. caledonium | Sterilized potting mix soil: sand: perlite (1:1:1 v/v/v)/Ebb and flow. | AMF increased yield and reduced fruit total soluble solids and pH compared to the control. | [45] |
| Strawberry | G. intraradices, Bacillus velezensis | Coconut fiber/Ebb and flow. | AMF increased chlorophyll content index (SPAD) and decreased fruit total soluble solids, titratable acidity and pH compared to the control | [46] |
| Cucumber (Cucumis sativus) | G. mosseae | Sand/Ebb and flow. | AMF increased leaf SPAD, fresh weight, and fruit quality component (antioxidant activity and phenol content). | [110] |
| Snapdragons (Antirrhinum majus) | G. intraradices | 90% perlite and 10% peat-moss/Ebb and flow. | AMF significantly increased flower vase-life and reduced ethylene production. | [40] |
| Cucumber | G. mossea | No substrate/Float culture. | AMF-plants had higher shoot SPAD, gas exchange (photosynthesis rate, transpiration, and stomatal conductance), root phenol content, and antioxidant activity. | [104] |
| Pepper (Capsicum annuum) | G.caledonium and G. clarum | Perlite/Ebb and flow. | AMF-increased shoot dry weight. | [39] |
| Tomato | Piriformospora indica | Sand/Ebb and flow. | AMF increased leaves biomass by 20% and reduced disease severity caused by Verticillium dahliae by more than 30%. | [111] |
| Tomato | G. monosporum, G. vesiculiferum, G. deserticola, G. intraradices, G. mosseae | Sawdust/Ebb and flow. | Inoculation with G. monosporum and G. mosseae significantly increased fruit yield and fruit number and reduced Fusarium oxysporum root infection compared to the untreated control plants. | [10] |
| gerbera (Gerbera jamesonii) | G. mosseae and G. intradices | Sand/Ebb and flow. | AMF inoculation increased yield by 16% and vase life by 19%. | [112] |
| Tomato | G. fasciculatum | Perlite/Ebb and flow. | AMF increased yield significantly. | [47] |
| Tomato | 9 species of mychorriza; G. intraradices, G. clarum, G. mosseae, G. margarita, G. aggregatum, G. etunicatum, G. monosporus, G. deserticola, G. brasilianum | Peatmoss: perlite (1:1, v/v)/Ebb and flow. | Mychorriza treatment increased tomato juice electrical conductivity and reduced vitamin C. | [113] |
| Melon (Cucumis melo) | G. intraradices, G. aggregatum, G. mosseage, G. clarum, G. monosporus, Glomus deserticola, G. brasilianum, G. etunicatum, Gigaspor margari | Coco-peat: perlite (1:1, v/v)/Ebb and flow. | Growing AMF-inoculated watermelon seedlings in hydroponic system (80% full strength nutrient) increased total yield by 49.5% (12.4 vs. 8.3 kg/m2) compared to the control. | [106] |
| Pepper | Mixture of 9 AMF species: G. intraradices, G. aggregatum, G. mosseae, G. clarum, G. monosporus, G. deserticola, G. brasilianum, G. etunicatum, and G. margarita | Coco pith slabs/Ebb and flow. | The use of 80% mineral fertilizers in combination with mycorrhiza and bacteria, provided a 32.4% higher yield than the control (100% mineral fertilizer without bio-fertilizers). | [18] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Othman, Y.A.; Alananbeh, K.M.; Tahat, M.M. Can Arbuscular Mycorrhizal Fungi Enhance Crop Productivity and Quality in Hydroponics? A Meta-Analysis. Sustainability 2024, 16, 3662. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093662
AMA Style
Othman YA, Alananbeh KM, Tahat MM. Can Arbuscular Mycorrhizal Fungi Enhance Crop Productivity and Quality in Hydroponics? A Meta-Analysis. Sustainability. 2024; 16(9):3662. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093662
Chicago/Turabian StyleOthman, Yahia A., Kholoud M. Alananbeh, and Monther M. Tahat. 2024. "Can Arbuscular Mycorrhizal Fungi Enhance Crop Productivity and Quality in Hydroponics? A Meta-Analysis" Sustainability 16, no. 9: 3662. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093662
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.