
Geomorphological and Bioclimatic Relationships in the Occurrence of Species of Agro-Extractivist Interest in the Cerrado Biome

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Target Species
2.3. Environmental Variables to Species Distribution Modeling
2.4. Data Collection for Species Distribution Modeling
2.5. Data Related to the Occurrence of Species
2.6. Selection of Variables to Adjust Species Distribution Models (SDM)
2.7. Algorithms Used, Processing Parameters, and Model Evaluation
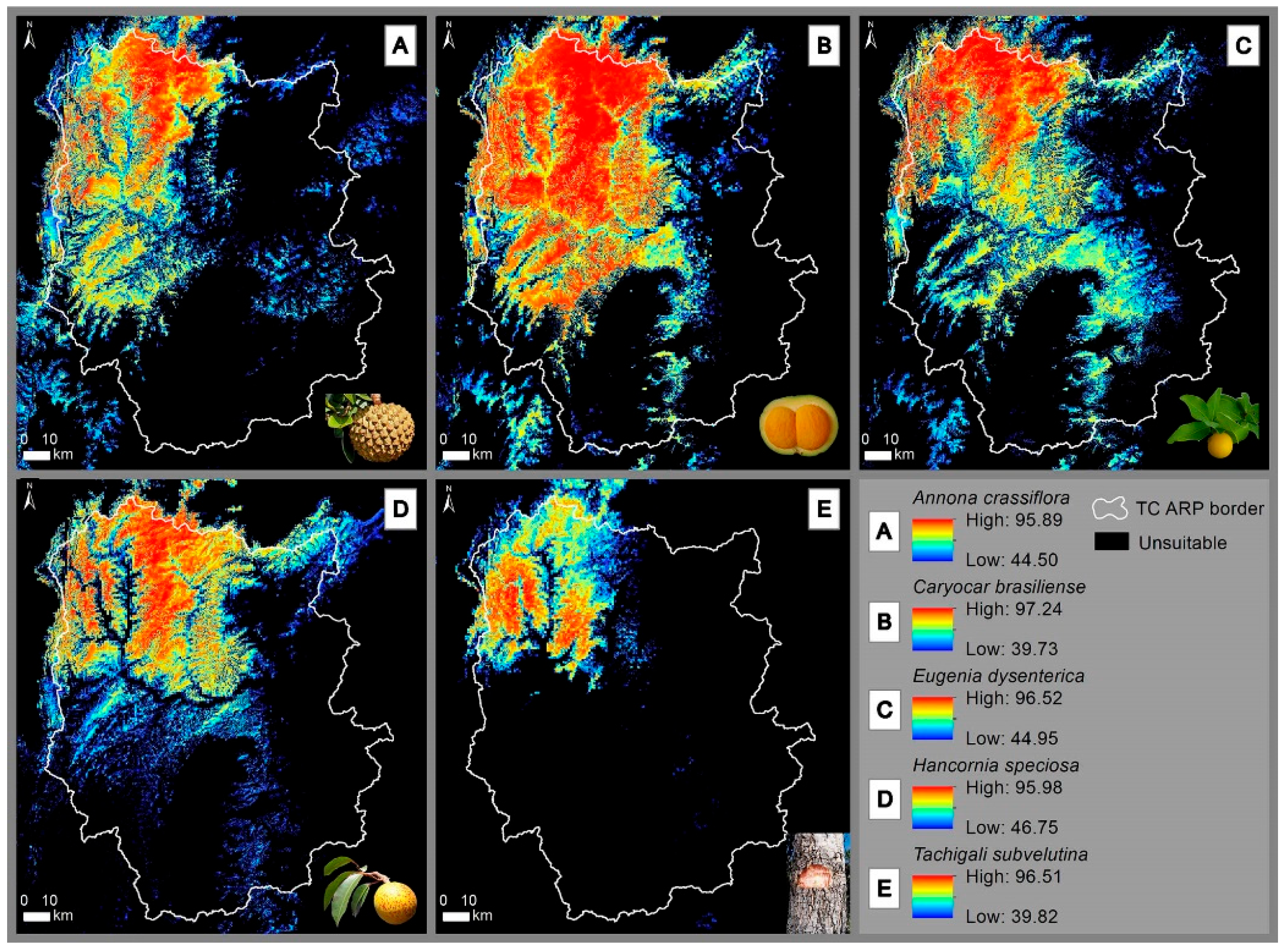
3. Results
3.1. Accuracy of Occurrence Suitability Models
3.2. Geomorphological and Bioclimatic Relationships in the Suitability of Occurrence of Agro-Extractivist Species Interest in the Cerrado Biome
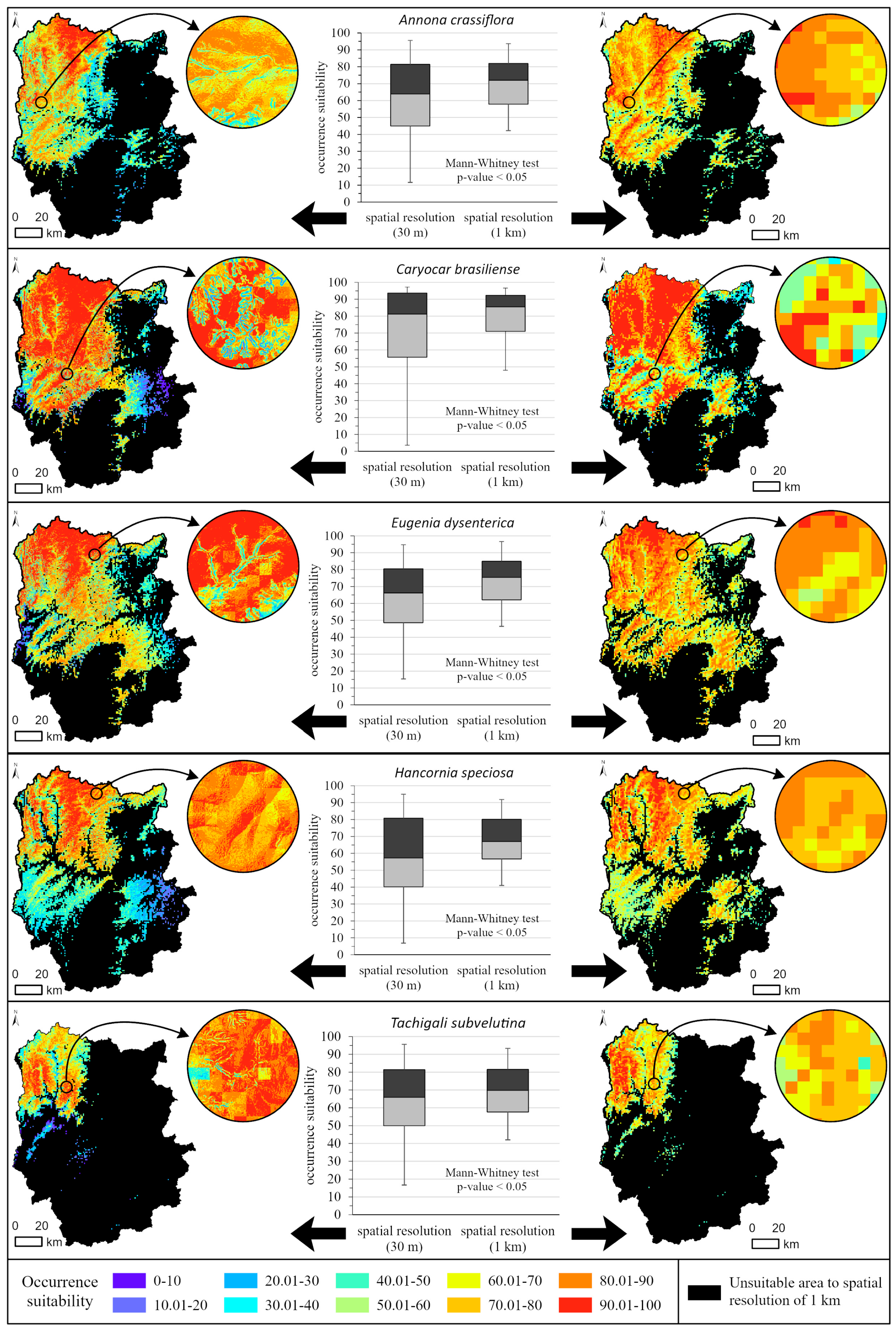
3.3. Improved Prediction of Suitability against Topographic Variables with High Spatial Resolution
4. Discussion
4.1. Bioclimatic Variables and the Suitability of Occurrence of Agro-Extractivist Species Interest in the Cerrado Biome
4.2. Geomorphological Variables and the Suitability of Occurrence of Agro-Extractivist Species Interest in the Cerrado Biome
4.3. Response of High Spatial Resolution Geomorphological Variables in Occurrence Suitability Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.L.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.M.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The Broad Footprint of Climate Change from Genes to Biomes to People. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.L.; Alves, T.S.; Oliveira, A.P.G.; Catalani, T.G.T.; Dalmas, F.B.; Paranhos Filho, A.C. Identificação e Quantificação Semiautomática de Desmatamento Por Sensoriamento Remoto. Res. Soc. Dev. 2020, 9, 30942721. [Google Scholar] [CrossRef]
- Evangelista-Vale, J.C.; Weihs, M.; José-Silva, L.; Arruda, R.; Sander, N.L.; Gomides, S.C.; Machado, T.M.; Pires-Oliveira, J.C.; Barros-Rosa, L.; Castuera-Oliveira, L.; et al. Climate Change May Affect the Future of Extractivism in the Brazilian Amazon. Biol. Conserv. 2021, 257, 109093. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological Annihilation via the Ongoing Sixth Mass Extinction Signaled by Vertebrate Population Losses and Declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.G.; Watson, J.E.M. Intact Ecosystems Provide Best Defence against Climate Change. Nat. Clim. Chang. 2016, 6, 122–124. [Google Scholar] [CrossRef]
- Rigonato, V.D.; Almeida, M.G.d. A Singularidade Do Cerrado: A Interrelação Das Populações Tradicionais Com as Fitofisionomias; Anais VIII EEREGEO; OGdG: Goiânia, Brazil, 2003. [Google Scholar]
- Mokarram, M.; Sathyamoorthy, D. Relationship between Landform Classification and Vegetation (Case Study: Southwest of Fars Province, Iran). Open Geosci. 2016, 8, 302–309. [Google Scholar] [CrossRef]
- Li, L.; Liu, M.; Ji, L.; Wang, F. Regional Analysis of the Potential Distribution of Heptacodium Miconioides and Its Competitor Species in China. Sustainability 2024, 16, 752. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting Species Distributions from Small Numbers of Occurrence Records: A Test Case Using Cryptic Geckos in Madagascar. J. Biogeogr. 2006, 34, 102–117. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the Impacts of Climate Change on the Distribution of Species: Are Bioclimate Envelope Models Useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Baek, S.; Kim, M.-J.; Lee, J.-H. Current and Future Distribution of Ricania Shantungensis (Hemiptera: Ricaniidae) in Korea: Application of Spatial Analysis to Select Relevant Environmental Variables for MaxEnt and CLIMEX Modeling. Forests 2019, 10, 490. [Google Scholar] [CrossRef]
- Mavárez, J.; Bézy, S.; Goeury, T.; Fernández, A.; Aubert, S. Current and Future Distributions of Espeletiinae (Asteraceae) in the Venezuelan Andes Based on Statistical Downscaling of Climatic Variables and Niche Modelling. Plant Ecol. Divers. 2019, 12, 633–647. [Google Scholar] [CrossRef]
- Nezer, O.; Bar-David, S.; Gueta, T.; Carmel, Y. High-Resolution Species-Distribution Model Based on Systematic Sampling and Indirect Observations. Biodivers. Conserv. 2017, 26, 421–437. [Google Scholar] [CrossRef]
- Hess, G.R.; Bartel, R.A.; Leidner, A.K.; Rosenfeld, K.M.; Rubino, M.J.; Snider, S.B.; Ricketts, T.H. Effectiveness of Biodiversity Indicators Varies with Extent, Grain, and Region. Biol. Conserv. 2006, 132, 448–457. [Google Scholar] [CrossRef]
- INMET. Banco de Dados Meteorológicos para Ensino e Pesquisa (BNDE). Available online: http://www.inmet.gov.br/portal/index.php?r=bdmep/bdmep (accessed on 4 December 2019).
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.D.M.; Sparovek, G. Köppen’s Climate Classification Map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Lima, I.L.P.; Scariot, A.; de Medeiros, M.B.; Sevilha, A.C. Diversidade e Uso de Plantas Do Cerrado Em Comunidade de Geraizeiros No Norte Do Estado de Minas Gerais, Brasil. Acta Bot. Bras. 2012, 26, 675–684. [Google Scholar] [CrossRef]
- Sevilha, A.C.; Scariot, A.; Matias, R.A.M.; Ávila, J.C.C.; Nascimento, M.M.; Viudes, P. Projeto Bem Diverso Sustenta e Inova: Integrando Conservação e Uso Sustentável da Biodiversidade às Práticas Produtivas de Produtos Florestais não Madeireiros e Sistemas Agroflorestais em Paisagens Florestais de Múltiplo Uso e Alto Valor de Conservação. 2021. Available online: https://bemdiverso.org.br/especies/ (accessed on 25 February 2024).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Global Land Areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Vasconcelos, V.; de Carvalho Junior, O.A.; de Souza Martins, E.; Junior, A.F.C.; Guimarães, R.F.; Gomes, R.A.T. Sistema de classificação geomorfométrica baseado em uma arquitetura sequencial em duas etapas: Árvore de decisão e classificador espectral, no parque nacional serra da canastra. Rev. Bras. De Geomorfol. 2012, 13, 171–186. [Google Scholar] [CrossRef]
- Sena-souza, J.P.; Martins, É.D.S.; Felipe, A.; Júnior, C.; Reatto, A.; Vasconcelos, V.; Gomes, M.P.; Abílio, O.; Júnior, D.C.; Reis, A.M. Mapeamento Geomorfológico Da Bacia Hidrográfica Do Rio São Bartolomeu, Escala 1:100.000; Embrapa: Brasília, Brazil, 2013; pp. 1–38. [Google Scholar]
- IBGE—Instituto Brasileiro de Geografia e Estatística. Manual Técnico de Geomorfologia, 2nd ed.; IBGE: Rio de Janeiro, Brazil, 2009; ISBN 978-85-240-4110-5. [Google Scholar]
- Sevilha, A.C. Systematic Conservation Planning for the Paranã River Basin, Brazil, under Climate Change. Ph.D. Thesis, James Cook University, Douglas, Australia, 2016. [Google Scholar]
- Arieira, J.; Padovani, C.R.; Schuchmann, K.-L.; Landeiro, V.L.; Santos, S.A. Modeling Climatic and Hydrological Suitability for an Encroaching Tree Species in a Neotropical Flooded Savanna. Ecol. Manag. 2018, 429, 244–255. [Google Scholar] [CrossRef]
- Coelho, G.L.N.; de Carvalho, L.M.T.; Gomide, L.R. Modelagem Preditiva de Distribuição de Espécies Pioneiras No Estado de Minas Gerais. Pesqui. Agropecu. Bras. 2016, 51, 207–214. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a Test of Whether One of Two Random Variables Is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Mazer, S. Potencial Produtivo de Plantas de Importância Socioeconômica da Reserva de Desenvolvimento Sustentável Nascentes Geraizeiras, Minas Gerais, Brasil. Mater’s Dissertation, Brasilia University, UnB, Brasília, Brazil, 2016. Available online: http://repositorio2.unb.br/jspui/handle/10482/22450 (accessed on 20 May 2021).
- Oliveira, L.W. Ecologia Populacional e Extrativismo de Frutos de Caryocar Brasiliense Camb. no Cerrado no Norte de Minas Gerais; IBICT: Brasilia, Brazil, 2009. [Google Scholar]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next Generation Python-Based GIS Toolkit for Landscape Genetic, Biogeographic and Species Distribution Model Analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Zwiener, V.P.; Lira-Noriega, A.; Grady, C.J.; Padial, A.A.; Vitule, J.R.S. Climate Change as a Driver of Biotic Homogenization of Woody Plants in the Atlantic Forest. Glob. Ecol. Biogeogr. 2018, 27, 298–309. [Google Scholar] [CrossRef]
- Castro, M.B.; Barbosa, A.C.M.C.; Pompeu, P.V.; Eisenlohr, P.V.; de Assis Pereira, G.; Apgaua, D.M.G.; Pires-Oliveira, J.C.; Barbosa, J.P.R.A.D.; Fontes, M.A.L.; dos Santos, R.M.; et al. Will the Emblematic Southern Conifer Araucaria Angustifolia Survive to Climate Change in Brazil? Biodivers. Conserv. 2020, 29, 591–607. [Google Scholar] [CrossRef]
- Leutner, B.; Horning, N. RStoolbox: Ferramentas Para Análise de Dados de Sensoriamento Remoto. Available online: https://cran.r-project.org/web/packages/RStoolbox/index.html (accessed on 15 January 2020).
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package “biomod2”. Available online: https://cran.r-project.org/web/packages/biomod2/biomod2.pdf (accessed on 6 January 2024).
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- De’ath, G.; Fabricius, K.E. Classification and Regression Trees: A Powerful Yet Simple Technique for Ecological Data Analysis. Ecology 2000, 81, 3178. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Buja, A. Flexible Discriminant Analysis by Optimal Scoring. J. Am. Stat. Assoc. 1994, 89, 1255. [Google Scholar] [CrossRef]
- Hastie, T.J.; Tibshirani, R.J. Generalized Additive Models; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Ridgeway, G. The State of Boosting. Comput. Sci. Stat. 1999, 31, 172–181. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Friedman, J.H. Multivariate Adaptive Regression Splines. Ann. Stat. 1991, 19, 123–141. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Package ‘biomod2′: Pacote R Versão 3.3-7.1. Pacote R Versão 3.3-7.1. Available online: http://cran.r-project.org/package=biomod2/ (accessed on 5 January 2020).
- José-Silva, L.; dos Santos, R.C.; de Lima, B.M.; Lima, M.; de Oliveira-Júnior, J.F.; Teodoro, P.E.; Eisenlohr, P.V.; da Silva Junior, C.A. Improving the Validation of Ecological Niche Models with Remote Sensing Analysis. Ecol Model. 2018, 380, 22–30. [Google Scholar] [CrossRef]
- Giannini, T.C.; Siqueira, M.F.; Acosta, A.L.; Barreto, F.C.C.; Saraiva, A.M.; Alves-dos-santos, I. Desafios Atuais Da Modelagem Preditiva de Distribuição de Espécies. Rodriguésia 2012, 63, 733–749. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-Absences for Species Distribution Models: How, Where and How Many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- ARAUJO, M.; NEW, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, S.; Sun, P.; Wang, T.; Wang, G.; Zhang, X.; Wang, L. Consensus Forecasting of Species Distributions: The Effects of Niche Model Performance and Niche Properties. PLoS ONE 2015, 10, e0120056. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Eisenlohr, P.V. Niche Modelling with Biomod2 Using 70 Environmental Variables Summarized in PCA Axes. Available online: https://github.com/pedroeisenlohr/niche_modelling (accessed on 15 October 2019).
- Bem Diverso. Geoportal Bem Diverso: Mapa Estrutural e Funcional Da Paisagem e de Uso Dos Solos Para a Região Do TC Alto Rio Pardo; Brasília. 2017. Available online: https://bemdiverso.maps.arcgis.com/apps/MapSeries/index.html?appid=4378836a7a4b42fb98280f707e70d13a (accessed on 9 January 2022).
- Williams, R.J.; Myers, B.A.; Muller, W.J.; Duff, G.A.; Eamus, D. Leaf Phenology of Woody Species in a North Australian Tropical Savanna. Ecology 1997, 78, 2542–2558. [Google Scholar] [CrossRef]
- Nabout, J.C.; Magalhães, M.R.; de Amorim Gomes, M.A.; da Cunha, H.F. The Impact of Global Climate Change on the Geographic Distribution and Sustainable Harvest of Hancornia Speciosa Gomes (Apocynaceae) in Brazil. Environ. Manag. 2016, 57, 814–821. [Google Scholar] [CrossRef]
- Carvalho, N.M.; Nakagawa, J. Sementes: Ciência, Tecnologia e Produção, 4th ed.; FUNEP: Jaboticabal, Brazil, 2000. [Google Scholar]
- Sousa, M.P.; Braga, L.F.; Braga, J.F.; Delachiave, M.E.A. Germinação de Sementes de Plantago Ovata Forsk. (Plantaginaceae): Temperatura e Fotoblastismo. Rev. Árvore 2008, 32, 51–57. [Google Scholar] [CrossRef]
- Pirani, F.R.; Sanchez, M.; Pedroni, F. Fenologia de Uma Comunidade Arbórea Em Cerrado Sentido Restrito, Barra Do Garças, MT, Brasil. Acta Bot. Bras. 2009, 23, 1096–1110. [Google Scholar] [CrossRef]
- Kubota, Y.; Murata, H.; Kikuzawa, K. Effects of Topographic Heterogeneity on Tree Species Richness and Stand Dynamics in a Subtropical Forest in Okinawa Island, Southern Japan. J. Ecol. 2004, 92, 230–240. [Google Scholar]
- Chapin, F.S.; Matson, P.A.; Mooney, H.A. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2002. [Google Scholar]
- Evans, I. An Integrated System of Terrain Analysis and Slope Mapping. Z. Geomorphol. Suppl. 1980, 36, 274–295. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Environmental Variables | VIF |
|---|---|---|
| Annona crassiflora | bio1—annual average temperature | 1.10 |
| bio3—isothermality | 1.25 | |
| bio12—annual precipitation | 1.21 | |
| Aspect—direction of slopes | 1.01 | |
| Slope—slope in degrees | 1.17 | |
| Caryocar brasiliense | bio1—annual average temperature | 1.10 |
| bio4—temperature seasonality | 1.57 | |
| bio12—annual precipitation | 1.45 | |
| Aspect—direction of slopes | 1.03 | |
| Slope—slope in degrees | 1.04 | |
| Eugenia dysenterica | bio4—temperature seasonality | 1.43 |
| bio9—average temperature of the driest quarter | 1.39 | |
| bio12—annual precipitation | 1.24 | |
| Slope—slope in degrees | 1.01 | |
| Hancornia speciosa | bio4—temperature seasonality | 1.43 |
| bio9—average temperature of the driest quarter | 1.10 | |
| bio12—annual precipitation | 1.26 | |
| Aspect—direction of slopes | 1.07 | |
| Tachigali subvelutina | bio3—isothermality | 2.18 |
| bio9—average temperature of the driest quarter | 2.06 | |
| bio16—precipitation in the wettest quarter | 1.11 | |
| Aspect—direction of slopes | 1.12 | |
| Slope—slope in degrees | 1.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, R.A.M.; Rezende, A.V.; Sevilha, A.C.; Scariot, A.O.; Matricardi, E.A.T.; Terribile, L.C. Geomorphological and Bioclimatic Relationships in the Occurrence of Species of Agro-Extractivist Interest in the Cerrado Biome. Sustainability 2024, 16, 3653. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093653
Matias RAM, Rezende AV, Sevilha AC, Scariot AO, Matricardi EAT, Terribile LC. Geomorphological and Bioclimatic Relationships in the Occurrence of Species of Agro-Extractivist Interest in the Cerrado Biome. Sustainability. 2024; 16(9):3653. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093653
Chicago/Turabian StyleMatias, Renan Augusto Miranda, Alba Valéria Rezende, Anderson Cássio Sevilha, Aldicir Osni Scariot, Eraldo Aparecido Trondoli Matricardi, and Levi Carina Terribile. 2024. "Geomorphological and Bioclimatic Relationships in the Occurrence of Species of Agro-Extractivist Interest in the Cerrado Biome" Sustainability 16, no. 9: 3653. https://0-doi-org.brum.beds.ac.uk/10.3390/su16093653